
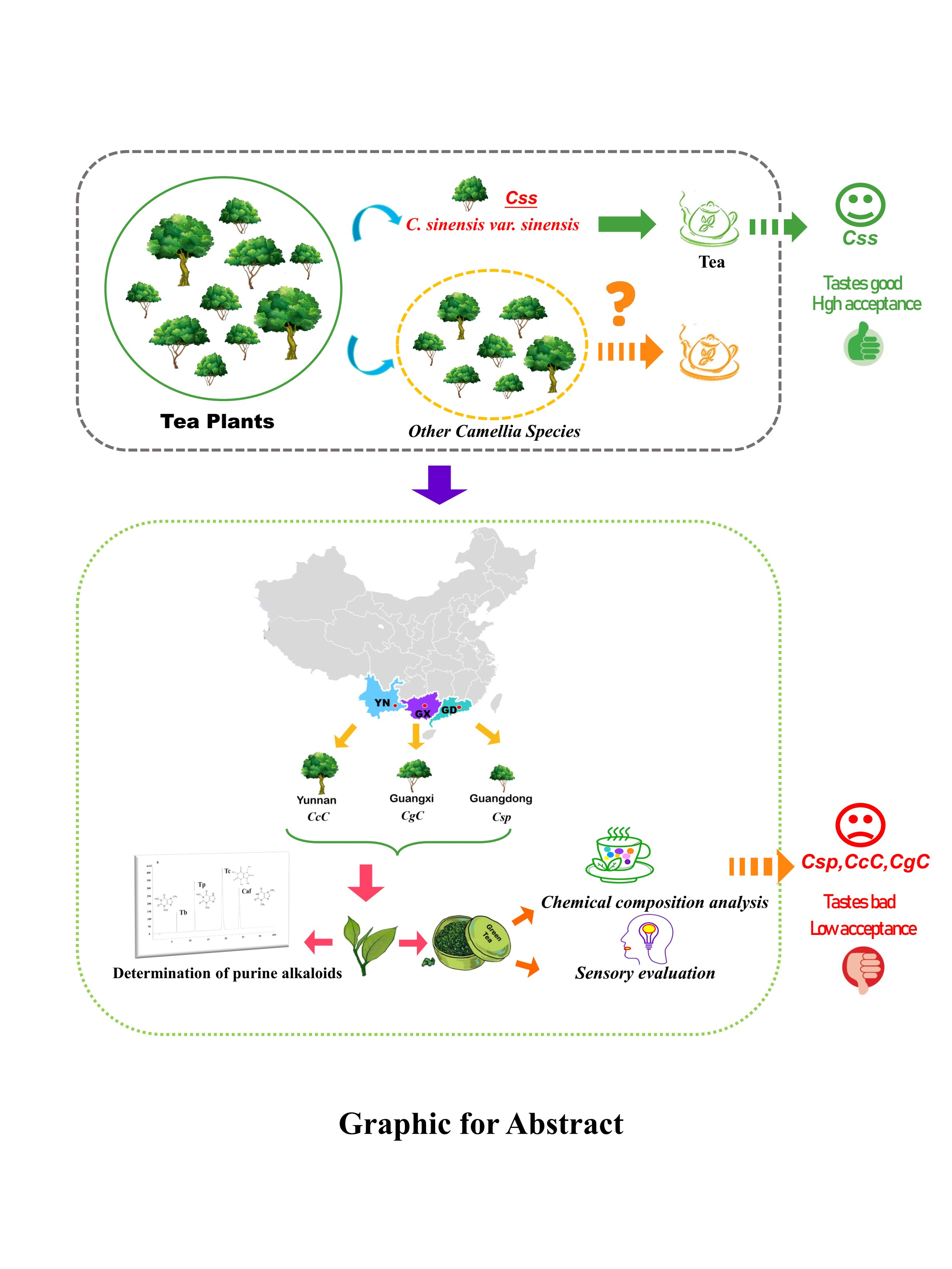
Comparative Analysis of Purine Alkaloids and Main Quality Components of the Three Camellia Species in China

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
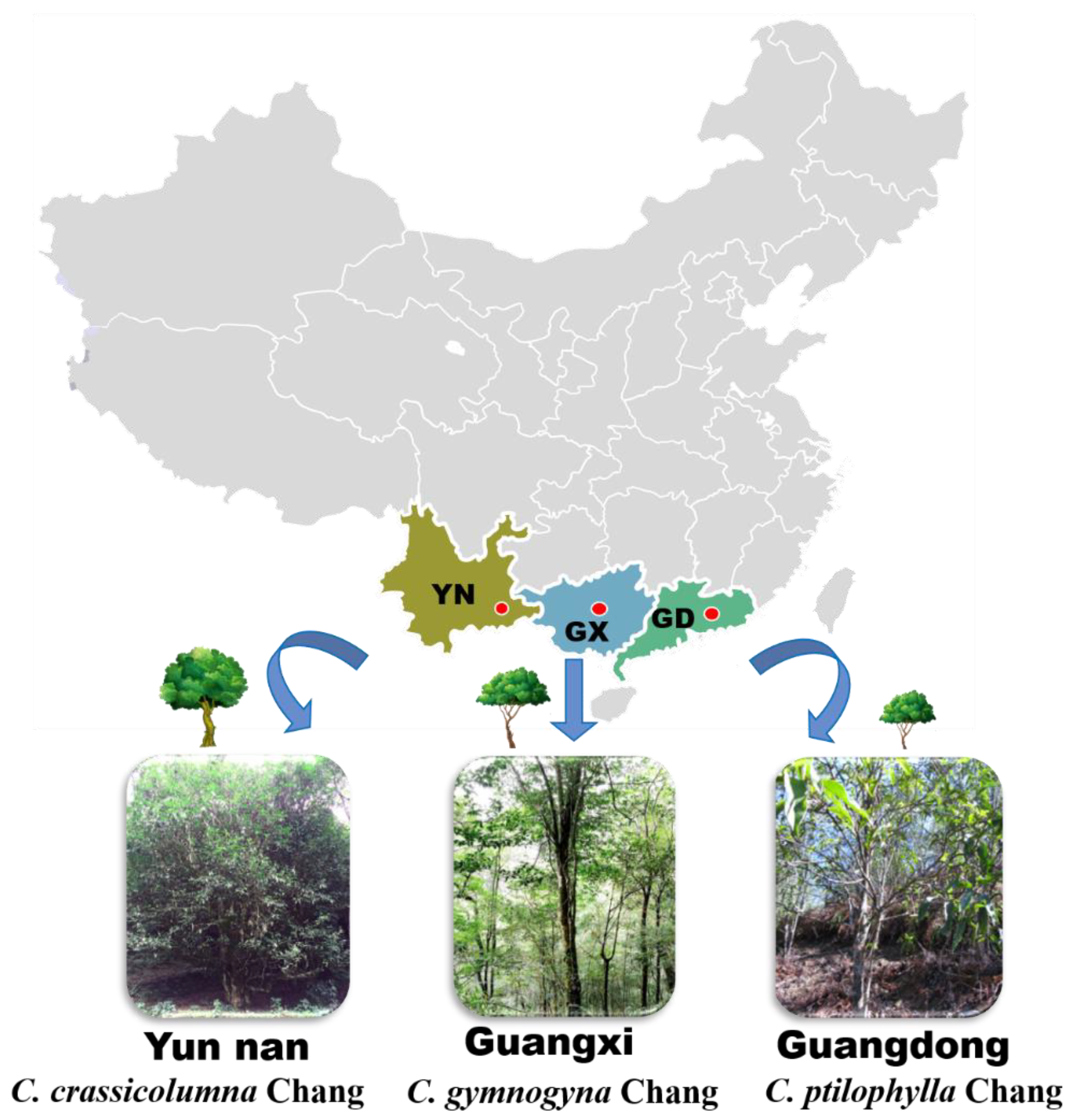
2.1. Collection and Pretreatment of Samples
2.2. Chemicals and Drugs
2.3. Collecting Data of Plant Morphology
2.4. Green Tea Processing Technique
2.5. Analysis of Purine Alkaloids in Tea Samples
2.6. Analysis of Catechins in Green Tea Samples
2.7. Analysis of Total Amino Acids in Green Tea Samples
2.8. Sensory Evaluation
2.9. Statistical Analysis of Data
3. Results and Discussion
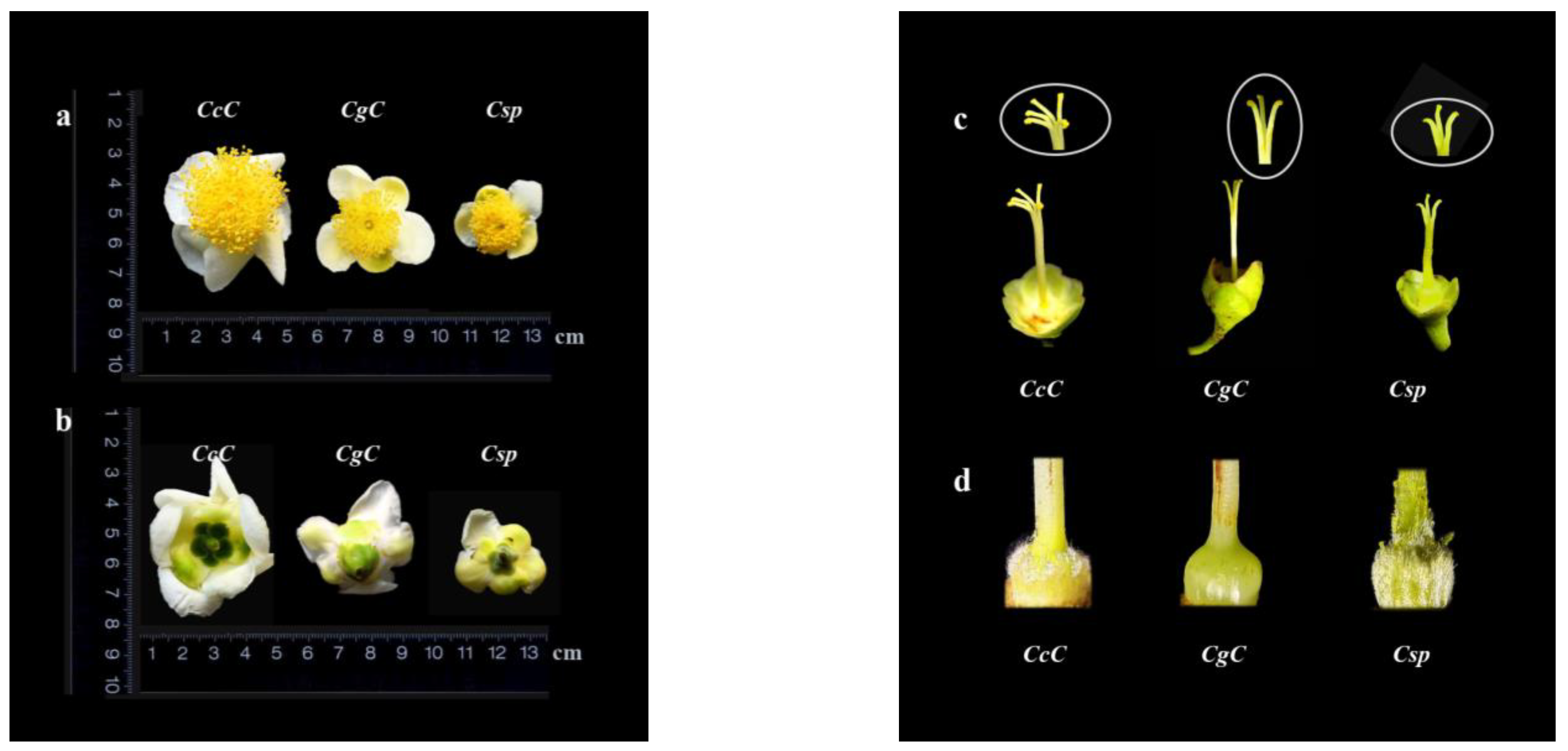
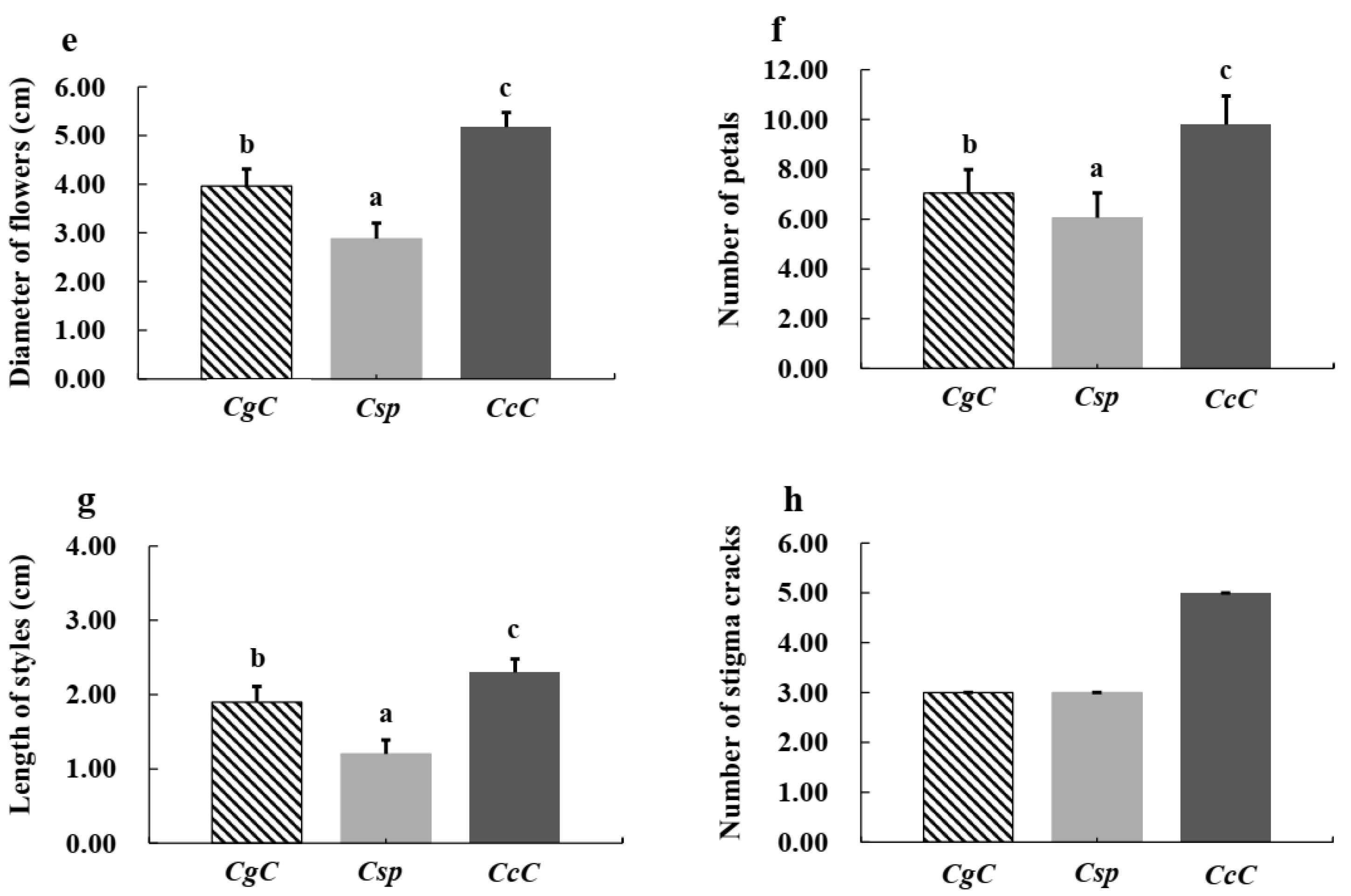
3.1. Morphological Characteristics of Three Camellia Species
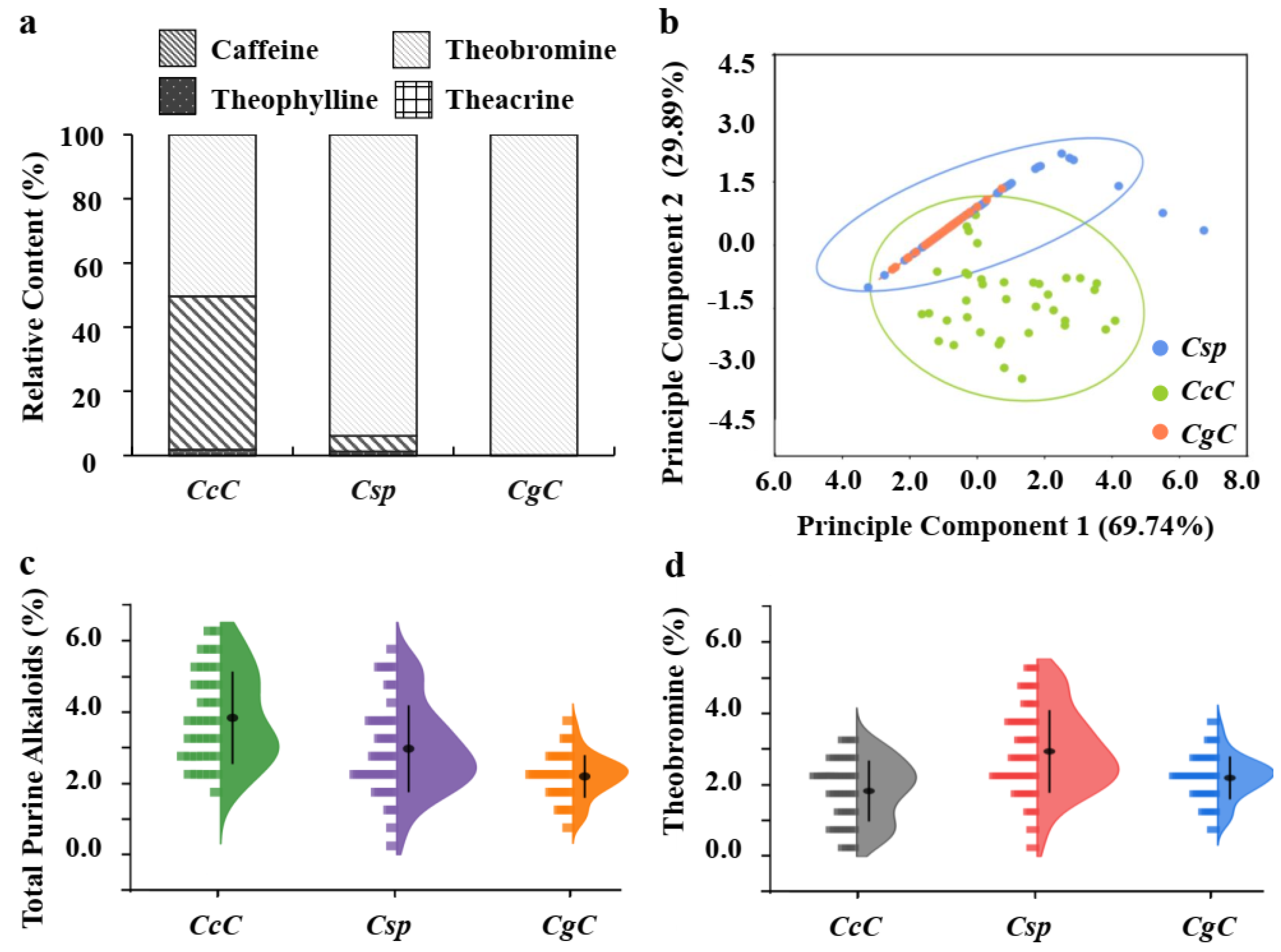
3.2. Patterns of Purine Alkaloids in Three Camellia Species
3.3. Variation of Theobromine Content in Three Camellia Species
3.4. Unique Germplasm Resources Discovered in Three Camellia Species
- Unique germplasm resources of C. sinensis var. pubilimba Chang (Csp)
- 2.
- Special germplasm resources C. crassicolumna Chang (CcC)
- 3.
- Special germplasm resources C. gymnogyna Chang (CgC)
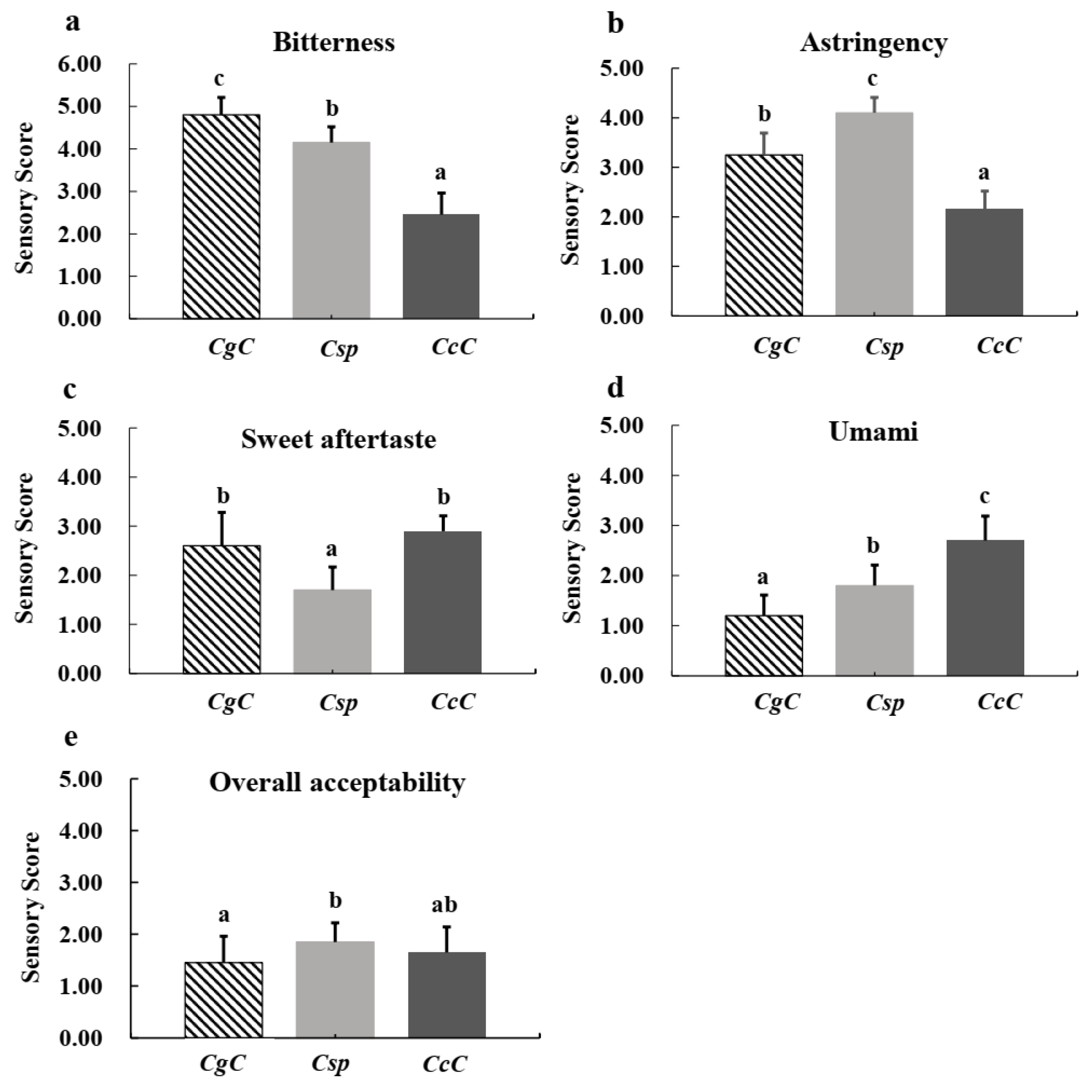
3.5. Quality Composition Analysis and Sensory Evaluation of Green Tea
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 529–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, E.H.; Zhang, H.B.; Sheng, J.; Li, K.; Zhang, Q.J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Gao, L. The tea tree genome provides insights into tea flavor and independent evolution of caffeine biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, M.; Gu, M.; Fan, S.; Huang, C.; Meyerhof, W. Bitter substances from plants used in traditional Chinese medicine exert biased activation of human bitter taste receptors. Chem. Biol. Drug Des. 2018, 91, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Ren, S.X. A Classification on the section tuberculate of Camellia. Acta Sci. Nat. Univ. Sunyatseni 1991, 20, 86–91. [Google Scholar]
