
Freeze-Thawing Treatment as a Simple Way to Tune the Gel Property and Digestibility of Minced Meat from Red Swamp Crayfish (Procambarus clarkiix)

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Freezing and Thawing Treatments
2.3. Crayfish Meat Collection, Meat Yield, and Drip Loss Measurement
2.4. Gel Preparation
2.5. Mechanical Properties and Color Analysis of Gels
2.6. Water-Holding Capacity Determination and SEM Observation
2.7. Ultraviolet (UV) Spectrum and Fourier Transform Infrared Spectroscopy (FTIR) Analysis
2.8. In Vitro Gastrointestinal Digestion and Antioxidant Activity of Crayfish Meat Gels
2.9. Protein Patterns of Gels before and after Digestion
2.10. Statistical Analysis
3. Results and Discussion
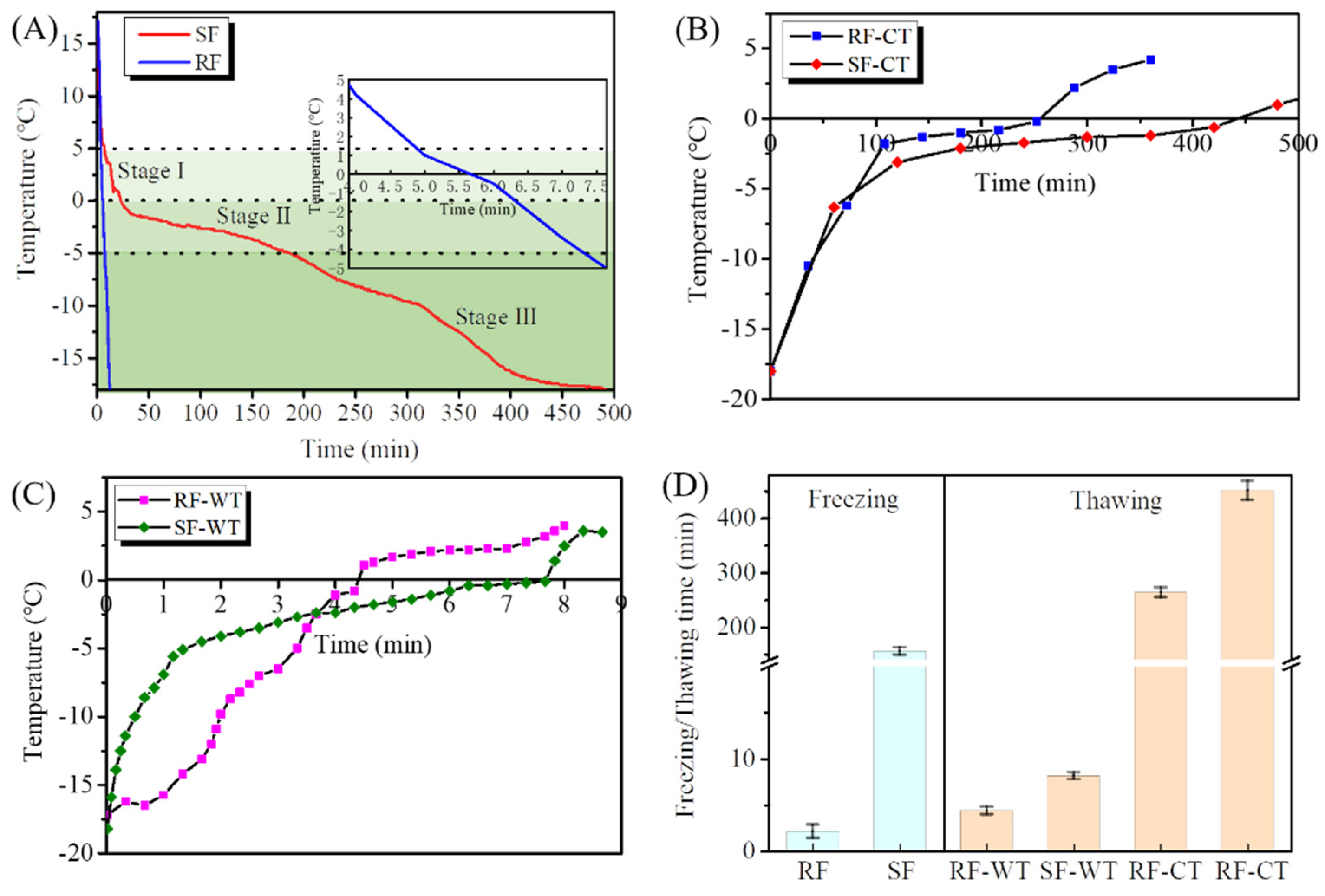
3.1. Freezing and Thawing Curves of Crayfish
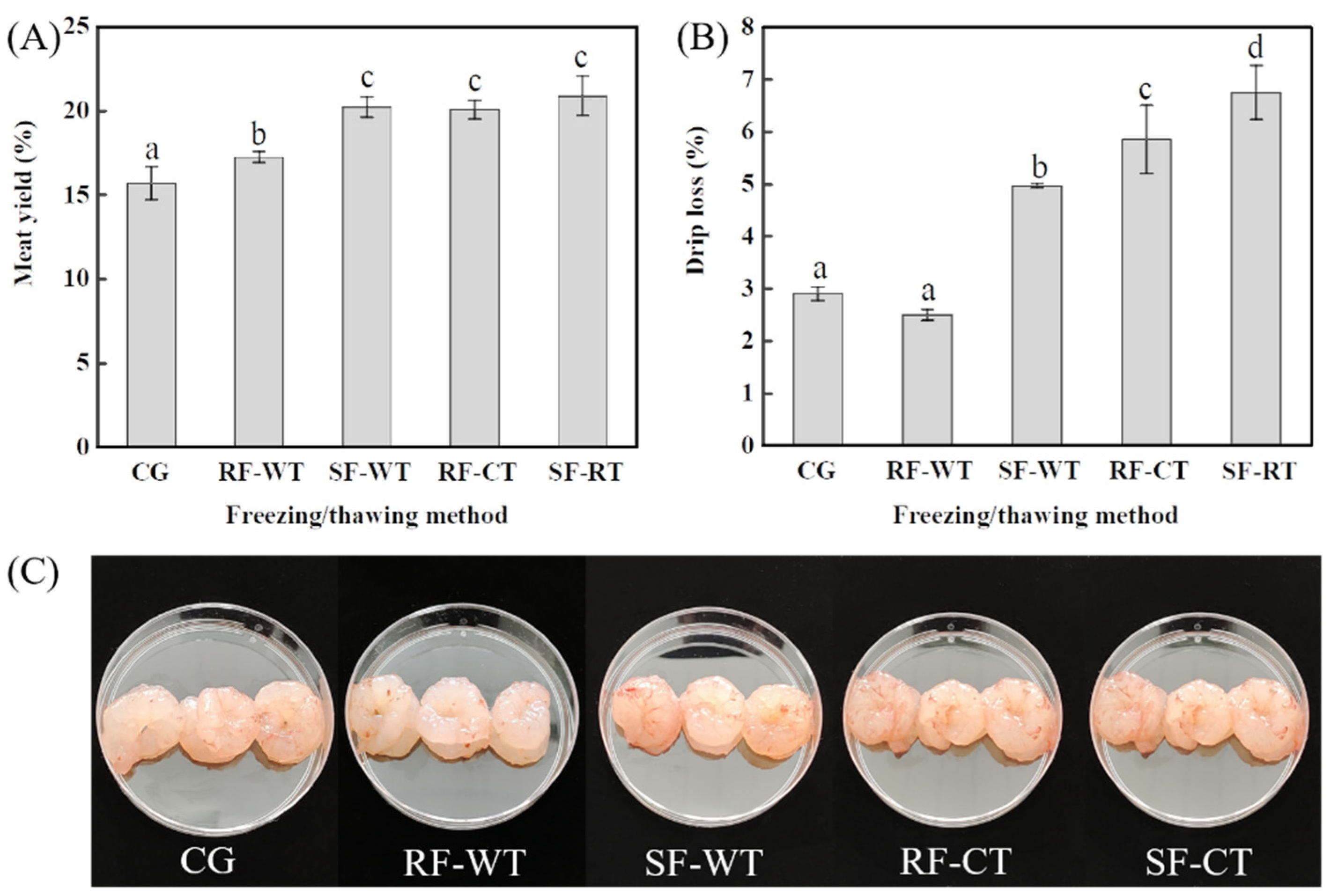
3.2. Meat Yield and Drip Loss of Crayfish Meat
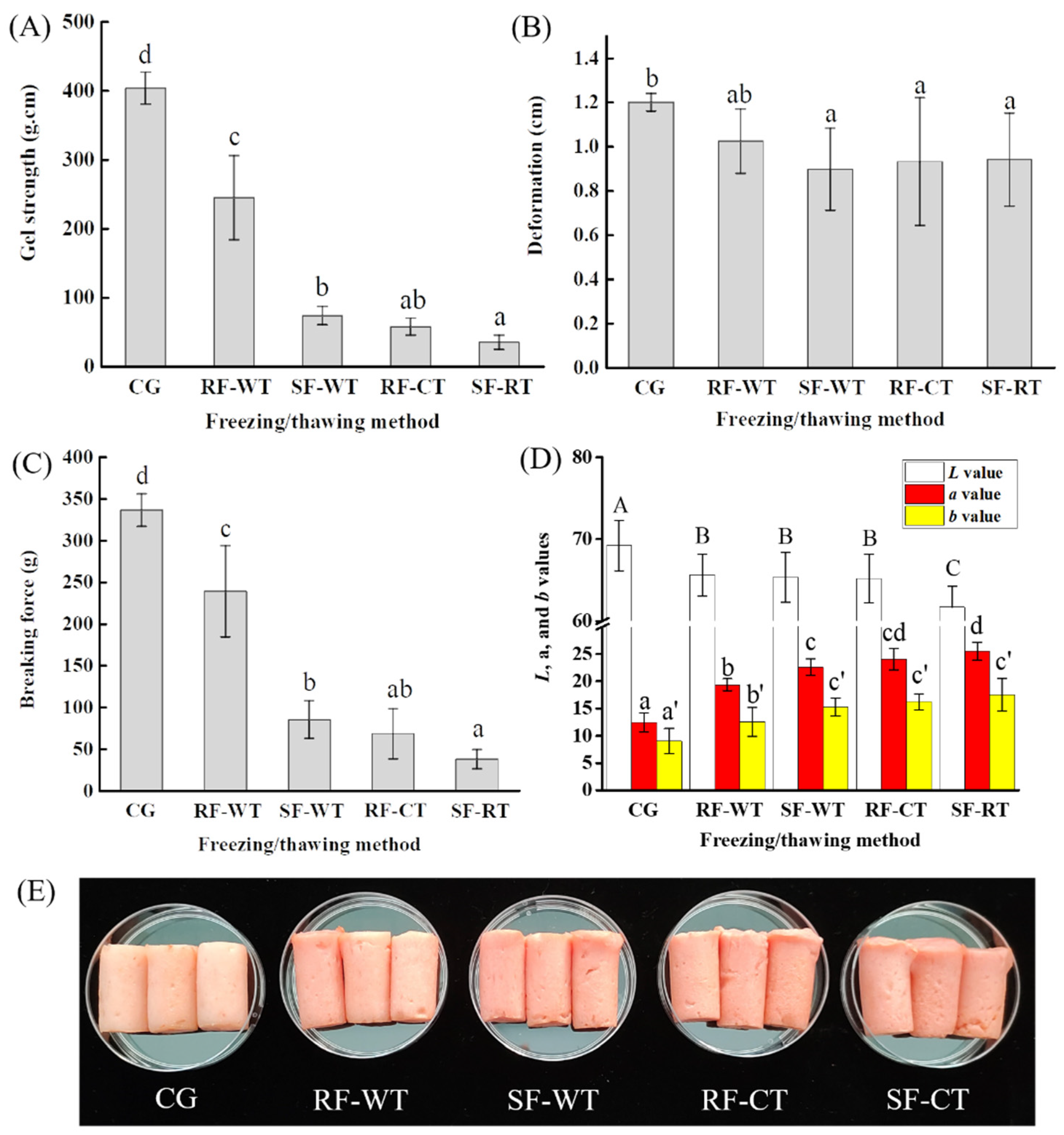
3.3. Texture and Color of Crayfish Meat Gels
3.4. WHC and SEM of Gels
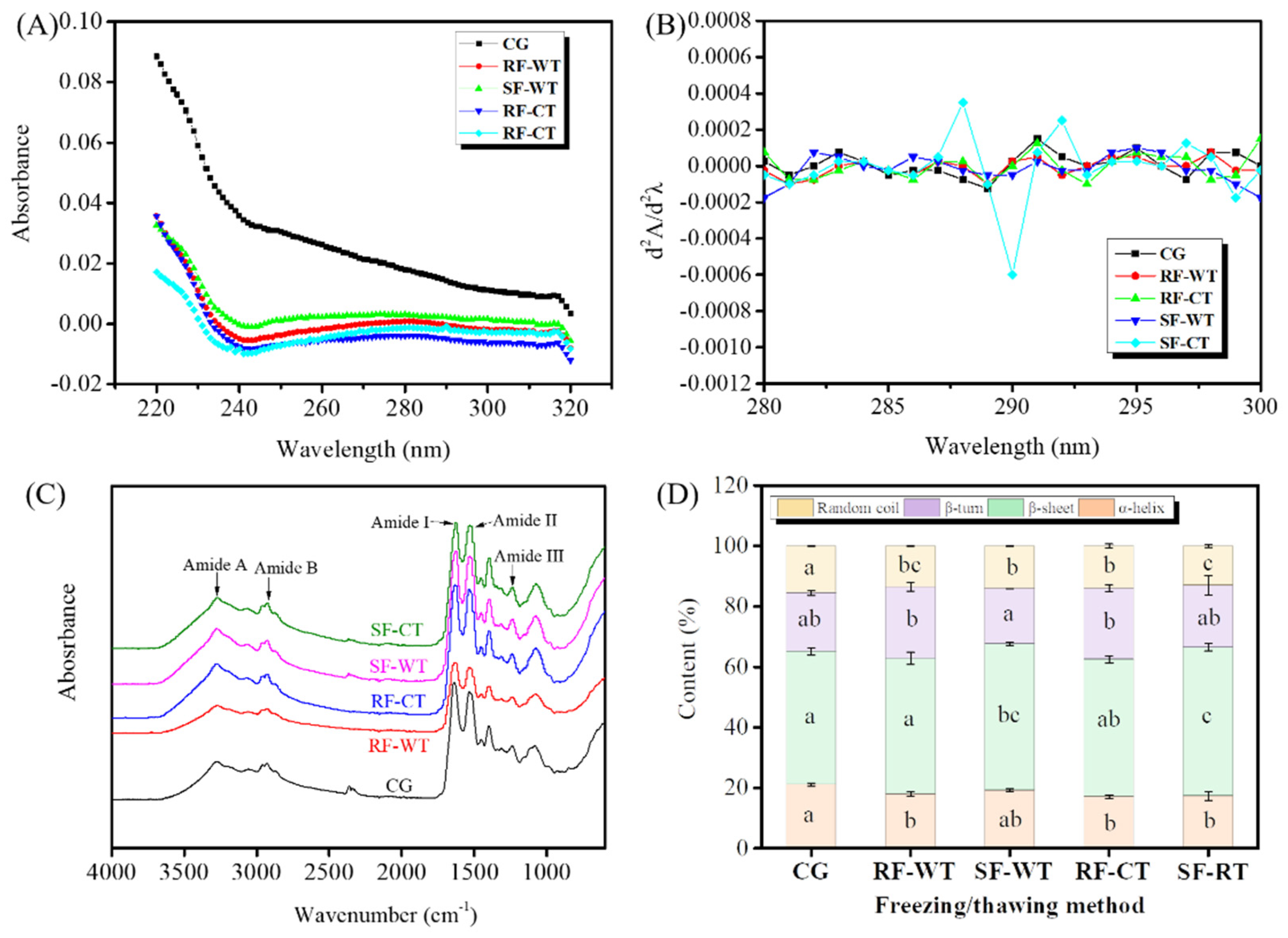
3.5. UV Spectrum and FTIR
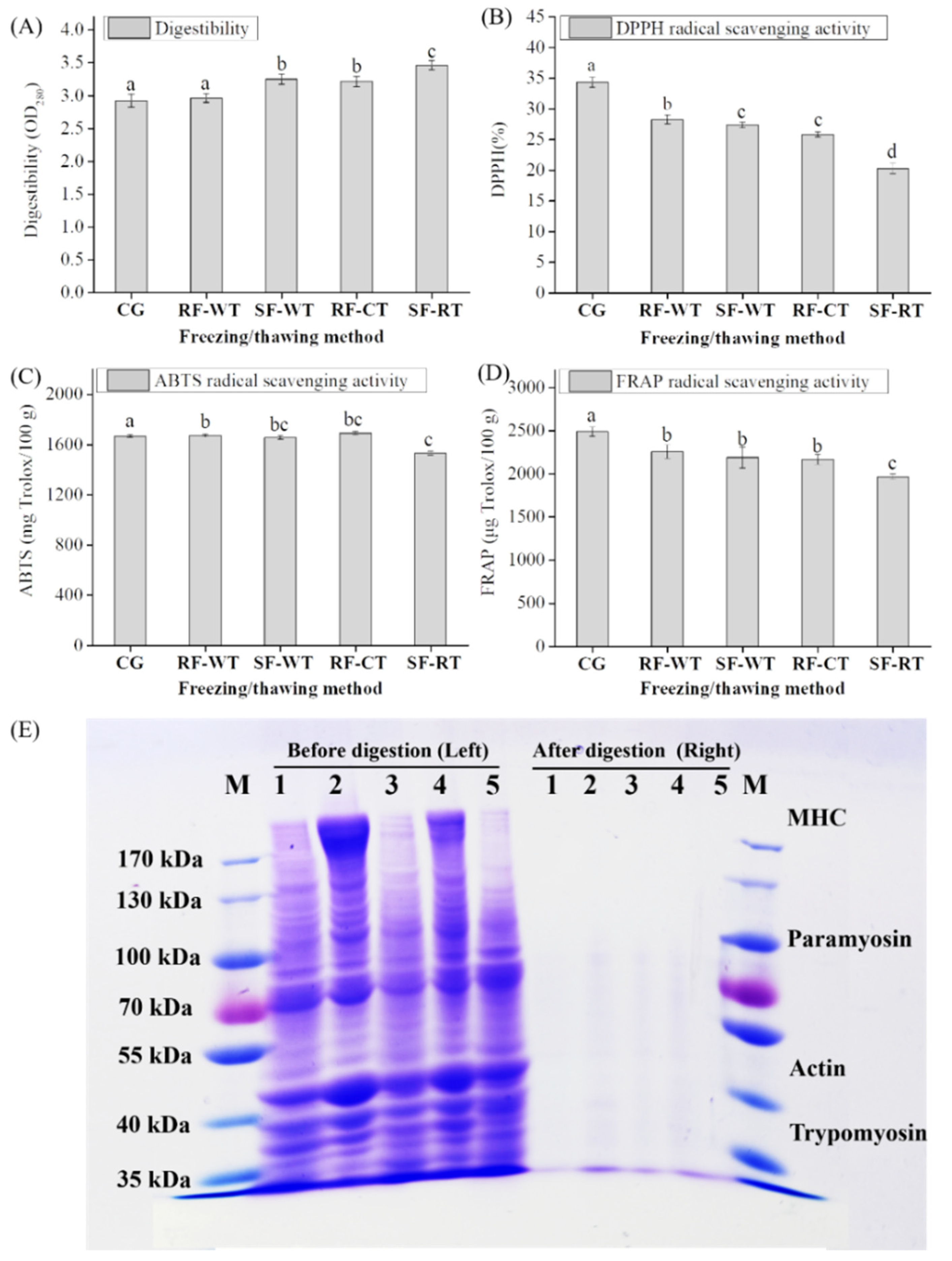
3.6. Digestibility, Antioxidant Activity, and SDS-PAGE
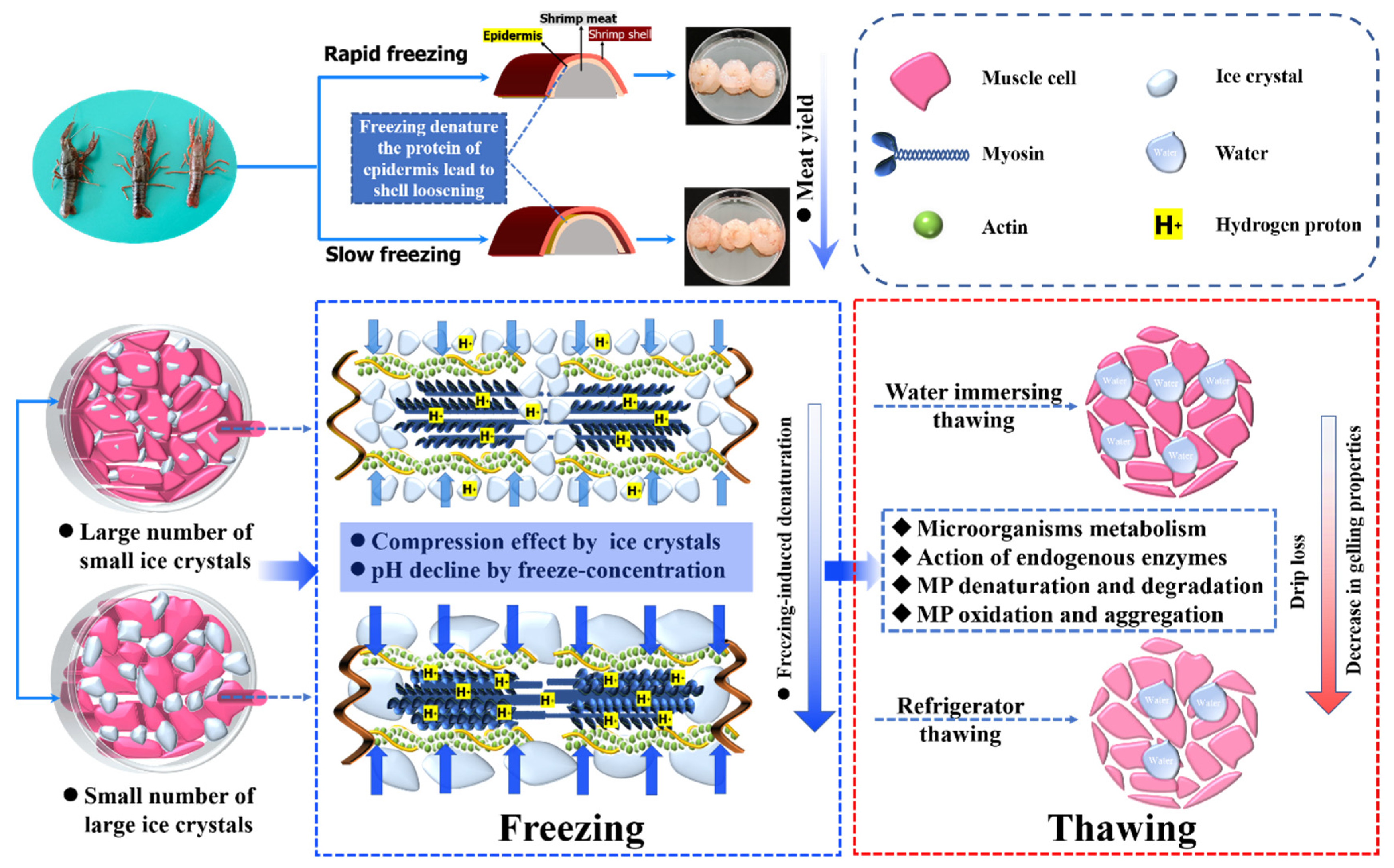
3.7. Schematic Model
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bao, Y.; Ertbjerg, P.; Estévez, M.; Yuan, L.; Gao, R. Freezing of meat and aquatic food: Underlying mechanisms and implications on protein oxidation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5548–5569. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, N.P.; Benjakul, S. Effect of catechin and ferulic acid on melanosis and quality of Pacific white shrimp subjected to prior freez—thawing during refrigerated storage. Food Control 2010, 21, 1263–1271. [Google Scholar] [CrossRef]
