
Wake Up! Resuscitation of Viable but Nonculturable Bacteria: Mechanism and Potential Application


Abstract
:1. Introduction
2. Confirmation of Resuscitation from the VBNC State
3. Resuscitation: The Reverse Process of the VBNC State?

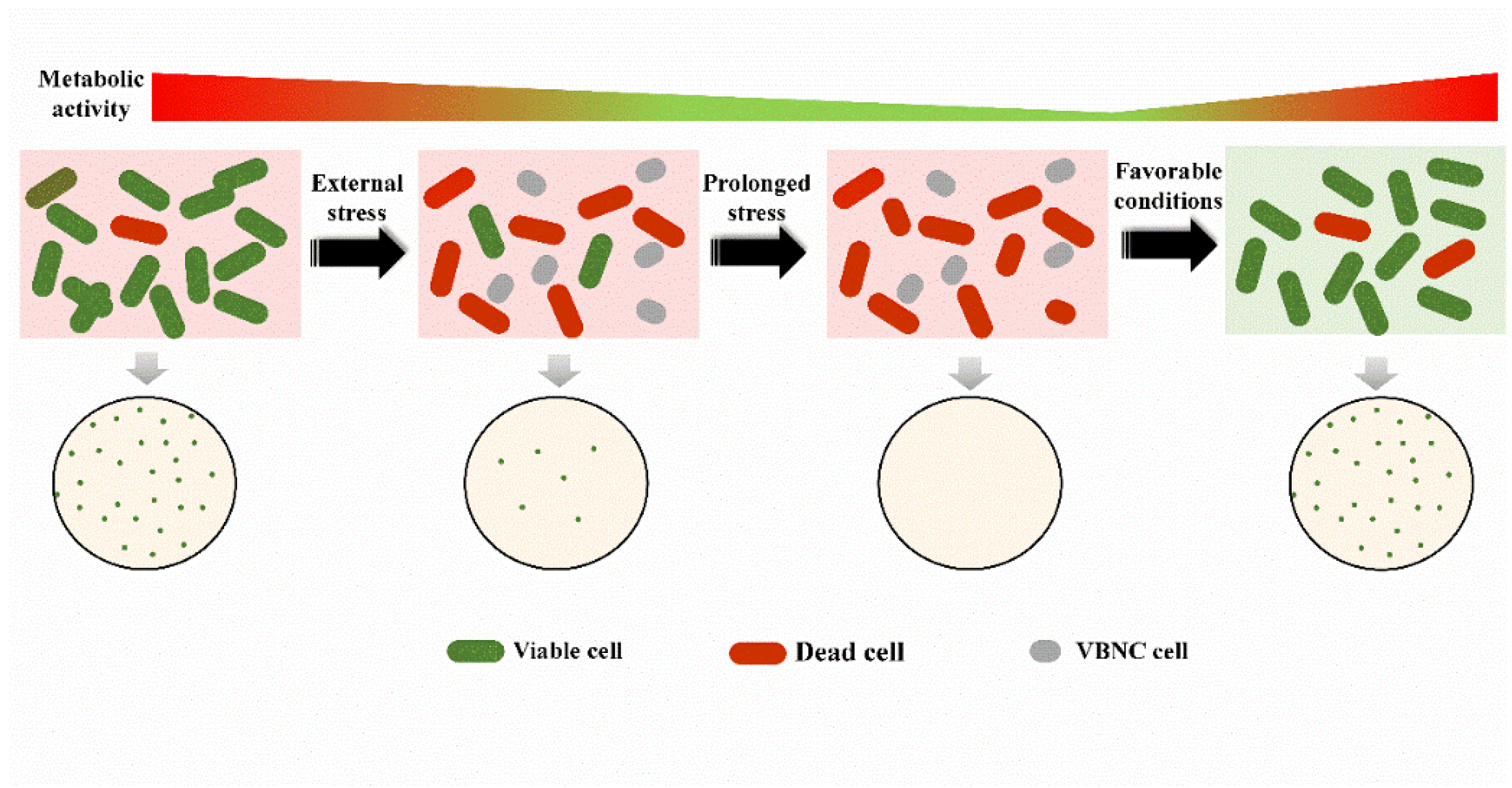{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resuscitation Factors | Bacterial Species | Resuscitation Conditions | References | |
|---|---|---|---|---|
| VBNC-State Induction Condition | Corresponding Resuscitation Condition | |||
| External stress removal | Arcobacter butzleri, Aeromonas hydrophila, Staphylococcus aureus, Vibrio vulnificus, E. coli | Low temperature | Temperature up-shift | [24,32,33,34,35,36] |
| Salmonella bovismorbificans, Enterococcus faecalis, Citrobacter sp., V. cholerae, Listeria monocytogenesisolates, Enterococci sp., Pasteurella piscicida, Yersinia pestis, V. shiloi, V. tasmaniensis, V. parahaemolyticus | Starvation | Addition of nutrients | [29,37,38,39,40,41,42,43,44,45] | |
| Enterobacter cloacae | Desiccation | Rewetting | [46] | |
| E. coli O157:H7, S. enterica serovar Typhimurium, L. monocytogenes | Low pH | Adjustment to the optimal pH | [47] | |
| Acetic acid bacteria, lactic acid bacteria | O2 deprivation | Addition of O2 | [48] | |
| E. coli O104:H4, Acidovorax citrulli, Erwinia amylovorain | Copper | Addition of chelating agent | [49,50,51] | |
| S. enterica, E. coli O157:H7 | Food processing techniques | Stress removal | [26,52] | |
| Supplementation with peroxidases | Yeasts, Ralstonia solanacearum, E. coli O157:H7, Enterococcus sp., Salmonella sp. S. aureus, V. cincinnatiensis | Catalase, sodium pyruvate, SOD, GST, CAT, acetaldehyde | [53,54,55,56,57,58,59,60,61] | |
| Host of VBNC cells | Legionella pneumophila, E. coli O157:H7, Campylobacter jejuni, Helicobacter pylori, L. monocytogenes, V. cholerae O1, Francisella tularensis, E. faecalis, Campylobacter sp. | Yolk sacs of embryonated eggs/1-week-old chicks, Caco-2 human intestinal epithelial cells, passage in the mouse intestine, co-culture with eukaryotic cells, injected intraperitoneally into mice, mice stomachs, co-culture with Acanthamoeba/Castellanii/Acanthamoeba polyphaga, ingestion by C. elegans, inoculated in iron-dextran-treated mice | [9,11,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79] | |
| Supplementation with substances that could promote resuscitation | Salmonella typhimurium, E. coli O157:H7, Vibrio sp., V. parahaemolyticus | Supplementation with autoinducer (AI) | [6,80,81,82] | |
| H. pylori, Mycobacterium tuberculosisare, Rhodococcus sp., actinobacteria, M. smegmatis, Sphingomonas and Pseudomonas, Rhodococcus biphenylivorans strain TG9T | Supplementation with resuscitation promoting factor (Rpf) | [83,84,85,86,87,88,89,90,91,92,93] | ||