- Chen, L. The research status of systematic taxonomy of Sect. Thea. J. Tea 1996, 2, 16–19. [Google Scholar]
- Chen, L.; Yu, F.L.; Tong, Q.Q. Discussions on phylogenetic classification and evolution of Sect. Thea. J. Tea Sci. 2000, 2, 89–94. [Google Scholar] [CrossRef]
- Ashihara, H.; Sano, H.; Crozier, A. Caffeine and related purine alkaloids: Biosynthesis, catabolism, function and genetic engineering. Phytochemistry 2008, 69, 841–856. [Google Scholar] [CrossRef]
- Uefuji, H.; Ogita, S.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Molecular cloning and functional characterization of three distinct N-methyltransferases involved in the caffeine biosynthetic pathway in coffee plants. Plant Physiol. 2003, 132, 372–380. [Google Scholar] [CrossRef] [Green Version]
- Mangels, D.R.; Mohler, E.R. Catechins as potential mediators of cardiovascular health. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 757–763. [Google Scholar] [CrossRef]
- Ma, J.; Jin, J.; Yao, M.Z.; Ma, C.L.; Xu, Y.X.; Hao, W.J.; Chen, L. Quantitative trait loci mapping for theobromine and caffeine contents in tea plant (Camellia sinensis). J. Agric. Food Chem. 2018, 66, 13321–13327. [Google Scholar] [CrossRef]
- Nehlig, A. Are we dependent upon coffee and caffeine? A review on human and animal data. Neurosci. Biobehav. Rev. 1999, 23, 563–576. [Google Scholar] [CrossRef]
- Glade, M.J. Caffeine-not just a stimulant. Nutrition 2010, 26, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.Q.; Ye, C.X.; Kato, M.; Crozier, A.; Ashihara, H. Theacrine (1,3,7,9-tetramethyluric acid) synthesis in leaves of a Chinese tea, kucha (Camellia assamica var. kucha). Phytochemistry 2002, 60, 129–134. [Google Scholar] [CrossRef]
- Xu, J.K.; Kurihara, H.; Zhao, L.; Yao, X.S. Theacrine, a special purine alkaloid with sedative and hypnotic properties from Cammelia assamica var. kucha in mice. J. Asian Nat. Prod. Res. 2007, 9, 665–672. [Google Scholar] [CrossRef]
- Jin, J.; Ma, J.; Ma, C. Determination of catechin content in representative Chinese tea germplasms. J. Agric. Food Chem. 2014, 62, 9436–9441. [Google Scholar] [CrossRef] [PubMed]