- Shi, L.; Xiong, G.; Ding, A.; Li, X.; Wu, W.; Qiao, Y.; Liao, L.; Wang, L. Effects of freezing temperature and frozen storage on the biochemical and physical properties of Procambarus clarkii. Int. J. Refrig. 2018, 91, 223–229. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, F.; Yao, Y.; He, J.; Tang, J.; Jiao, Y. Effects of freeze-thaw cycles of Pacific white shrimp (Litopenaeus vannamei) subjected to radio frequency tempering on melanosis and quality. Innov. Food Sci. Emerg. Technol. 2021, 74, 102860. [Google Scholar] [CrossRef]
- Tan, M.; Ye, J.; Xie, J. Freezing-induced myofibrillar protein denaturation: Role of pH change and freezing rate. LWT Food Sci. Technol. 2021, 152, 112381. [Google Scholar] [CrossRef]
- Li, F.; Wang, B.; Kong, B.; Shi, S.; Xia, X. Decreased gelling properties of protein in mirror carp (Cyprinus carpio) are due to protein aggregation and structure deterioration when subjected to freeze-thaw cycles. Food Hydrocoll. 2019, 97, 105223. [Google Scholar] [CrossRef]
- Dang, T.T.; Gringer, N.; Jessen, F.; Olsen, K.; Bøknæs, N.; Nielsen, P.L.; Orlien, V. Emerging and potential technologies for facilitating shrimp peeling: A review. Innov. Food Sci. Emerg. Technol. 2018, 45, 228–240. [Google Scholar] [CrossRef]
