
Improving the Quality of Frozen Lamb by Microencapsulated Apple Polyphenols: Effects on Cathepsin Activity, Texture, and Protein Oxidation Stability

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Frozen Samples
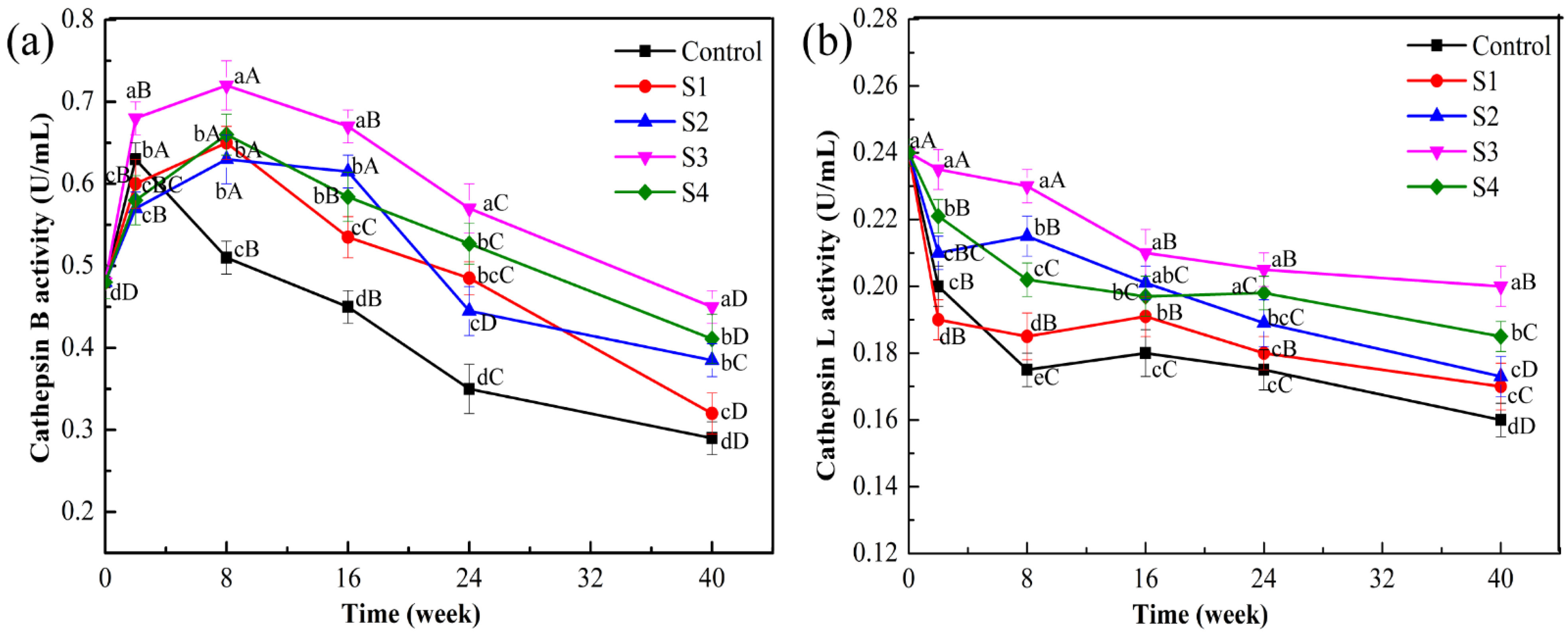
2.3. Determination of Cathepsin Activity in Lamb during Frozen Storage
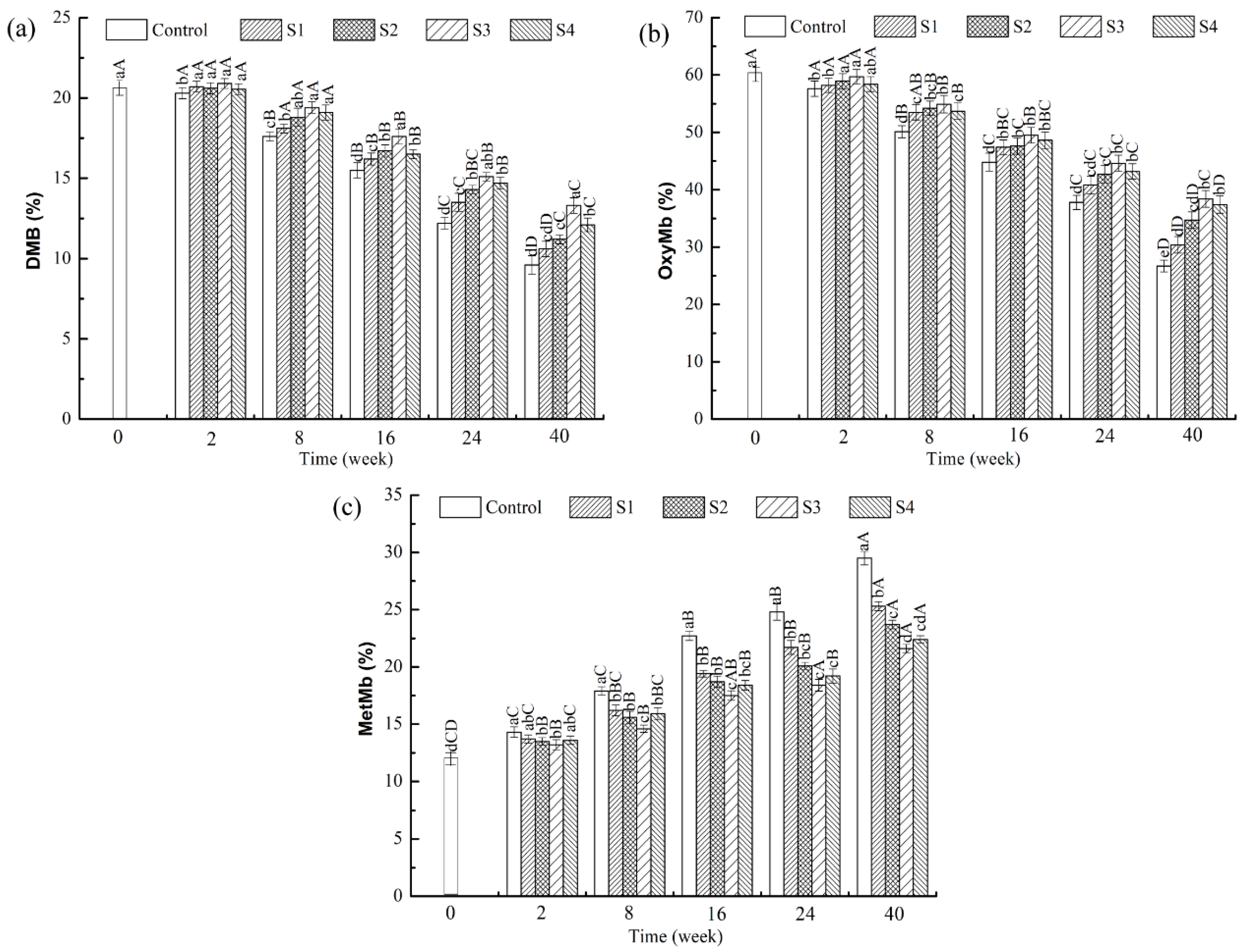
2.4. Determination of Deoxymyoglobin, Oxymyoglobin, and Metmyoglobin Contents
2.5. Texture Analysis
2.6. Extraction of Myofibrillar Proteins
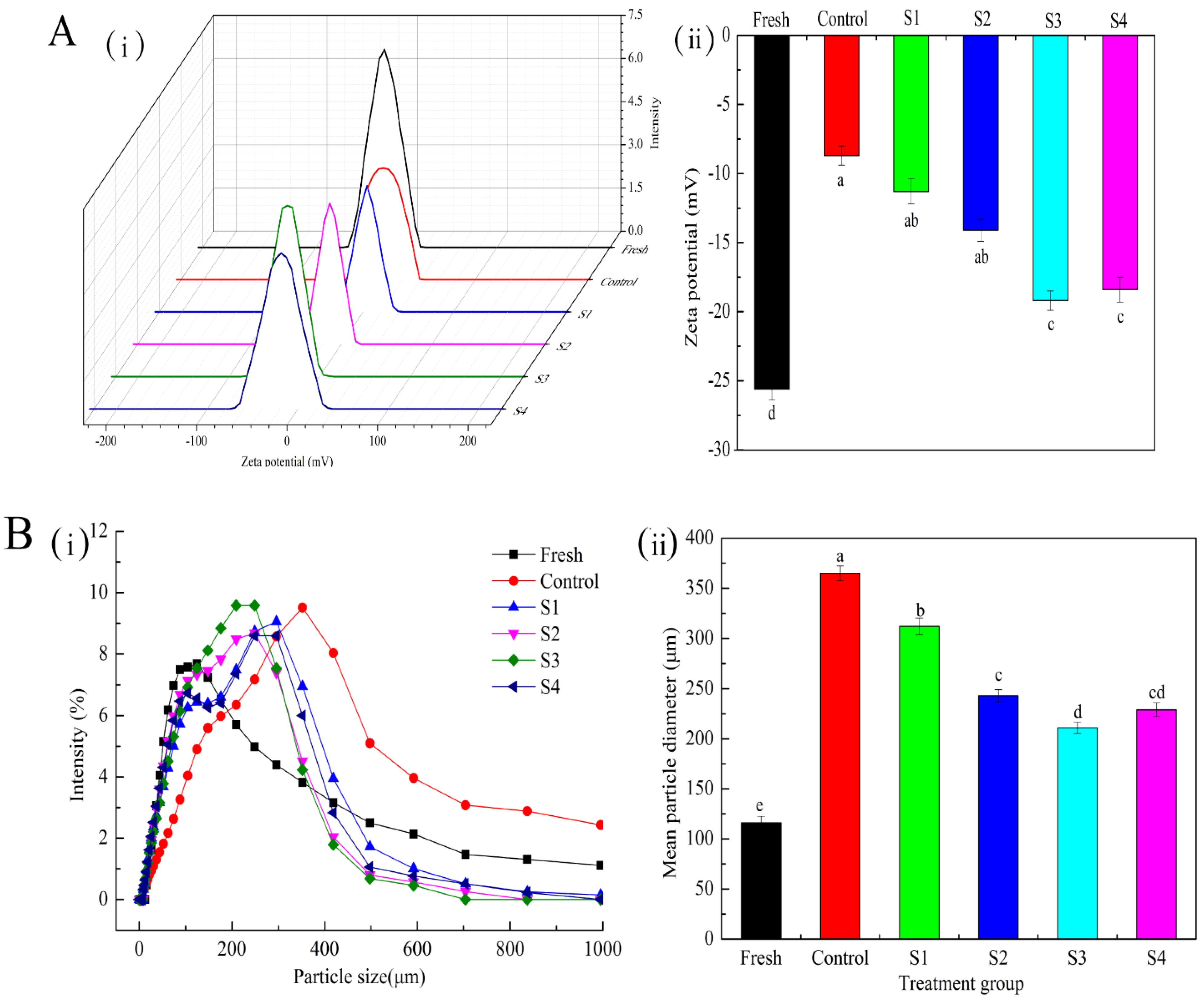
2.7. Zeta Potential Measurement
2.8. Particle Size Measurement
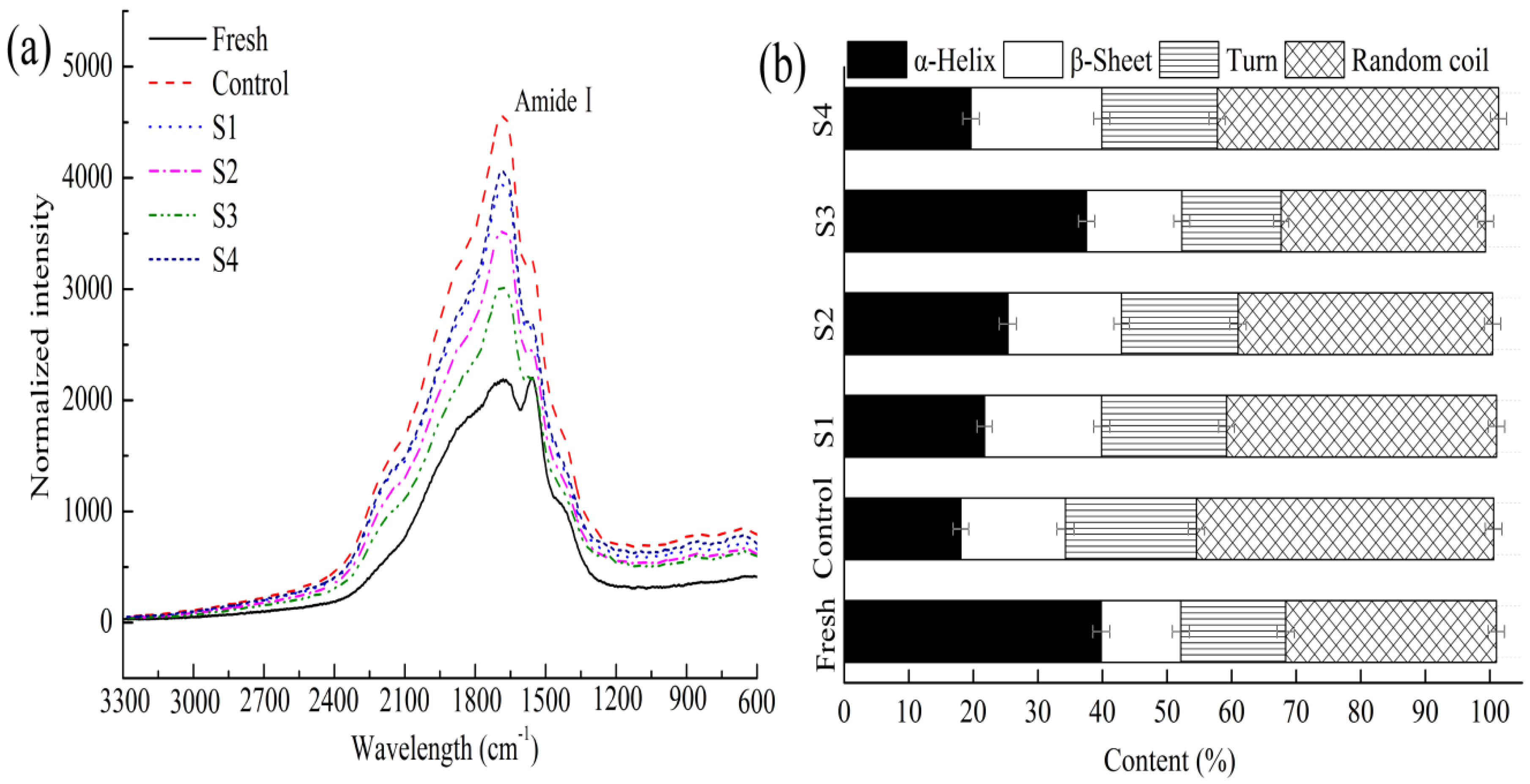
2.9. Raman Spectroscopy
2.10. In Vitro Degradation of Myofibrillar Protein by Cathepsin
2.11. Hematoxylin-Eosin Staining
2.12. Statistical Analysis
3. Results and Discussion
3.1. Effect of Different Treatments on Cathepsin Activities in the Frozen Lamb
3.2. Effects of Different Treatments on the Contents of Deoxymyoglobin, Oxymyoglobin, and Metmyoglobin in the Frozen Lamb
3.3. Effect of Different Treatments on the Texture of Lamb Meat during Frozen Storage
3.4. Effects of Different Treatments on Protein Stability during Frozen Storage
3.4.1. Zeta Potential
3.4.2. Particle Size
3.4.3. Secondary Structure in Myofibrillar Proteins
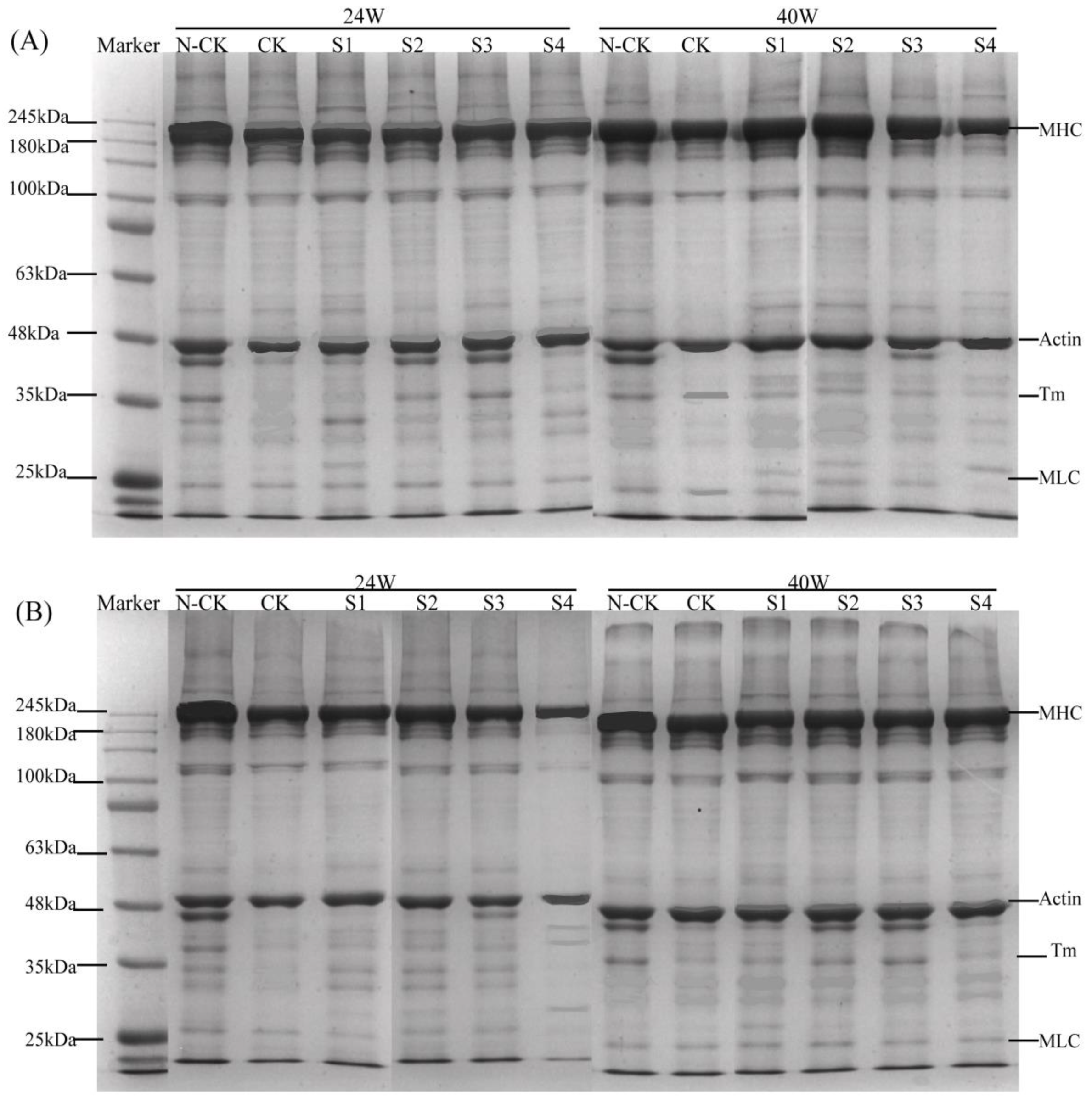
3.5. SDS–PAGE Analysis of Myofibrillar Protein Degradation by Cathepsin In Vitro
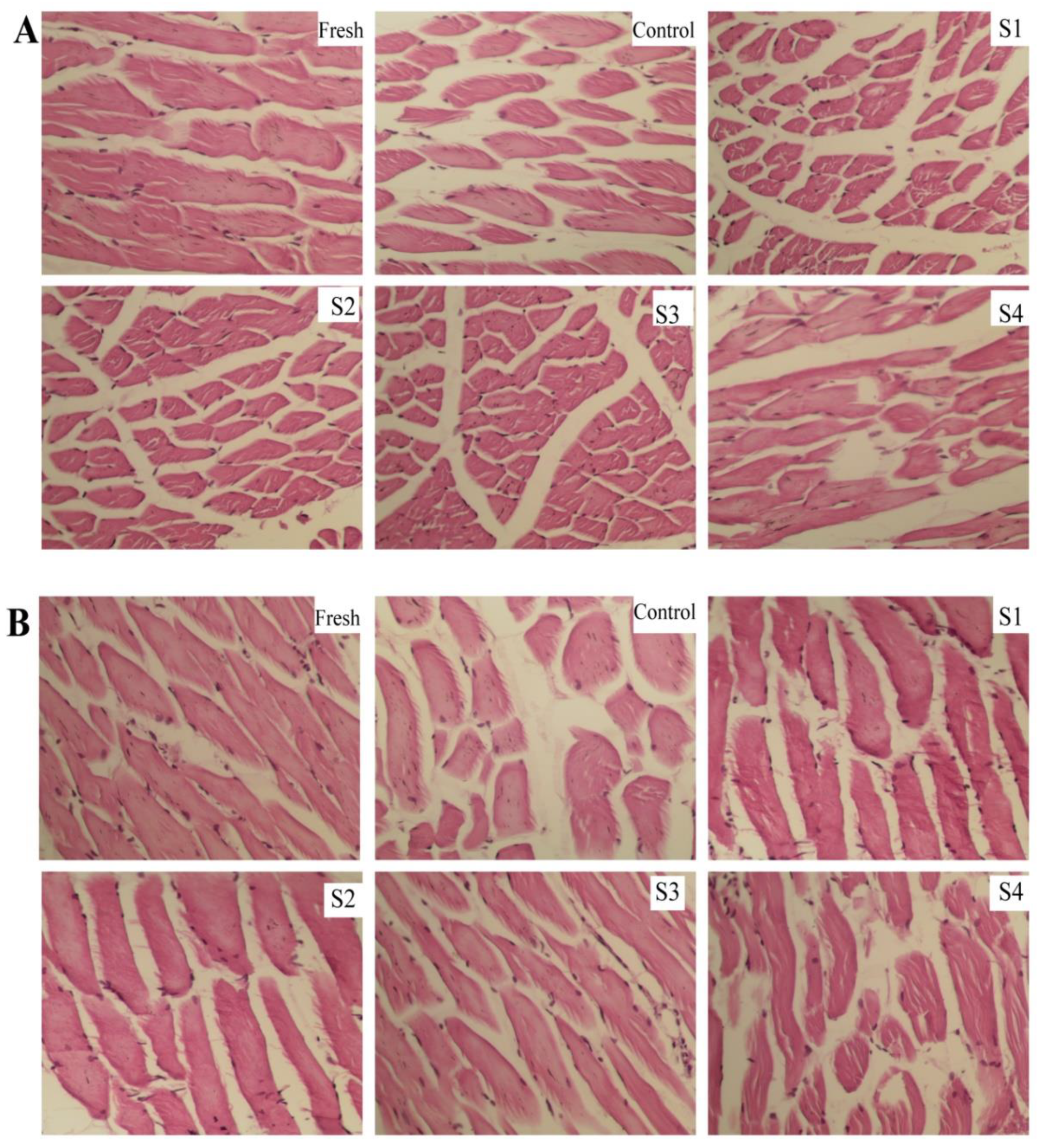
3.6. Effects of Different Treatments on the Microstructure during Frozen Storage
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Williams, P. Nutritional composition of red meat. J. Nutr. Diet. 2007, 64, S113–S119. [Google Scholar] [CrossRef] [Green Version]