- Isozaki, T.; Tamura, H. Epigallocatechin Gallate (EGCG) inhibits the sulfation of 1-naphthol in a human colon carcinoma cell line, Caco-2. Biol. Pharm. Bull. 2001, 24, 1076–1078. [Google Scholar] [CrossRef] [Green Version]
- Reis, J.P.; Loria, C.M.; Steffen, L.M.; Zhou, X.; Van Horn, L.; Siscovick, D.S.; Carr, J.J. Coffee, decaffeinated coffee, caffeine, and tea consumption in young adulthood and atherosclerosis later in life: The CARDIA study. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2059–2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Hu, T.; Han, Y.; Huang, W.; Yuan, H.B.; Zhang, Y.T.; Jiang, Y.W. Preventive effects of catechins on cardiovascular disease. Molecules 2016, 21, 1759. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, G.W.; Ranadive, A.S.; Eisenberg, L.S.; Farrell, F.J.; Simons, R.; Manley, C.H.; Coggon, P. Contribution of polyphenolic compounds to the taste of tea. Acs Symp. 1976, 26, 14–46. [Google Scholar] [CrossRef]
- Xu, Y.Q.; Zhang, Y.N.; Chen, J.X.; Wang, F.; Du, Q.Z.; Yin, J.F. Quantitative analyses of the bitterness and astringency of catechins from green tea. Food Chem. 2018, 258, 16–24. [Google Scholar] [CrossRef]
- Mao, Z.F.; Shi, H.G.; Li, Q. Processing technology and quality of famous and excellent green tea. China Tea Processing 2007, 2, 51–53. [Google Scholar]
- Teng, J.; Zeng, Z.; Huang, Y. Composition characteristics of purine alkaloids and biochemical components of Camellia gymnogyna Chang. Guihaia 2018, 38, 568–576. [Google Scholar]
- Cao, Q.Q.; Zou, C.; Zhang, Y.H.; Du, Q.Z.; Yin, J.F.; Shi, J.; Xu, Y.Q. Improving the taste of autumn green tea with tannase. Food Chem. 2019, 277, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Yeo, A.S.L.; Low, M.Y.; Zhou, W. Identifying key non-volatile compounds in ready-to-drink green tea and their impact on taste profile. Food Chem. 2014, 155, 9–16. [Google Scholar] [CrossRef] [PubMed]