- Yi, J.; Ding, G.; Hu, X.; Dong, P.; Li, Y.; Liao, X.; Zhang, Y. Comparison of shucking techniques for white shrimp and its effect on quality of peeled shrimp. Trans. Chin. Soc. Agr. Eng. 2012, 28, 287–292. [Google Scholar] [CrossRef]
- Shi, L.; Li, X.; Xiong, G.; Wu, W.; Ding, A.; Qiao, Y.; Liao, L.; Chen, D.; Wang, L. Correlation analysis of microstructure, protein pattern, and thermal properties of Procambarus clarkia subjected to different cryogenic treatments. Food Sci. Nutr. 2021, 9, 4946–4951. [Google Scholar] [CrossRef]
- Yearbook, C.F.S. China Fishery Statistic Yearbook; China Agriculture Press: Beijing, China, 2021; pp. 98–102. [Google Scholar]
- Shi, L.; Xiong, G.; Yin, T.; Ding, A.; Li, X.; Wu, W.; Qiao, Y.; Liao, L.; Jiao, C.; Wang, L. Effects of ultra-high pressure treatment on the protein denaturation and water properties of red swamp crayfish (Procambarus clarkia). LWT—Food Sci. Technol. 2020, 133, 110–124. [Google Scholar] [CrossRef]
- Li, X.; Li, S.; Shi, G.; Xiong, G.; Shi, L.; Kang, J.; Su, J.; Ding, A.; Li, X.; Qiao, Y.; et al. Quantitative proteomics insights into gel properties changes of myofibrillar protein from Procambarus clarkii under cold stress. Food Chem. 2021, 372, 130935. [Google Scholar] [CrossRef]
- Li, F.; Wang, B.; Liu, Q.; Chen, Q.; Zhang, H.; Xia, X.; Kong, B. Changes in myofibrillar protein gel quality of porcine longissimus muscle induced by its stuctural modification under different thawing methods. Meat Sci. 2019, 147, 108–115. [Google Scholar] [CrossRef]
- Liang, F.; Lin, L.; He, T.; Zhou, X.; Jiang, S.; Lu, J. Effect of transglutaminase on gel properties of surimi and precocious Chinese mitten crab (Eriocheir sinensis) meat. Food Hydrocoll. 2020, 98, 105261. [Google Scholar] [CrossRef]