- Deards, B.; Leith, R.; Mifsud, C.; Murray, C.; Martin, P.; Gleeson, T. Live Export Trade Assessment; ABARES Report to Client Prepared for the Live Animal Exports Reform Taskforce of the Department of Agriculture; ABARES: Canberra, Australia, 2014. [Google Scholar]
- Fernandes, R.D.P.P.; Freire, M.T.D.A.; Carrer, C.D.C.; Trindade, M.A. Evaluation of physicochemical, microbiological and sensory stability of frozen stored vacuum-packed lamb meat. J. Integr. Agric. 2013, 12, 1946–1952. [Google Scholar] [CrossRef]
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estévez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Guyon, C.; Meynier, A.; de Lamballerie, M. Protein and lipid oxidation in meat: A review with emphasis on high-pressure treatments. Trends Food Sci. Technol. 2016, 50, 131–143. [Google Scholar] [CrossRef]
- Qiu, H.H.; Guo, X.; Deng, X.R.; Guo, X.B.; Mao, X.Y.; Xu, C.J.; Zhang, J. The influence of endogenous cathepsin in different subcellular fractions on the quality deterioration of Northern pike (Esox lucius) fillets during refrigeration and partial freezing storage. Food Sci. Biotechnol. 2020, 29, 1331–1341. [Google Scholar] [CrossRef]
- Jenkelunas, P.J.; Ecy, L.C. Production and assessment of pacific hake (merluccius productus) hydrolysates as cryoprotectants for frozen fish mince. Food Chem. 2018, 239, 535–543. [Google Scholar] [CrossRef]
- Giddings, G.G.; Hill, L.H. Relationship of freezing preservation parameters to texture-related structural damage to thermally processed crustacean muscle. J. Food Process. Preserv. 2010, 2, 249–264. [Google Scholar] [CrossRef]
- Shah, M.A.; Bosco, S.J.; Mir, S.A. Plant extracts as natural antioxidants in meat and meat products. Meat Sci. 2014, 98, 21–33. [Google Scholar] [CrossRef]
- Jongberg, S.; Terkelsen Lde, S.; Miklos, R.; Lund, M.N. Green tea extract impairs meat emulsion properties by disturbing protein disulfide cross-linking. Meat Sci. 2015, 100, 2–9. [Google Scholar] [CrossRef]