- İbanoglu, S.; Ainsworth, P.; Özer, E.A. Physical and sensory evaluation of a nutritionally balanced gluten-free extruded snack. J. Food Eng. 2006, 75, 469–472. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, J.; Chen, J.X.; Wang, F.; Du, Q.Z.; Jiang, Y.W.; Xu, Y.Q. Improving the sweet aftertaste of green tea infusion with tannase. Food Chem. 2016, 192, 470–476. [Google Scholar] [CrossRef]
- Maier, S.W.; Ludeker, W.; Gunther, K.P. Slop-pigments of photosynthetic biomembranes. Remote Sens. Environ. 1999, 68, 273–280. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Lu, J.L.; Wang, D.M.; Shi, X.G.; Yang, D.P.; Zheng, X.Q.; Ye, C.X. Determination of purine alkaloids and catechins in different parts of Camellia assamica var. kucha by HPLC-DAD/ESI-MS/MS. J. Sci. Food Agric. 2009, 89, 2024–2029. [Google Scholar] [CrossRef]
- Ye, C.X.; Lin, Y.C.; Su, J.Y.; Song, X.H.; Chang, H.D. Purine alkaloids in Camellia assamica var. kucha Chang. Acta Sci. Nat. UnivSun 1999, 38, 82–86. [Google Scholar] [CrossRef]
- Ye, C.X.; Zheng, X.Q.; Yuan, C.C.; Gao, K.; Shi, X.G.; Zhang, H.D. A summary of research on new caffeine-free tea resources cocoa tea. Guangdong Agric. Sci. 2001, 2, 12–15. [Google Scholar]
- Chang, H.T.; Ye, C.X.; Zhang, R.M. Diseovery of new tea resouree coeoa tea tree containing theobromine from China. Acta Sci. Nat. Univ. Sunyatseni 1988, 3, 131–133. [Google Scholar]
- Chen, J.; Wang, P.; Xia, Y.; Xu, M.; Pei, S. Genetic diversity and differentiation of Camellia sinensis L. (cultivated tea) and its wild relatives in Yunnan province of China, revealed by morphology, biochemistry and allozyme studies. Genet. Resour. Crop Evol. 2005, 52, 41–52. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, Y.J.; Yang, C.R.; Xu, M. Phenolic antioxidants from green tea produced from Camellia Crassicolumna Var. multiplex. J. Agric. Food Chem. 2009, 57, 586–590. [Google Scholar] [CrossRef]
- Jin, J.Q.; Zhou, C.Y.; Ma, C.L.; Yao, M.Z.; Ma, J.Q.; Chen, L. Identification on purine alkaloids of representative tea germplasms in China. J. Plant Genet. Resour. 2014, 15, 279–285. [Google Scholar]