4. Mechanisms of Resuscitation
4.1. Rpfs
4.2. Quorum Sensing
4.3. Pyruvates Sensing and Application
4.4. Mechanisms Based on Global Metabolism Analysis
5. Potential Application of the Resuscitation of VBNC Cells
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, K.; Pan, H.; Yang, D.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. Induction, detection, formation, and resuscitation of viable but non-culturable state microorganisms. Compr. Rev. Food Sci. Food Saf. 2020, 19, 149–183. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.D. The viable but nonculturable state in bacteria. J. Microbiol. 2005, 43, 93–100. [Google Scholar] [PubMed]
- Trevors, J.T. Can dead bacterial cells be defined and are genes expressed after cell death? J. Microbiol. Methods 2012, 90, 25–28. [Google Scholar] [CrossRef]
- Bogosian, G.; Bourneuf, E.V. A matter of bacterial life and death. EMBO Rep. 2001, 2, 770–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roszak, D.B.; Grimes, D.J.; Colwell, R.R. Viable but nonrecoverable stage of Salmonella enteritidis in aquatic systems. Can. J. Microbiol. 1984, 30, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Bae, Y.M.; Jo, S.; Moon, S.K.; Oh, S.W.; Lee, S.Y. Optimization of resuscitation-promoting broths for the revival of Vibrio parahaemolyticus from a viable but nonculturable state. Food Sci. Biotechnol. 2021, 30, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Ferro, S.; Amorico, T.; Deo, P. Role of food sanitising treatments in inducing the ‘viable but nonculturable’ state of microorganisms. Food Control 2018, 91, 321–329. [Google Scholar] [CrossRef]
- Makino, S.I.; Kii, T.; Asakura, H.; Shirahata, T.; Ikeda, T.; Takeshi, K.; Itoh, K. Does enterohemorrhagic Escherichia coli O157:H7 enter the viable but nonculturable state in salted salmon roe? Appl. Environ. Microbiol. 2000, 66, 5536–5539. [Google Scholar] [CrossRef] [Green Version]
- Alleron, L.; Khemiri, A.; Koubar, M.; Lacombe, C.; Coquet, L.; Cosette, P.; Jouenne, T.; Frere, J. VBNC Legionella pneumophila cells are still able to produce virulence proteins. Water Res. 2013, 47, 6606–6617. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, C.; Tyrrell, G.; Li, X.F. Production of Shiga-like toxins in viable but nonculturable Escherichia coli O157:H7. Water Res. 2010, 44, 711–718. [Google Scholar] [CrossRef]