- Jongberg, S.; Torngren, M.A.; Gunvig, A.; Skibsted, L.H.; Lund, M.N. Effect of green tea or rosemary extract on protein oxidation in Bologna type sausages prepared from oxidatively stressed pork. Meat Sci. 2013, 93, 538–546. [Google Scholar] [CrossRef]
- Butstraen, C.; Salaün, F. Preparation of microcapsules by complex coacervation of gum Arabic and chitosan. Carbohydr. Polym. 2014, 99, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Li, D.L.; Zhu, M.J.; Liu, X.M.; Wang, Y.T.; Cheng, J.R. Insight into the effect of microcapsule technology on the processing stability of mulberry polyphenols. LWT Food Sci. Technol. 2020, 126, 109144. [Google Scholar] [CrossRef]
- Xu, L.; Cheng, J.R.; Liu, X.M.; Zhu, M.J. Effect of microencapsulated process on stability of mulberry polyphenol and oxidation property of dried minced pork slices during heat processing and storage. LWT Food Sci. Technol. 2019, 100, 62–68. [Google Scholar] [CrossRef]
- Cunha, L.C.M.; Monteiro, M.L.G.; da Costa-Lima, B.R.C.; Guedes-Oliveira, J.M.; Rodrigues, B.L.; Fortunato, A.R.; Baltar, J.D.; Tonon, R.V.; Koutchma, T.; Conte, C.A. Effect of microencapsulated extract of pitaya (Hylocereus costaricensis) peel on oxidative quality parameters of refrigerated ground pork patties subjected to UV-C radiation. J. Food Process. Preserv. 2021, 45, e15272. [Google Scholar] [CrossRef]
- Zheng, L.; Ding, Z.; Zhang, M.; Sun, J. Microencapsulation of bayberry polyphenols by ethyl cellulose: Preparation and characterization. J. Food Eng. 2011, 104, 89–95. [Google Scholar] [CrossRef]
- Zhong, Y.Y.; Han, P.; Sun, S.L.; An, N.; Ren, X.D.; Lu, S.L.; Wang, Q.L.; Dong, J. Effects of apple polyphenols and hydroxypropyl-beta-cyclodextrin inclusion complexes on the oxidation of myofibrillar proteins and microstructures in lamb during frozen storage. Food Chem. 2022, 375, 131874. [Google Scholar] [CrossRef]