- Ahmad, M.Z.; Li, P.; She, G.; Xia, E.; Benedito, V.A.; Wan, X.C.; Zhao, J. Genome-wide analysis of serine carboxypeptidase-like acyltransferase gene family for evolution and characterization of enzymes involved in the biosynthesis of galloylated catechins in the tea plant (Camellia sinensis). Front. Plant Sci. 2020, 11, 848. [Google Scholar] [CrossRef]
- Drewnowski, A. The science and complexity of bitter taste. Nutr. Rev. 2001, 59, 163–169. [Google Scholar] [CrossRef]
- Hayashi, N.; Ujihara, T.; Kohata, K. Reduction of catechin astringency by the complexation of gallate-type catechins with pectin. J. Agric. Chem. Soc. Jpn. 2005, 69, 1306–1310. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.F.; Zhang, Y.N.; Du, Q.Z.; Chen, J.X.; Yuan, H.B.; Xu, Y.Q. Effect of Ca2+ concentration on the tastes from the main chemicals in green tea infusions. Food Res. Int. 2014, 62, 941–946. [Google Scholar] [CrossRef]
- Ney, K.H. Bitterness of peptides: Amino acid composition and chain length. Food Taste Chem. 1979, 115, 149–173. [Google Scholar] [CrossRef]
- Chen, L.; Zeno, A.; Zong, M.; Chen, Z.M. Global Tea Breeding: Achievements, Challenges and Perspectives; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Xia, E.H.; Tong, W.; Wu, Q.; Wei, S.; Zhao, J.; Zhang, Z.Z.; Wan, X.C. Tea plant genomics: Achievements, challenges and perspectives. Hortic. Res. 2020, 7, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.K.; Ma, J.Q.; Liu, Y.F.; Chen, J.D.; Ni, D.J.; Chen, L. Identification and distribution of a single nucleotide polymorphism responsible for the catechin content in tea plants. Hortic. Res. 2020, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Wan, X. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, 4151–4158. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Color and Size | Number of Petals | Styles | Ovary |
|---|---|---|---|---|
| C. crassicolumna Chang | White, 4.8–5.5 cm in diameter | 9–12 | 5-parted, about 2.3 cm in average length, deeply cleft | Spherical, covered with white fluff |
| C. gymnogyna Chang | White, 3.4–4.6 cm in diameter | 6–8 | 3-parted, about 1.9 cm in average length, deeply cleft | Spherical, smooth and glabrous |
