
Chlorella pyrenoidosa Polysaccharides as a Prebiotic to Modulate Gut Microbiota: Physicochemical Properties and Fermentation Characteristics In Vitro


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Polysaccharide Extraction
2.3. Physicochemical Analysis
2.4. Simulated Saliva Digestion
2.5. Simulated Gastrointestinal Digestion
2.6. In Vitro Fermentation
2.7. Determination of pH
2.8. Analysis of Short-Chain Fatty Acids (SCFAs)
2.9. Analysis of Gut Microbiota
2.10. Statistical Analysis
3. Results and Discussion
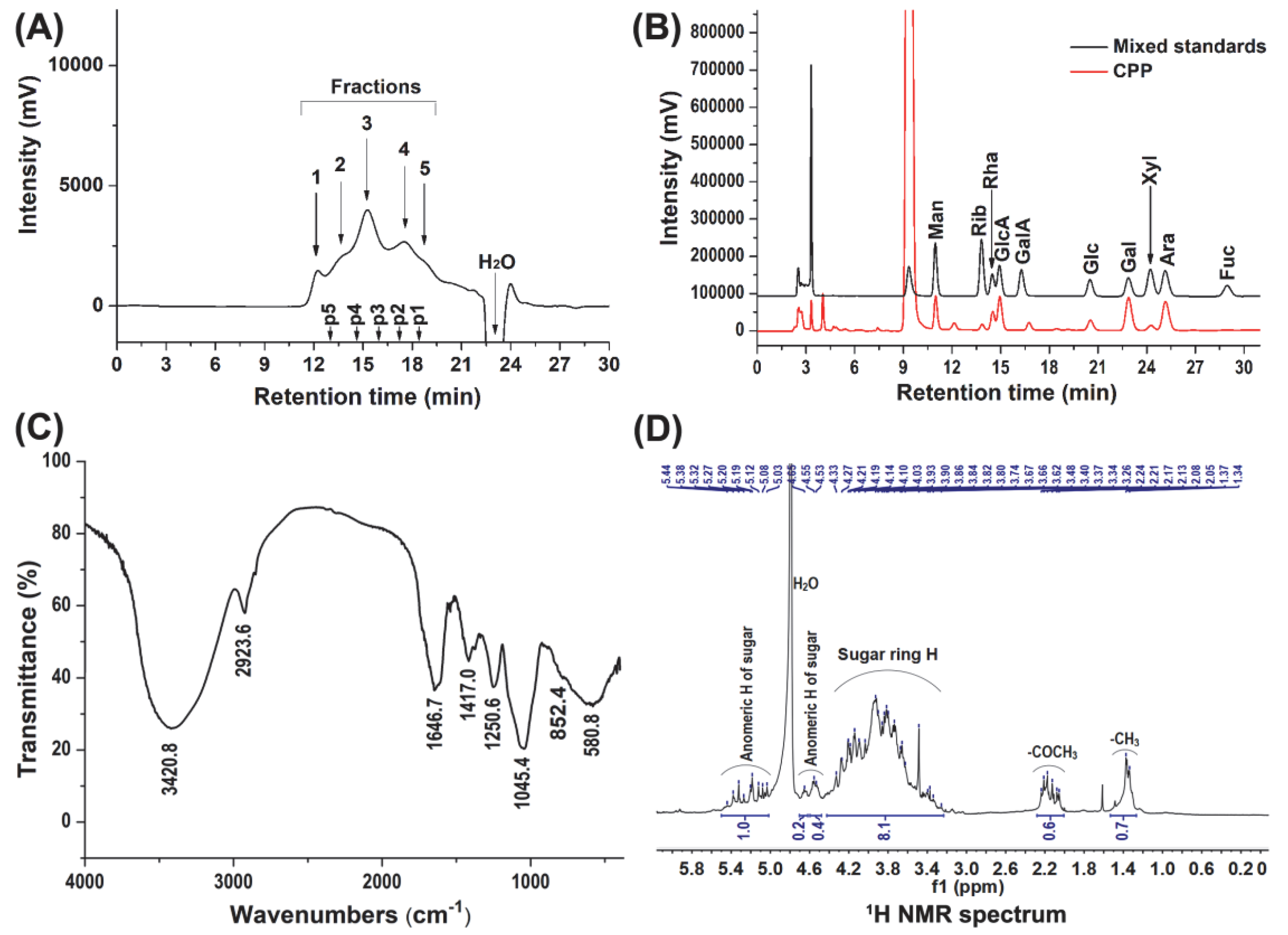
3.1. Physicochemical Characteristics of CPP
3.2. Characterization of CPP during Simulated Saliva and Gastrointestinal Digestion
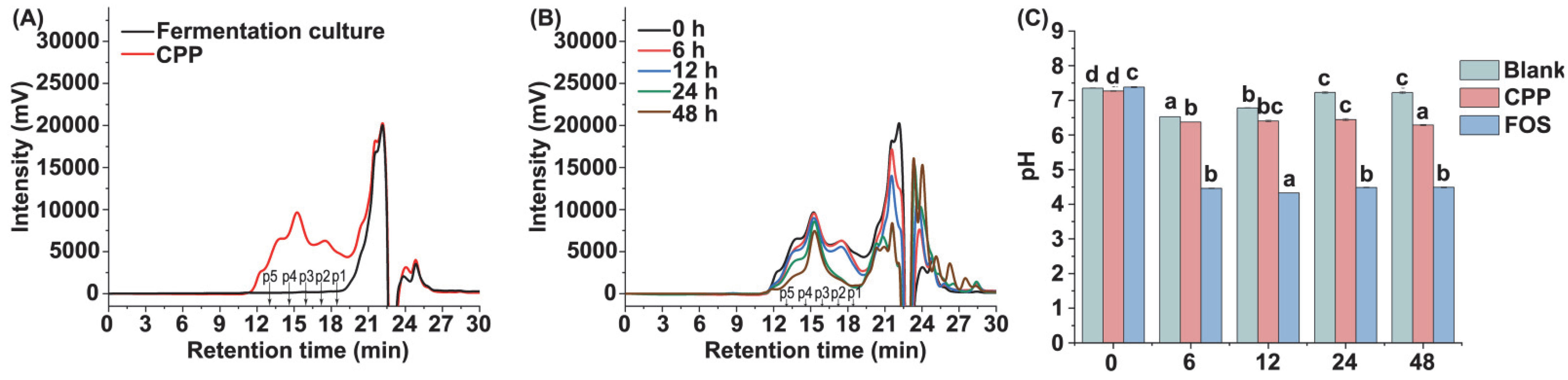
3.3. Characterization of CPP during In Vitro Fermentation
3.4. Changes of pH Value during Fermentation
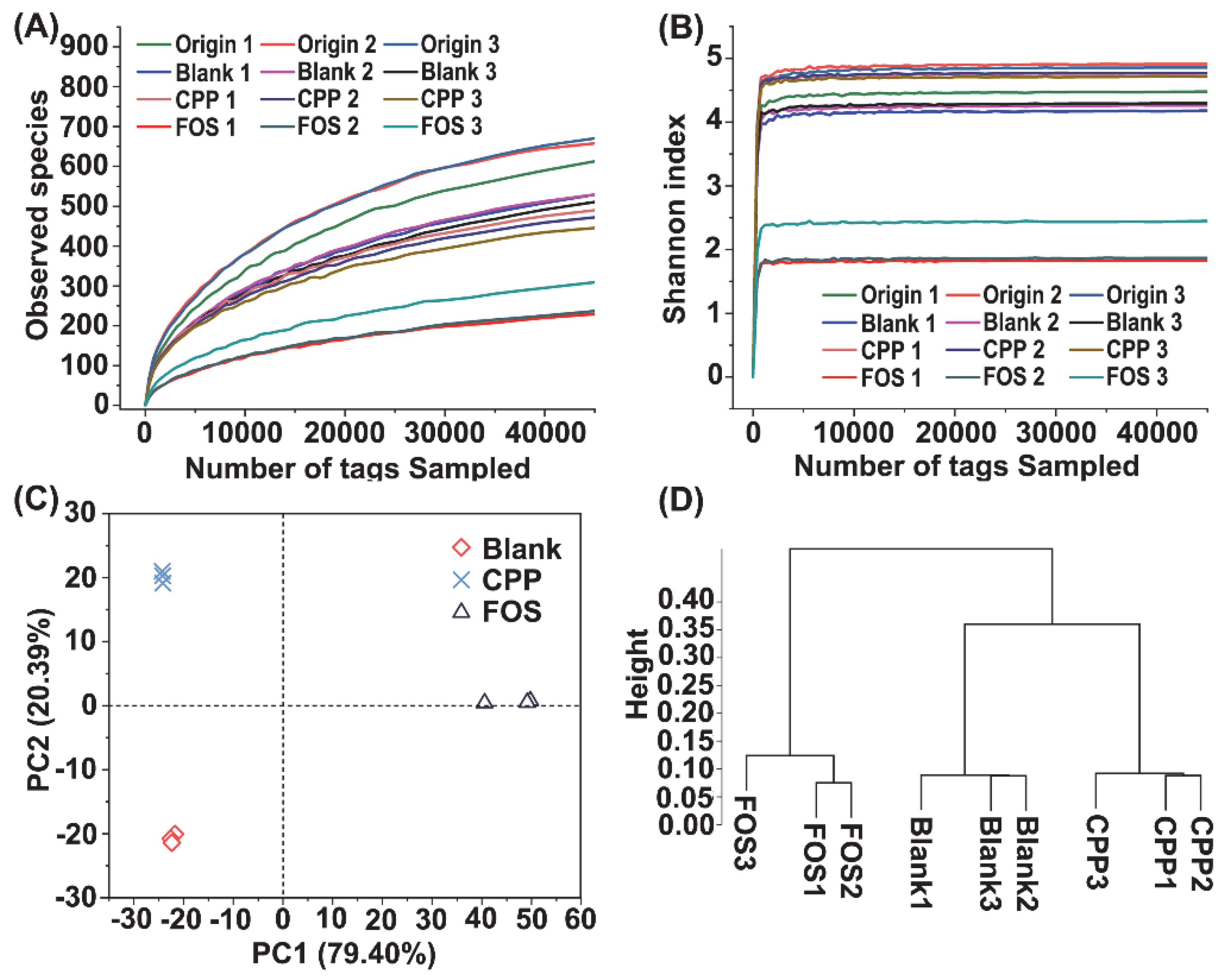
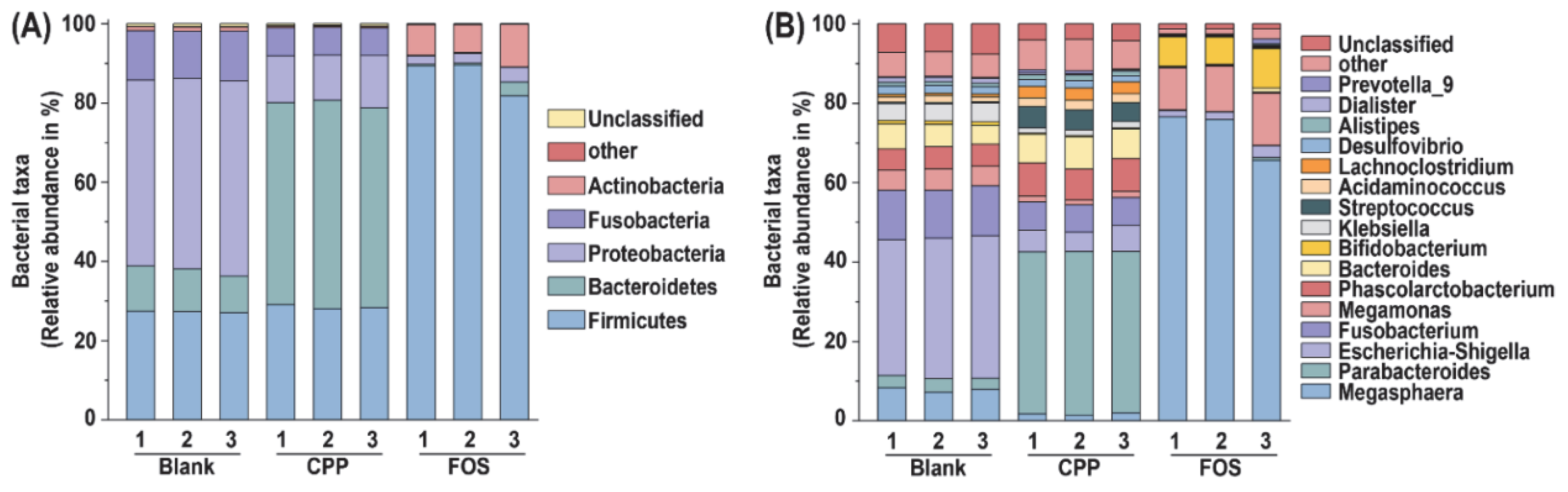
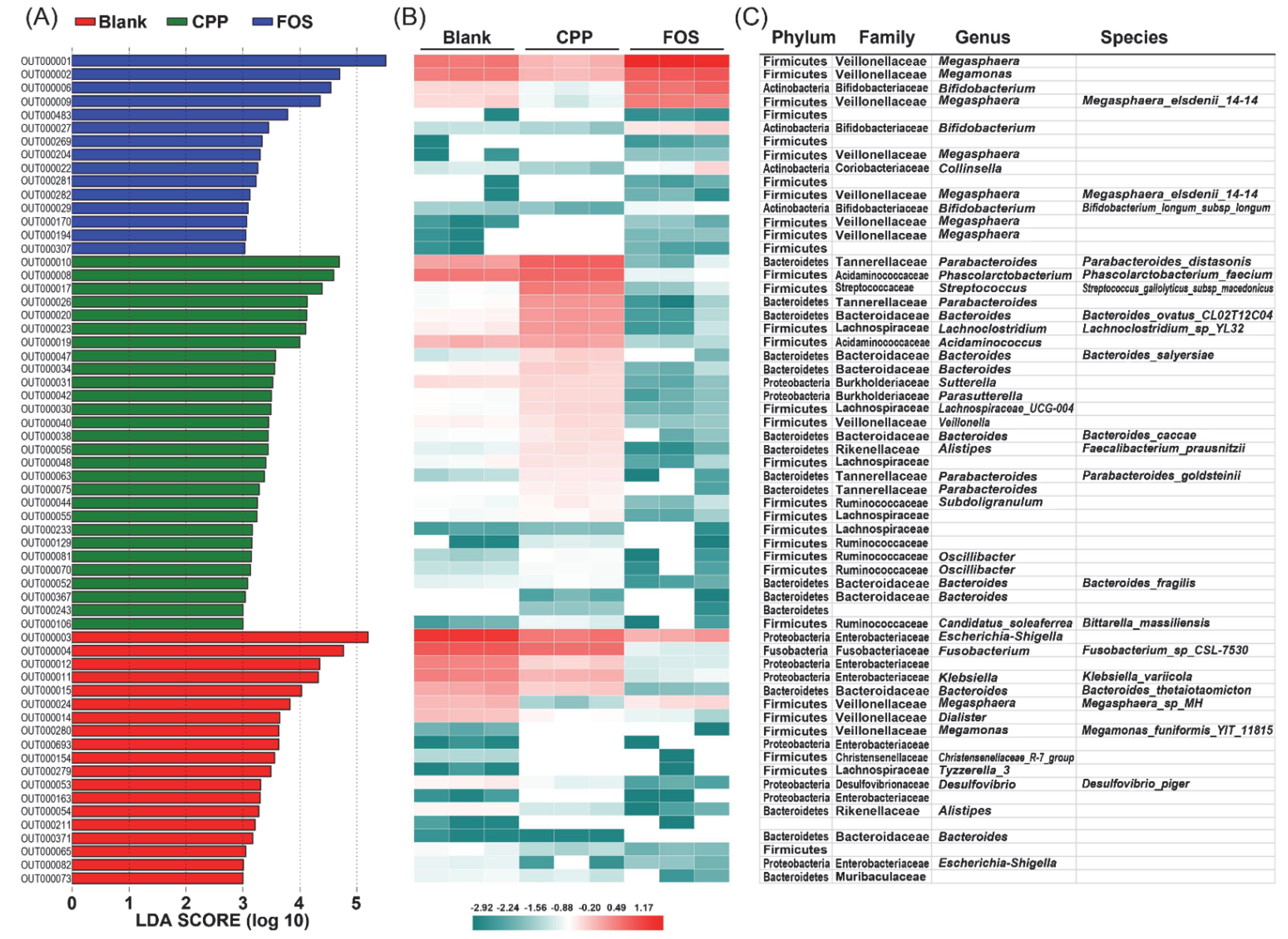
3.5. Effect of CPP on Gut Microbiota
3.6. Effect of CPP on SCFAs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a future food source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Horii, N.; Hasegawa, N.; Fujie, S.; Uchida, M.; Iemitsu, K.; Inoue, K.; Iemitsu, M. Effect of combination of chlorella intake and aerobic exercise training on glycemic control in type 2 diabetic rats. Nutrition 2019, 63, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.X.; Li, H.; Wei, Z.Y.; Lv, K.L.; Gao, C.H.; Liu, Y.H.; Zhao, L.Y. Isolation, structures and biological activities of polysaccharides from Chlorella: A review. Int. J. Biol. Macromol. 2020, 163, 2199–2209. [Google Scholar] [CrossRef]
- Capek, P.; Matulová, M.; Šutovská, M.; Barboríková, J.; Molitorisová, M.; Kazimierová, I. Chlorella vulgaris α-L-arabino-α-L-rhamno-α, β-D-galactan structure and mechanisms of its anti-inflammatory and anti-remodelling effects. Int. J. Biol. Macromol. 2020, 162, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.M.; Shi, Y.; Sheng, J.C.; Hu, Q.H. In vivo immunomodulatory activity of polysaccharides derived from Chlorella pyrenoidosa. Eur. Food Res. Technol. 2006, 224, 225–228. [Google Scholar] [CrossRef]