- Carlez, A.; Veciana-Nogues, T.; Cheftel, J.C. Changes in colour and myoglobin of minced beef meat due to high pressure processing. LWT Food Sci. Technol. 1995, 28, 528–538. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Jiang, Q. The impact of desmin on texture and water-holding capacity of ice-stored grass carp (Ctenopharyngodon idella) fillet. Int. J. Food Sci. Technol. 2017, 52, 464–471. [Google Scholar] [CrossRef]
- Lefevre, F.; Fauconneau, B.; Thompson, J.W.; Gill, T.A. Thermal denaturation and aggregation properties of Atlantic salmon myofibrils and myosin from white and red muscles. J. Agric. Food Chem. 2007, 55, 4761–4770. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 171, 751–766. [Google Scholar] [CrossRef]
- Lee, W.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, W668–W673. [Google Scholar]
- Ladrat, C.; Verrezbagnis, V.; Noel, J.; Fleurence, J. In vitro proteolysis of myofibrillar and sarcoplasmic proteins of white muscle of sea bass (Dicentrarchus labrax L.): Effects of cathepsins B, D and L. Food Chem. 2003, 81, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.; Xu, Y.; Xia, W.; Jiang, Q.; Jiang, X. Differential role of endogenous cathepsin and microorganism in texture softening of ice-stored grass carp (Ctenopharyngodon idella) fillets. J. Sci. Food Agric. 2016, 96, 3233–3239. [Google Scholar] [CrossRef]
- Nagaraj, N.S.; Santhanam, K. Effect of frozen temperature and storage time on calpains, cathepsins (B, B+L, Hand D) and their endogenous inhibitors in goat muscles. J. Food Biochem. 2010, 30, 155–173. [Google Scholar] [CrossRef]
- Hultmann, L.; Rustad, T. Iced storage of Atlantic salmon (Salmo salar)-Effectson endogenous enzymes and their impact on muscle proteins and texture. Food Chem. 2004, 87, 31–41. [Google Scholar] [CrossRef]