| C. ptilophylla Chang | White, 2.3–3.4 cm in diameter | 5–8 | 3-parted, about 1.2 cm in average length, shallow cleft | Spherical, covered with white fluff |
| Species | Tree Type | Leaf Size (cm) | Leaf Shape |
|---|---|---|---|
| C. crassicolumna Chang | Small trees or arbor, 6–15 m high or higher | (6.8–) 9–(–11.5) 15 × 3(5.5)–6 | Oval, oblong, oblong-elliptiptic |
| C. ptilophylla Chang | Shrub or small tree, about 4–7 m high | (7.6–) 8–12 × 3(5.5)–6 | Oblong, oblong-elliptiptic |
| C. gymnogyna Chang | Shrub or small tree, about 5–12 m high | 86 × 3.6–5 | Oval, oblong-elliptiptic |
| Species | Caffeine | Theobromine | Theophylline | Theacrine | TPA | |
|---|---|---|---|---|---|---|
| CcC (n = 38) | mean ± SD (%) | 1.93 ± 0.16 b | 1.83 ± 0.14 a | 0.07 ± 0.03 b | 0.00 ± 0.00 a | 3.83 ± 1.29 a |
| min (%) | 0.00 | 0.32 | 0.00 | 0.00 | 1.68 | |
| max (%) | 3.63 | 3.18 | 0.64 | 0.08 | 6.37 | |
| median (%) | 2.01 | 1.97 | 0.01 | 0.00 | 3.63 | |
| CV (%) | 0.50 | 0.46 | 2.28 | 4.09 | 0.34 | |
| kurtosis | −0.59 | −1.10 | 4.89 | 25.65 | −0.98 | |
| skewness | −0.24 | −0.23 | 2.40 | 4.89 | 0.31 | |
| Csp (n = 52) | mean ± SD (%) | 0.15 ± 0.08 a | 2.93 ± 0.16 b | 0.04 ± 0.01 ab | 0.00 ± 0.00 a | 3.11 ± 1.58 c |
| min (%) | 0.00 | 0.38 | 0.00 | 0.00 | 0.38 | |
| max (%) | 3.09 | 5.25 | 0.29 | 0.00 | 8.63 | |
| median (%) | 0.01 | 2.80 | 0.03 | 0.00 | 2.80 | |
| CV (%) | 0.00 | 0.39 | 2.29 | 0.00 | 0.51 | |
| kurtosis | 0.00 | −0.42 | 2.69 | 0.00 | 2.86 | |
| skewness | 0.00 | 0.26 | 2.05 | 0.00 | 1.44 | |
| CgC (n = 36) | mean ± SD (%) | 0.00 ± 0.00 a | 2.19 ± 0.10 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 2.19 ± 0.60 b |
| min (%) | 0.00 | 0.97 | 0.00 | 0.00 | 0.97 | |
| max (%) | 0.00 | 3.64 | 0.00 | 0.00 | 3.64 | |
| median (%) | 0.00 | 2.28 | 0.00 | 0.00 | 2.28 | |
| CV (%) | 0.00 | 0.40 | 0.00 | 0.00 | 0.27 | |
| kurtosis | 0.00 | 0.40 | 0.00 | 0.00 | 0.39 | |
| skewness | 0.00 | −0.11 | 0.00 | 0.00 | −0.11 |
| Variable | Principle Component | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| Caffeine | 0.41 | −0.71 | −0.30 |