- Yi, S.; Li, Q.; Qiao, C.; Zhang, C.; Wang, W.; Xu, Y.; Mi, H.; Li, X.; Li, J. Myofibrillar protein conformation enhance gel properties of mixed surimi gels with Nemipterus virgatus and Hypophthalmichthys molitrix. Food Hydrocoll. 2020, 106, 105924. [Google Scholar] [CrossRef]
- Singh, A.; Prabowo, F.F.; Benjakul, S.; Pranoto, Y.; Chantakun, K. Combined effect of microbial transglutaminase and ethanolic coconut husk extract on the gel properties and in-vitro digestibility of spotted golden goatfish (Parupeneus heptacanthus) surimi gel. Food Hydrocoll. 2020, 109, 106107. [Google Scholar] [CrossRef]
- Okita, A.; Takahashi, K.; Itakura, M.; Horio, A.; Yamamoto, R.; Nakamura, Y.; Osako, K. A novel soft surimi gel with functionality prepared using alcalase for people suffering from dysphagia. Food Chem. 2021, 344, 128641. [Google Scholar] [CrossRef]
- Li, Y.; Xiong, S.; Yin, T.; Hu, Y.; You, J. The gastric digestion kinetics of silver carp (Hypophthalmichthys molitrix) surimi gels induced by transglutaminase. Food Chem. 2019, 283, 148–154. [Google Scholar] [CrossRef]
- Xia, W. Food Technology; China Light Industry Press: Beijing, China, 2007; pp. 215–221. [Google Scholar]
- Tan, M.; Ding, Z.; Mei, J.; Xie, J. Effect of cellobiose on the myofibrillar protein denaturation induced by pH changes during freeze-thaw cycles. Food Chem. 2022, 373, 131511. [Google Scholar] [CrossRef]