- Wan, X.Z.; Ai, C.; Chen, Y.H.; Gao, X.X.; Zhong, R.T.; Liu, B.; Chen, X.H.; Zhao, C. Physicochemical characterization of a polysaccharide from green microalga Chlorella pyrenoidosa and its hypolipidemic activity via gut microbiota regulation in rats. J. Agric. Food Chem. 2020, 68, 1186–1197. [Google Scholar] [CrossRef]
- Salama, N.N.; Eddington, N.D.; Fasano, A. Tight junction modulation and its relationship to drug delivery. Adv. Drug Deliv. Rev. 2006, 58, 15–28. [Google Scholar] [CrossRef]
- Sato, Y.; Joumura, T.; Takekuma, Y.; Sugawara, M. Transfer of orally administered hyaluronan to the lymph. Eur. J. Pharm. Biopharm. 2020, 154, 210–213. [Google Scholar] [CrossRef]
- Zheng, Z.M.; Pan, X.L.; Wang, H.Y.; Wu, Z.J.; Sullivan, M.A.; Liu, Y.X.; Liu, J.X.; Wang, K.P.; Zhang, Y. Mechanism of lentinan intestinal absorption: Clathrin-mediated endocytosis and macropinocytosis. J. Agric. Food Chem. 2021, 69, 7344–7352. [Google Scholar] [CrossRef]
- Gallego-Lobillo, P.; Ferreira-Lazarte, A.; Hernández-Hernández, O.; Villamiel, M. In vitro digestion of polysaccharides: InfoGest protocol and use of small intestinal extract from rat. Food Res. Int. 2021, 140, 110054. [Google Scholar] [CrossRef]
- Fonseca, R.J.C.; Sucupira, I.D.; Oliveira, S.N.M.C.G.; Santos, G.R.C.; Mourão, P.A.S. Improved anticoagulant effect of fucosylated chondroitin sulfate orally administered as gastro-resistant tablets. Thromb. Haemost. 2017, 117, 662–670. [Google Scholar] [PubMed] [Green Version]
- Yuan, Y.Q.; Li, C.; Zheng, Q.W.; Wu, J.X.; Zhu, K.X.; Shen, X.R.; Cao, J. Effect of simulated gastrointestinal digestion in vitro on the antioxidant activity, molecular weight and microstructure of polysaccharides from a tropical sea cucumber (Holothuria leucospilota). Food Hydrocoll. 2019, 89, 735–741. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Qin, Y.J.; Guan, R.W.; Zheng, W.Q.; Liu, J.K.; Zhao, J.H. Digestibility of fucosylated glycosaminoglycan from sea cucumber and its effects on digestive enzymes under simulated salivary and gastrointestinal conditions. Carbohydr. Polym. 2018, 186, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Beheshtipour, H.; Mortazavian, A.M.; Mohammadi, R.; Sohrabvandi, S.; Khosravi-Daran, K. Supplementation of Spirulina platensis and Chlorella vulgaris algae into probiotic fermented milks. Compr. Rev. Food Sci. Food Saf. 2013, 12, 144–154. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Dodgston, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.X.; Zhang, J.; Xiao, C.L.; Harqin, C.L.G.; Ma, M.Y.; Long, T.; Li, Z.H.; Yang, Y.L.; Liu, J.K.; Zhao, L.Y. Structural characterization of a low-molecular-weight polysaccharide from Angelica pubescens Maxim. f. biserrata Shan et Yuan root and evaluation of its antioxidant activity. Carbohydr. Polym. 2020, 236, 116047. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Wu, M.Y.; Xiao, C.; Yang, L.; Zhou, L.T.; Gao, N.; Li, Z.; Chen, J.; Chen, J.C.; Liu, J.K.; et al. Discovery of an intrinsic tenase complex inhibitor: Pure nonasaccharide from fucosylated glycosaminoglycan. Proc. Natl. Acad. Sci. USA 2015, 112, 8284–8289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.L.; Chen, G.J.; Li, X.; Zheng, F.; Zeng, X.X. Yeast β-glucan, a potential prebiotic, showed a similar probiotic activity to inulin. Food Funct. 2020, 11, 10386–10396. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Jiang, S.S.; Zeng, M.Y. In vitro simulated digestion and fermentation characteristics of polysaccharide from oyster (Crassostrea gigas), and its effects on the gut microbiota. Food Res. Int. 2021, 149, 110646. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.T.; Yan, Y.M.; Mi, J.; Zhang, H.C.; Lu, L.; Luo, Q.; Li, X.Y.; Zeng, X.X.; Cao, Y.L. Simulated digestion and fermentation in vitro by human gut microbiota of polysaccharides from bee collected pollen of Chinese wolfberry. J. Agric. Food Chem. 2018, 66, 898–907. [Google Scholar] [CrossRef]
- Zhao, Z.X.; Ming, J.; Zhao, G.H.; Lei, L. Color, starch digestibility, and in vitro fermentation of roasted highland barley flour with different fractions. Foods 2022, 11, 287. [Google Scholar] [CrossRef]
- Chen, Y.X.; Liu, X.Y.; Wu, L.X.; Tong, A.J.; Zhao, L.N.; Liu, B.; Zhao, C. Physicochemical characterization of polysaccharides from Chlorella pyrenoidosa and its anti-ageing effects in Drosophila melanogaster. Carbohydr. Polym. 2018, 185, 120–126. [Google Scholar] [CrossRef]
- Chen, P.; Chen, X.Q.; Hao, L.L.; Du, P.; Li, C.; Han, H.Y.; Xu, H.X.; Liu, L.B. The bioavailability of soybean polysaccharides and their metabolites on gut microbiota in the simulator of the human intestinal microbial ecosystem (SHIME). Food Chem. 2021, 362, 130233. [Google Scholar] [CrossRef]