| Theobromine | 0.39 | 0.71 | −0.31 |
| Theophylline | 0.02 | 0.01 | 0.86 |
| Theacrine | 0.00 | 0.00 | 0.03 |
| Total purine alkaloids | 0.82 | 0.01 | 0.27 |
| Eigenalue | 69.74% | 29.89% | 0.00 |
| Cumulative | 69.74% | 99.63% | 1.00 |
| Species | Sample Number | Caffeine | Theobromine | Theophylline | Theacrine |
|---|---|---|---|---|---|
| Csp | 1 | 3.09 ± 0.01 | 1.27 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| 40 | 0.00 ± 0.00 | 2.55 ± 0.00 | 0.29 ± 0.00 | 0.00 ± 0.00 | |
| 34 | 0.00 ± 0.00 | 3.71 ± 0.01 | 0.27 ± 0.01 | 0.00 ± 0.00 | |
| 33 | 0.00 ± 0.00 | 3.79 ± 0.01 | 0.26 ± 0.00 | 0.00 ± 0.00 | |
| 37 | 0.00 ± 0.00 | 3.15 ± 0.00 | 0.23 ± 0.01 | 0.00 ± 0.00 | |
| 20 | 0.02 ± 0.00 | 2.40 ± 0.01 | 0.22 ± 0.00 | 0.00 ± 0.00 | |
| 13 | 0.03 ± 0.00 | 2.14 ± 0.00 | 0.19 ± 0.00 | 0.00 ± 0.00 | |
| 26 | 0.01 ± 0.00 | 1.93 ± 0.00 | 0.18 ± 0.00 | 0.00 ± 0.00 | |
| 45 | 0.04 ± 0.00 | 2.33 ± 0.00 | 0.14 ± 0.00 | 0.00 ± 0.00 | |
| 47 | 0.00 ± 0.00 | 2.06 ± 0.01 | 0.13 ± 0.00 | 0.00 ± 0.00 | |
| CcC | 4 | 2.13 ± 0.07 | 2.76 ± 0.54 | 0.33 ± 0.02 | 0.03 ± 0.01 |
| 11 | 3.42 ± 0.03 | 0.68 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| 5 | 3.63 ± 0.03 | 2.51 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| 16 | 3.02 ± 0.01 | 0.65 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| 25 | 2.00 ± 0.05 | 2.43 ± 0.02 | 0.13 ± 0.02 | 0.02 ± 0.01 | |
| 23 | 0.30 ± 0.02 | 2.53 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| 13 | 0.01 ± 0.01 | 2.99 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| 28 | 0.00 ± 0.00 | 2.38 ± 0.02 | 0.51 ± 0.02 | 0.08 ± 0.01 |
| Csp | CcC | CgC | |
|---|---|---|---|
| GC | 1.70 ± 0.01 c | 0.00 ± 0.00 a | 0.79 ± 0.01 b |
| C | 2.25 ± 0.01 c | 0.00 ± 0.00 a | 0.39 ± 0.01 b |
| EGC | 0.69 ± 0.01 a | 1.57 ± 0.01 b | 1.73 ± 0.01 c |
| EGCG | 1.81 ± 0.01 c | 1.23 ± 0.01 a | 1.46 ± 0.01 b |
| EC | 0.64 ± 0.01 b | 0.75 ± 0.01 c | 0.35 ± 0.01 a |
| GCG | 5.57 ± 0.01 c | 0.00 ± 0.00 a | 0.02 ± 0.01 b |
| ECG | 0.28 ± 0.01 a | 0.46 ± 0.01 c | 0.38 ± 0.01 b |