- Mancini, R.A.; Hunt, M.C. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef]
- Sadeghinejad, N.; Sarteshnizi, R.A.; Gavlighi, H.A.; Barzegar, M. Pistachio green hull extract as a natural antioxidant in beef patties: Effect on lipid and protein oxidation, color deterioration, and microbial stability during chilled storage. LWT Food Sci. Technol. 2019, 102, 393–402. [Google Scholar] [CrossRef]
- Ganhão, R.; Morcuende, D.; Estévez, M. Protein oxidation in emulsified cooked burger patties with added fruit extracts: Influence on colour and texture deterioration during chill storage. Meat Sci. 2010, 85, 402–409. [Google Scholar] [CrossRef]
- Wang, J.; Tang, J.; Rasco, B.; Sablani, S.S.; Ovissipour, M.; Qu, Z. Kinetics of quality changes of shrimp (Litopenaeus setiferus) during pasteurization. Food Bioprocess Technol. 2018, 11, 1027–1038. [Google Scholar] [CrossRef]
- Sun, Y.N.; Zhang, M.; Bhandari, B.; Yang, C.H. Ultrasound treatment of frozen crayfish with chitosan Nano-composite water-retaining agent: Influence on cryopreservation and storage qualities. Food Res. Int. 2019, 126, 108670. [Google Scholar] [CrossRef]
- Wei, P.Y.; Zhu, K.X.; Cao, J.; Dong, Y.; Li, M.Z.; Shen, X.R.; Duan, Z.H.; Li, C. The inhibition mechanism of the texture deterioration of tilapia fillets during partial freezing after treatment with polyphenols. Food Chem. 2021, 335, 127647. [Google Scholar] [CrossRef] [PubMed]
- Shilpashree, B.; Arora, S.; Chawla, P.; Tomar, S. Effect of succinylation on physicochemical and functional properties of milk protein concentrate. Food Res. Int. 2015, 72, 223–230. [Google Scholar] [CrossRef]