- Liu, Y.T.; Duan, X.Y.; Duan, S.Q.; Li, C.; Hu, B.; Liu, A.P.; Wu, Y.L.; Wu, H.J.; Chen, H.; Wu, W.J.; et al. Effects of in vitro digestion and fecal fermentation on the stability and metabolic behavior of polysaccharides from Craterellus cornucopioides. Food Funct. 2020, 11, 6899–6910. [Google Scholar] [CrossRef]
- Chen, P.L.; Lei, S.Z.; Chen, Y.Y.; Chang, Q.; Zhang, Y.; Hu, X.K.; Zeng, H.L. Structural characterization of a novel galactoglucan from Fortunella margarita and its molecular structural change following simulated digestion in vitro. J. Funct. Foods 2020, 71, 104024. [Google Scholar] [CrossRef]
- Chen, G.J.; Xie, M.H.; Wan, P.; Chen, D.; Ye, H.; Chen, L.G.; Zeng, X.X. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from Fuzhuan brick tea. Food Chem. 2018, 244, 331–339. [Google Scholar] [CrossRef]
- Ma, G.X.; Xu, Q.; Du, H.J.; Kimatu, M.B.; Su, A.X.; Yang, W.J.; Hu, Q.H.; Xiao, H. Characterization of polysaccharide from Pleurotus eryngii during simulated gastrointestinal digestion and fermentation. Food Chem. 2022, 370, 131303. [Google Scholar] [CrossRef] [PubMed]
- Payling, L.; Fraser, K.; Loveday, S.M.; Sims, I.; Roy, N.; McNabb, W. The effects of carbohydrate structure on the composition and functionality of the human gut microbiota. Trends Food Sci. Technol. 2020, 97, 233–248. [Google Scholar] [CrossRef]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.L.; Zhu, Y.M.; Shao, D.Y.; Zhao, K.; Xu, C.L.; Li, Q.; Yang, H.; Huang, Q.S.; Shi, J.L. Effects of polysaccharide from mycelia of Ganoderma lucidum on intestinal barrier functions of rats. Int. J. Biol. Macromol. 2017, 94, 1–9. [Google Scholar] [CrossRef]
- Ke, S.Z.; Yu, Y.L.; Xu, Q.L.; Zhang, B.; Wang, S.J.; Jin, W.H.; Wei, B.; Wang, H. Composition-activity relationships of polysaccharides from Saccharina japonicain regulating gut microbiota in short-term high-fat diet-fed mice. J. Agric. Food Chem. 2021, 69, 11121–11130. [Google Scholar] [CrossRef]