- Lorentzen, G.; Hustad, A.; Lian, F.; Grip, A.E.; Schrødter, E.; Medeiros, T.; Siikavuopio, S.I. Effect of freezing methods, frozen storage time, and thawing methods on the quality of mildly cooked snow crab (Chionoecetes opilio) clusters. LWT—Food Sci. Technol. 2020, 123, 109103. [Google Scholar] [CrossRef]
- Gao, W.; Huang, Y.; Zeng, X.-A.; Brennan, M.A. Effect of soluble soybean polysaccharides on freeze-denaturation and structure of myofibrillar protein of bighead carp surimi with liquid nitrogen freezing. Int. J. Biol. Macromol. 2019, 135, 839–844. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, P.; Zhu, Z.; Sun, D.-W. Development of a single/dual-frequency orthogonal ultrasound-assisted rapid freezing technique and its effects on quality attributes of frozen potatoes. J. Food Eng. 2020, 286, 110112. [Google Scholar] [CrossRef]
- Li, D.; Zhao, H.; Muhammad, A.I.; Song, L.; Guo, M.; Liu, D. The comparison of ultrasound-assisted thawing, air thawing and water immersion thawing on the quality of slow/fast freezing bighead carp (Aristichthys nobilis) fillets. Food Chem. 2020, 320, 126614. [Google Scholar] [CrossRef]
- Lan, W.; Zhao, Y.; Gong, T.; Mei, J.; Xie, J. Effects of different thawing methods on the physicochemical changes, water migration and protein characteristic of frozen pompano (Trachinotus ovatus). J. Food Biochem. 2021, 45, e13826. [Google Scholar] [CrossRef]
- Ye, T.; Chen, Z.; Wu, Y.; Tao, J.; Wang, X.; Zhou, S.; Wang, Y.; Lu, J. Impact of high pressure processing on the shelling efficacy, myofibrillar protein, and cooking characteristics of fresh crayfish. Food Ferment. Ind. 2020, 46, 149–156. [Google Scholar]