| CG | 0.32 ± 0.01 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| EGCG+ECG | 2.09 ± 0.02 c | 1.69 ± 0.02 a | 1.84 ± 0.03 b |
| EGC+EC | 1.33 ± 0.01 a | 2.32 ± 0.03 c | 2.08 ± 0.01 b |
| Total Catechins | 13.25 ± 0.04c | 4.01 ± 0.01 a | 5.12 ± 0.01 b |
| Total Ester-type Catechins | 7.98 ± 0.03 c | 1.69 ± 0.02 a | 1.86 ± 0.03 b |
| Total Non-ester type Catechins | 5.27 ± 0.04 c | 2.32 ± 0.03 a | 3.26 ± 0.05 b |
| Percentage of Ester-type catechins in total catechins | 60.23% | 42.14% | 36.33% |
| Total Amino Acids | 1.06 ± 0.03 b | 1.31 ± 0.03 c | 0.75 ± 0.01 a |
| Caffeine | 0.07 ± 0.01 b | 1.56 ± 0.01 c | 0.00 ± 0.00 a |
| Theobromine | 3.12 ± 0.01 c | 1.62 ± 0.01 a | 2.25 ± 0.01 b |
| Theophylline | 0.01 ± 0.00 b | 0.03 ± 0.00 c | 0.00 ± 0.00 a |
| Theacrine | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Total Purine Alkaloids | 3.21 ± 0.02 b | 3.21 ± 0.01 b | 2.25 ± 0.01a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, W.; Zeng, Z.; Teng, J.; Rothenberg, D.O.; Zhou, M.; Lai, R.; Lai, X.; Zhao, W.; Li, D.; Yan, C.; et al. Comparative Analysis of Purine Alkaloids and Main Quality Components of the Three Camellia Species in China. Foods 2022, 11, 627. https://doi.org/10.3390/foods11050627
Zeng W, Zeng Z, Teng J, Rothenberg DO, Zhou M, Lai R, Lai X, Zhao W, Li D, Yan C, et al. Comparative Analysis of Purine Alkaloids and Main Quality Components of the Three Camellia Species in China. Foods. 2022; 11(5):627. https://doi.org/10.3390/foods11050627
Chicago/Turabian StyleZeng, Wen, Zhen Zeng, Jie Teng, Dylan O’Neill Rothenberg, Mengzhen Zhou, Ronghui Lai, Xingfei Lai, Wenfang Zhao, Dan Li, Changyu Yan, and et al. 2022. "Comparative Analysis of Purine Alkaloids and Main Quality Components of the Three Camellia Species in China" Foods 11, no. 5: 627. https://doi.org/10.3390/foods11050627
APA StyleZeng, W., Zeng, Z., Teng, J., Rothenberg, D. O., Zhou, M., Lai, R., Lai, X., Zhao, W., Li, D., Yan, C., & Huang, Y. (2022). Comparative Analysis of Purine Alkaloids and Main Quality Components of the Three Camellia Species in China. Foods, 11(5), 627. https://doi.org/10.3390/foods11050627