- Ferrari, M.L.; Malardé, V.; Grassart, A.; Salavessa, L.; Nigro, G.; Descorps-Declere, S.; Rohde, J.R.; Schnupf, P.; Masson, V.; Arras, G.; et al. Shigella promotes major alteration of gut epithelial physiology and tissue invasion by shutting off host intracellular transport. Proc. Natl. Acad. Sci. USA 2019, 116, 13582–13591. [Google Scholar] [CrossRef] [Green Version]
- Serapio-Palacios, A.; Finlay, B.B. Dynamics of expression, secretion and translocation of type III effectors during enteropathogenic Escherichia coli infection. Curr. Opin. Microbiol. 2020, 54, 67–76. [Google Scholar] [CrossRef]
- Olm, M.R.; Bhattacharya, N.; Crits-Christoph, A.; Firek, B.A.; Baker, R.; Song, Y.S.; Morowitz, M.J.; Banfield, J.F. Necrotizing enterocolitis is preceded by increased gut bacterial replication, Klebsiella, and fimbriae-encoding bacteria. Sci. Adv. 2019, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hashizume, K.; Tsukahara, T.; Yamada, K.; Koyama, H.; Ushida, K. Megasphaera elsdenii JCM1772T normalizes hyperlactate production in the large intestine of fructooligosaccharide-fed rats by stimulating butyrate production. J. Nutr. 2003, 133, 3187–3190. [Google Scholar] [CrossRef] [Green Version]
- Kienbacher, C.; Traussnigg, S.; Halilbasic, E.; Fuchs, C.D.; Dolak, W.; Munda, P.; Stojakovic, T.; Fauler, G.; Forster, M.; Heinsen, F.A.; et al. Bloom of fecal Megamonas after a 4 week high oral fructose challenge disturbs energy and lipid metabolism: Linking diet to microbiota, bile acid and host metabolism alterations. Gastroenterology 2016, 150, S1056–S1057. [Google Scholar] [CrossRef]
- Wang, F.; Yin, Q.; Chen, L.; Davis, M.M. Bifidobacterium can mitigate intestinal immunopathology in the context of CTLA-4 blockade. Proc. Natl. Acad. Sci. USA 2018, 115, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Liao, M.F.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.Y.; Wang, Y.J.; Liu, C.; Wang, W.Z.; Wang, J.; et al. Parabacteroides distasonis alleviates obesity and metabolic dysfunctions via production of succinate and secondary bile acids. Cell Rep. 2019, 26, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef]
- Del Dot, T.; Osawa, R.; Stackebrandt, E. Phascolarctobacterium faecium gen. nov, spec. nov., a novel taxon of the Sporomusa group of bacteria. Syst. Appl. Microbiol. 1993, 16, 380–384. [Google Scholar] [CrossRef]