- Arredondo-Parada, I.; Torres-Arreola, W.; Suarez-Jimenez, G.M.; Ramirez-Suarez, J.C.; Juarez-Onofre, J.E.; Rodriguez-Felix, F.; Marquez-Rios, E. Effect of ultrasound on physicochemical and foaming properties of a protein concentrate from giant squid (Dosidicus gigas) mantle. LWT Food Sci. Technol. 2020, 121, 108954. [Google Scholar] [CrossRef]
- Lametsch, R.; Knudsen, J.C.; ErtbjergR, P.; Oksbjerg, N.; Therkildsen, M. Novel method for determination of myofibril fragmentation post-mortem. Meat Sci. 2007, 75, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.G.; Ma, W.H.; Huang, J.R.; Xiong, Y.L.L. Effects of sodium pyrophosphate coupled with catechin on the oxidative stability and gelling properties of myofibrillar protein. Food Hydrocoll. 2020, 104, 105722. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Tang, X.; Ni, W.; Zhou, L. Effects of pulsed ultrasound on rheological and structural properties of chicken myofibrillar protein. Ultrason. Sonochem. 2017, 38, 225–233. [Google Scholar] [CrossRef]
- Sow, L.C.; Toh, N.Z.Y.; Wong, C.W.; Yang, H. Combination of sodium alginate with tilapia fish gelatin for improved texture properties and nanostructure modification. Food Hydrocoll. 2019, 94, 459–467. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, Y.; Zhao, L.; Chen, L.; He, Y.; Yang, H. Vacuum impregnation of fish gelatin combined with grape seed extract inhibits protein oxidation and degradation of chilled tilapia fillets. Food Chem. 2019, 294, 316–325. [Google Scholar] [CrossRef]
- Martínez, M.; Velazquez, G.; Cando, D.; Núñez-Flores, R.; Borderías, A.J.; Moreno, H. Effects of high pressure processing on protein fractions of blue crab (Callinectes sapidus) meat. Innov. Food Sci. Emerg. Technol. 2017, 41, 323–329. [Google Scholar] [CrossRef]
- Kasicka, V. Amino acids, peptides and proteins. Electrophoresis 2011, 32, 2777–2778. [Google Scholar] [CrossRef]