- Zhang, Y.; Ertbjerg, P. On the origin of thaw loss: Relationship between freezing rate and protein denaturation. Food Chem. 2019, 299, 125104. [Google Scholar] [CrossRef]
- Qian, S.; Hu, F.; Mehmood, W.; Li, X.; Zhang, C.; Blecker, C. The rise of thawing drip: Freezing rate effects on ice crystallization and myowater dynamics changes. Food Chem. 2022, 373, 131461. [Google Scholar] [CrossRef]
- Du, X.; Li, H.; Nuerjiang, M.; Rui, L.; Kong, B.; Xia, X.; Shao, M. Influence of repeated freeze–thaw treatments on the functional and structural properties of myofibrillar protein from mirror carp (Cyprinus carpio L.). Food Biophys. 2021, 16, 492–501. [Google Scholar] [CrossRef]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Impact of freezing and thawing on the quality of meat: Review. Meat Sci. 2012, 91, 93–98. [Google Scholar] [CrossRef]
- Zhong, C.; Cai, Q.; Liu, G.; Sun, L.; Hara, K.; Su, W.; Cao, M. Purification and characterisation of cathepsin L from the skeletal muscle of blue scad (Decapterus maruadsi) and comparison of its role with myofibril-bound serine proteinase in the degradation of myofibrillar proteins. Food Chem. 2012, 133, 1560–1568. [Google Scholar] [CrossRef]
- Martínez-Maldonado, M.A.; Velazquez, G.; Ramírez de León, J.A.; Borderías, A.J.; Moreno, H.M. Effect of high pressure processing on heat-induced gelling capacity of blue crab (Callinectes sapidus) meat. Innov. Food Sci. Emerg. Technol. 2020, 59, 102253. [Google Scholar] [CrossRef]