- Wu, F.F.; Guo, X.F.; Zhang, J.C.; Zhang, M.; Ou, Z.H.; Peng, Y.Z. Phascolarctobacterium faecium abundant colonization in human gastrointestinal tract. Exp. Ther. Med. 2017, 14, 3122–3126. [Google Scholar] [CrossRef] [Green Version]
- Breyner, N.M.; Michon, C.; de Sousa, C.S.; Vilas Boas, P.B.; Chain, F.; Azevedo, V.A.; Langella, P.; Chatel, J.M. Microbial anti-inflammatory molecule (MAM) from Faecalibacterium prausnitzii shows a protective effect on DNBS and DSS-induced colitis model in mice through inhibition of NF-κB pathway. Front. Microbiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhao, L.P.; Zhang, M.H. Gut microbial SNPs induced by high-fiber diet dominate nutrition metabolism and environmental adaption of Faecalibacterium prausnitzii in obese children. Front. Microbiol. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Quévrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B.; et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.T.; Yuan, Q.; Guo, H.; Fu, Y.; Li, F.; Wang, S.P.; Gan, R.Y. Dynamic changes of structural characteristics of snow chrysanthemum polysaccharides during in vitro digestion and fecal fermentation and related impacts on gut microbiota. Food Res. Int. 2021, 141, 109888. [Google Scholar] [CrossRef] [PubMed]
- Al-Lahham, S.H.; Peppelenbosch, M.P.; Roelofsen, H.; Vonk, R.J.; Venema, K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Bourriaud, C.; Robins, R.J.; Martin, L.; Kozlowski, F.; Tenailleau, E.; Cherbut, C.; Michel, C. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 2005, 99, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.Y.; Tang, L.; Liu, S.; Hu, S.P.; Wu, L.Y.; Liu, Y.J.; Yang, M.; Huang, S.J.; Tang, X.; Tang, T.; et al. Parabacteroides produces acetate to alleviate heparanase-exacerbated acute pancreatitis through reducing neutrophil infiltration. Microbiome 2021, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.D.; Liu, Z.M.; Zhu, C.L.; Mou, H.J.; Kong, Q. Nondigestible carbohydrates, butyrate, and butyrate-producing bacteria. Crit. Rev. Food Sci. Nutr. 2019, 59, S130–S152. [Google Scholar] [CrossRef]
- Yu, C.X.; Wu, D.M.; Zhu, K.; Hou, L.J.; Xiao, H.; Ding, T.; Liu, D.H.; Ye, X.Q.; Linhardt, R.J.; Chen, S.G. Challenges of pectic polysaccharides as a prebiotic from the perspective of fermentation characteristics and anti-colitis activity. Carbohydr. Polym. 2021, 270, 118377. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Time (h) | Residual Carbohydrates (%) | Reducing Sugars (%) |