- Jia, N.; Wang, L.; Shao, J.; Liu, D.; Kong, B. Changes in the structural and gel properties of pork myofibrillar protein induced by catechin modification. Meat Sci. 2017, 127, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Xiong, Y.L.; Alderton, A.L. Concentration effects of hydroxyl radical oxidizing systems on biochemical properties of porcine muscle myofibrillar protein. Food Chem. 2007, 101, 1239–1246. [Google Scholar] [CrossRef]
- Tironi, V.; Lamballerie, M.D.; Lebail, A. Quality changes during the frozen storage of sea bass (Dicentrarchus labrax) muscle after pressure shift freezing and pressure assisted thawing. Innov. Food Sci. Emerg. Technol. 2010, 11, 565–573. [Google Scholar] [CrossRef]
- He, Q.; Zhu, L.; Shen, Y.; Lin, X.D.; Xiao, K.J. Evaluation of the effects of frozen storage on the microstructure of tilapia (Perciformes: Cichlidae) through fractal dimension method. LWT Food Sci. Technol. 2015, 64, 1283–1288. [Google Scholar] [CrossRef]
- He, Q.; Yang, Z.; Gong, B.; Wang, J.; Xiao, K.; Yang, S.T. Quality evaluation focusing on tissue fractal dimension and chemical changes for frozen tilapia with treatment by tangerine peel extract. Sci. Rep. 2017, 7, 42202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Zhang, X.L.; Shen, C.L.; Deng, S.G. Understanding the influence of carrageenan oligosaccharides and xylooligosaccharides on ice-crystal growth in peeled shrimp (Litopenaeus vannamei) during frozen storage. Food Funct. 2018, 9, 4394–4403. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks | Fresh | Control | S1 | S2 | S3 | S4 | |
|---|---|---|---|---|---|---|---|
| Hardness (kg) Springiness (mm) | 2 8 16 24 40 2 8 16 24 40 | 0.84 ± 0.02 aA 1.03 ± 0.01 aA | 0.81 ± 0.01 aA 0.76 ± 0.01 bB 0.61 ± 0.03 bcC 0.53 ± 0.02 dC 0.31 ± 0.04 dD 0.98 ± 0.02 aA 0.83 ± 0.03 cB 0.79 ± 0.01 cBC 0.74 ± 0.02 cC 0.63 ± 0.03 dD | 0.78 ± 0.02 bB 0.77 ± 0.03 