- Cando, D.; Herranz, B.; Borderías, A.J.; Moreno, H.M. Effect of high pressure on reduced sodium chloride surimi gels. Food Hydrocoll. 2015, 51, 176–187. [Google Scholar] [CrossRef]
- Yang, S.; Hu, Y.; Takaki, K.; Yuan, C.; Yu, H. The impact of thawing on the quality attributes of swimming crab (Portunus trituberculatus) frozen by liquid nitrogen freezing. CYTA J. Food 2021, 19, 33–39. [Google Scholar] [CrossRef]
- Chen, X.; Tume, R.K.; Xiong, Y.; Xu, X.; Zhou, G.; Chen, C.; Nishiumi, T. Structural modification of myofibrillar proteins by high-pressure processing for functionally improved, value-added, and healthy muscle gelled foods. Crit. Rev. Food Sci. Nutr. 2018, 58, 2981–3003. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Zhou, P.; Zhang, X.; Wang, J. Effects of high pressure modification on conformation and gelation properties of myofibrillar protein. Food Chem. 2017, 217, 678–686. [Google Scholar] [CrossRef]
- Yu, W.; Wang, Z.; Pan, Y.; Jiang, P.; Pan, J.; Yu, C.; Dong, X. Effect of κ-carrageenan on quality improvement of 3D printed Hypophthalmichthys molitrix-sea cucumber compound surimi product. LWT—Food Sci. Technol. 2022, 154, 112279. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, K.; Tu, D.; Yu, X.; Dai, Z.; Shen, Q. Characterization of dietary fiber from wheat bran (Triticum aestivum L.) and its effect on the digestion of surimi protein. LWT—Food Sci. Technol. 2019, 102, 106–112. [Google Scholar] [CrossRef]
- Fang, M.; Xiong, S.; Hu, Y.; Yin, T.; You, J. In vitro pepsin digestion of silver carp (Hypophthalmichthys molitrix) surimi gels after cross-linking by Microbial Transglutaminase (MTGase). Food Hydrocoll. 2019, 95, 152–160. [Google Scholar] [CrossRef]