|---|---|---|
| 0 | 100.00 ± 2.39 a | 100.00 ± 2.80 a |
| 6 | 91.74 ± 4.96 b | 76.61 ± 2.88 b |
| 12 | 76.20 ± 5.19 c | 75.00 ± 3.75 b |
| 24 | 65.86 ± 3.55 d | 54.55 ± 0.66 c |
| 48 | 44.20 ± 2.65 e | 28.61 ± 0.57 d |
| Group | Index | |||
|---|---|---|---|---|
| Chao1 | ACE | Shannon | Simpson | |
| Origin | 705.71 ± 25.71 a | 741.54 ± 28.05 a | 4.75 ± 0.24 a | 0.87 ± 0.00 a |
| Blank | 634.39 ± 8.05 b | 662.43 ± 14.01 a | 4.25 ± 0.06 a | 0.85 ± 0.01 a |
| CPP | 549.30 ± 20.05 c | 575.29 ± 23.51 b | 4.75 ± 0.03 a | 0.91 ± 0.00 a |
| FOS | 350.98 ± 39.66 d | 369.21 ± 48.91 c | 2.05 ± 0.34 b | 0.53 ± 0.07 b |
| SCFAs (mM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group | Time (h) | Lactic Acid | Acetic Acid | Propionic Acid | i-Butyric Acid | n-Butyric Acid | i-Valeric Acid | n-Valeric Acid | Total SCFAs |
| Blank | 0 | ND | 0.973 ± 0.026 a,A | 0.681 ± 0.006 a,A | ND | 0.565 ± 0.006 a,A | ND | 0.601 ± 0.000 a,A | 2.819 ± 0.037 a,A |
| 6 | 0.557 ± 0.000 a,A | 3.460 ± 0.243 b,A | 2.327 ± 0.124 b,A | ND | 0.667 ± 0.012 a,C | 0.147 ± 0.006 a,A | 0.655 ± 0.004 a,C | 7.812 ± 0.363 b,A | |
| 12 | ND | 7.566 ± 0.133 c,A | 3.517 ± 0.081 d,A | 0.448 ± 0.042 a | 1.481 ± 0.089 b,C | 1.510 ± 0.040 b,A | 1.063 ± 0.034 b,C | 15.585 ± 0.380 c,B | |
| 24 | ND | 8.531 ± 0.416 e,A | 3.483 ± 0.136 d,A | 1.055 ± 0.036 b,B | 2.160 ± 0.045 d,B | 2.130 ± 0.066 d,B | 1.940 ± 0.054 c,B | 19.300 ± 0.736 d,A | |
| 48 | ND | 8.309 ± 0.449 d,B | 2.836 ± 0.276 c,B | 0.945 ± 0.066 b,B | 1.916 ± 0.112 c,A | 1.907 ± 0.100 c,B | 2.059 ± 0.117 c,A | 17.971 ± 1.107 d,A | |
| CPPs | 0 | ND | 0.973 ± 0.026 a,A | 0.681 ± 0.006 a,A | ND | 0.565 ± 0.006 a,A | ND | 0.601 ± 0.000 a,A | 2.819 ± 0.037 a,A |
| 6 | 1.493 ± 0.045 c,B | 5.488 ± 0.440 b,B | 2.831 ± 0.134 b,B | ND | 0.601 ± 0.003 ab,A | 0.110 ± 0.000 a,B | 0.605 ± 0.000 a,A | 11.174 ± 0.566 b,B | |
| 12 | 0.127 ± 0.001 a,A | 6.780 ± 0.500 c,A | 5.288 ± 0.412 c,B | ND | 0.679 ± 0.006 b,A | 0.160 ± 0.000 a,A | 0.616 ± 0.003 a,A | 13.649 ± 0.219 c,A | |