bDB 0.7 ± 0.01 bB 0.65 ± 0.02 bcB 0.42 ± 0.03 cD 0.94 ± 0.01 bA 0.88 ± 0.02 bB 0.76 ± 0.02 cC 0.72 ± 0.01 cC 0.66 ± 0.02 cD | 0.81 ± 0.04 abA 0.76 ± 0.02 bCA 0.69 ± 0.01 bB 0.56 ± 0.03 cC 0.41 ± 0.04 cD 0.95 ± 0.02 bA 0.87 ± 0.01 bcB 0.79 ± 0.01 cC 0.76 ± 0.03 bcC 0.67 ± 0.02 cD | 0.86 ± 0.03 aA 0.79 ± 0.01 bA 0.73 ± 0.02 bA 0.66 ± 0.01 bcB 0.53 ± 0.02 bC 0.97 ± 0.02 aA 0.94 ± 0.01 aA 0.86 ± 0.02 bB 0.78 ± 0.02 bC 0.71 ± 0.03 bcC | 0.79 ± 0.02 bA 0.78 ± 0.04 bA 0.69 ± 0.03 bAB 0.62 ± 0.02 bcB 0.47 ± 0.02 bcC 0.95 ± 0.03 bA 0.88 ± 0.03 bB 0.82 ± 0.01 bcB 0.76 ± 0.02 bcC 0.68 ± 0.01 cD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Liu, Y.; Xing, L.; Zhao, M.; Wu, W.; Wang, Q.; Ji, H.; Dong, J. Improving the Quality of Frozen Lamb by Microencapsulated Apple Polyphenols: Effects on Cathepsin Activity, Texture, and Protein Oxidation Stability. Foods 2022, 11, 537. https://doi.org/10.3390/foods11040537
Zhong Y, Liu Y, Xing L, Zhao M, Wu W, Wang Q, Ji H, Dong J. Improving the Quality of Frozen Lamb by Microencapsulated Apple Polyphenols: Effects on Cathepsin Activity, Texture, and Protein Oxidation Stability. Foods. 2022; 11(4):537. https://doi.org/10.3390/foods11040537
Chicago/Turabian StyleZhong, Yuanyuan, Yangming Liu, Lijie Xing, Mou Zhao, Wenxia Wu, Qingling Wang, Hua Ji, and Juan Dong. 2022. "Improving the Quality of Frozen Lamb by Microencapsulated Apple Polyphenols: Effects on Cathepsin Activity, Texture, and Protein Oxidation Stability" Foods 11, no. 4: 537. https://doi.org/10.3390/foods11040537
APA StyleZhong, Y., Liu, Y., Xing, L., Zhao, M., Wu, W., Wang, Q., Ji, H., & Dong, J. (2022). Improving the Quality of Frozen Lamb by Microencapsulated Apple Polyphenols: Effects on Cathepsin Activity, Texture, and Protein Oxidation Stability. Foods, 11(4), 537. https://doi.org/10.3390/foods11040537