- Shao, Y.; Xiong, G.; Ling, J.; Hu, Y.; Shi, L.; Qiao, Y.; Yu, J.; Cui, Y.; Liao, L.; Wu, W.; et al. Effect of ultra-high pressure treatment on shucking and meat properties of red swamp crayfish (Procambarus clarkia). LWT—Food Sci. Technol. 2018, 87, 234–240. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, T.; Chen, X.; Zhu, Y.; Chen, Z.; Wang, Y.; Lin, L.; Zheng, Z.; Lu, J. Freeze-Thawing Treatment as a Simple Way to Tune the Gel Property and Digestibility of Minced Meat from Red Swamp Crayfish (Procambarus clarkiix). Foods 2022, 11, 837. https://doi.org/10.3390/foods11060837
Ye T, Chen X, Zhu Y, Chen Z, Wang Y, Lin L, Zheng Z, Lu J. Freeze-Thawing Treatment as a Simple Way to Tune the Gel Property and Digestibility of Minced Meat from Red Swamp Crayfish (Procambarus clarkiix). Foods. 2022; 11(6):837. https://doi.org/10.3390/foods11060837
Chicago/Turabian StyleYe, Tao, Xing Chen, Yajun Zhu, Zhina Chen, Yun Wang, Lin Lin, Zhi Zheng, and Jianfeng Lu. 2022. "Freeze-Thawing Treatment as a Simple Way to Tune the Gel Property and Digestibility of Minced Meat from Red Swamp Crayfish (Procambarus clarkiix)" Foods 11, no. 6: 837. https://doi.org/10.3390/foods11060837
APA StyleYe, T., Chen, X., Zhu, Y., Chen, Z., Wang, Y., Lin, L., Zheng, Z., & Lu, J. (2022). Freeze-Thawing Treatment as a Simple Way to Tune the Gel Property and Digestibility of Minced Meat from Red Swamp Crayfish (Procambarus clarkiix). Foods, 11(6), 837. https://doi.org/10.3390/foods11060837