| 24 | 0.333 ± 0.016 b | 15.463 ± 0.383 d,C | 8.681 ± 0.174 d,B | 0.306 ± 0.048 a,A | 1.616 ± 0.084 c,A | 0.817 ± 0.284 b,A | 0.732 ± 0.125 a,A | 27.938 ± 0.869 d,C | |
| 48 | 0.362 ± 0.014 b | 18.968 ± 0.302 e,C | 9.617 ± 0.158 e,C | 1.009 ± 0.019 a,B | 2.357 ± 0.029 d,B | 1.847 ± 0.025 c,B | 1.917 ± 0.007 b,A | 36.076 ± 0.272 e,C | |
| FOS | 0 | ND | 0.973 ± 0.026 a,A | 0.681 ± 0.006 a,A | ND | 0.565 ± 0.006 a,A | ND | 0.601 ± 0.000 a,A | 2.819 ± 0.037 a,A |
| 6 | 5.567 ± 0.022 a,C | 7.523 ± 0.188 c,C | 5.688 ± 0.236 d,C | ND | 0.641 ± 0.010 ab,B | ND | 0.623 ± 0.002 a,B | 20.041 ± 0.387 b,C | |
| 12 | 5.314 ± 0.033 a,B | 13.367 ± 0.292 e,B | 7.076 ± 0.010 e,C | ND | 0.955 ± 0.016 b,B | ND | 0.700 ± 0.001 a,B | 27.422 ± 0.320 d,C | |
| 24 | ND | 10.052 ± 0.025 d,B | 3.666 ± 0.020 c,A | 0.255 ± 0.004 a,A | 6.372 ± 0.071 c,C | 0.950 ± 0.044 a,A | 3.778 ± 0.039 b,C | 25.074 ± 0.073 c,B | |
| 48 | ND | 4.580 ± 0.364 b,A | 1.335 ± 0.054 b,A | 0.557 ± 0.018 a,A | 11.611 ± 0.259 d,C | 1.353 ± 0.045 a,A | 7.200 ± 0.196 c,B | 26.637 ± 0.832 d,B | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, K.; Yuan, Q.; Li, H.; Li, T.; Ma, H.; Gao, C.; Zhang, S.; Liu, Y.; Zhao, L. Chlorella pyrenoidosa Polysaccharides as a Prebiotic to Modulate Gut Microbiota: Physicochemical Properties and Fermentation Characteristics In Vitro. Foods 2022, 11, 725. https://doi.org/10.3390/foods11050725
Lv K, Yuan Q, Li H, Li T, Ma H, Gao C, Zhang S, Liu Y, Zhao L. Chlorella pyrenoidosa Polysaccharides as a Prebiotic to Modulate Gut Microbiota: Physicochemical Properties and Fermentation Characteristics In Vitro. Foods. 2022; 11(5):725. https://doi.org/10.3390/foods11050725
Chicago/Turabian StyleLv, Kunling, Qingxia Yuan, Hong Li, Tingting Li, Haiqiong Ma, Chenghai Gao, Siyuan Zhang, Yonghong Liu, and Longyan Zhao. 2022. "Chlorella pyrenoidosa Polysaccharides as a Prebiotic to Modulate Gut Microbiota: Physicochemical Properties and Fermentation Characteristics In Vitro" Foods 11, no. 5: 725. https://doi.org/10.3390/foods11050725
APA StyleLv, K., Yuan, Q., Li, H., Li, T., Ma, H., Gao, C., Zhang, S., Liu, Y., & Zhao, L. (2022). Chlorella pyrenoidosa Polysaccharides as a Prebiotic to Modulate Gut Microbiota: Physicochemical Properties and Fermentation Characteristics In Vitro. Foods, 11(5), 725. https://doi.org/10.3390/foods11050725