- Chaisowwong, W.; Kusumoto, A.; Hashimoto, M.; Harada, T.; Maklon, K.; Kawamoto, K. Physiological characterization of Campylobacter jejuni under cold stresses conditions, its potential for public threat. J. Vet. Med. Sci. 2012, 74, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ahmad, W.; Zhu, X.; Chen, J.; Austion, B. Viable but nonculturable bacteria and their resuscitation: Implications for cultivating uncultured marine microorganisms. Mar. Life Sci. Technol. 2021, 3, 189–203. [Google Scholar] [CrossRef]
- Bogosian, G.; Aardema, N.D.; Bourneuf, E.V.; Morris, P.J.; O’Neil, J.P. Recovery of hydrogen peroxide-sensitive culturable cells of Vibrio vulnificus gives the appearance of resuscitation from a viable but nonculturable state. J. Bacteriol. 2000, 182, 5070–5075. [Google Scholar] [CrossRef] [Green Version]
- Kong, I.-S.; Bates, T.C.; Hulsmann, A.; Hassan, H.; Smith, B.E.; Oliver, J.D. Role of catalase and oxyR in the viable but nonculturable state of Vibrio vulnificus. FEMS Microbiol. Ecol. 2004, 50, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Knight, I.T.; Monahan, C.E.; Hill, R.T.; Colwell, R.R. Temperature-induced recovery of Vibrio cholerae from the viable but nonculturable state: Growth or resuscitation? Microbiology 1995, 141, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Ordax, M.; Marco-Noales, E.; Lopez, M.M.; Biosca, E.G. Survival strategy of Erwinia amylovora against copper: Induction of the viable-but-nonculturable state. Appl. Environ. Microbiol. 2006, 72, 3482–3488. [Google Scholar] [CrossRef] [Green Version]
- Whitesides, M.D.; Oliver, J.D. Resuscitation of Vibrio vulnificus from the viable but nonculturable state. Appl. Environ. Microbiol. 1997, 63, 1002–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puspita, I.D.; Kitagawa, W.; Kamagata, Y.; Tanaka, M.; Nakatsu, C.H. Increase in bacterial colony formation from a permafrost ice wedge dosed with a tomitella biformata recombinant resuscitation-promoting factor protein. Microbes Environ. 2015, 30, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basaglia, M.; Povolo, S.; Casella, S. Resuscitation of viable but not culturable Sinorhizobium meliloti 41 pRP4-luc: Effects of oxygen and host plant. Curr. Microbiol. 2007, 54, 167–174. [Google Scholar] [CrossRef]
- Boaretti, M.; Lleo, M.D.M.; Bonato, B.; Signoretto, C.; Canepari, P. Involvement of rpoS in the survival of Escherichia coli in the viable but non-culturable state. Environ. Microbiol. 2003, 5, 986–996. [Google Scholar] [CrossRef]
- Gupte, A.R.; de Rezende, C.L.E.; Joseph, S.W. Induction and resuscitation of viable but nonculturable Salmonella enterica Serovar Typhimurium DT104. Appl. Environ. Microbiol. 2003, 69, 6669–6675. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Wang, Y.; Zhao, L.; Rao, L.; Liao, X. Extracellular pH decline introduced by high pressure carbon dioxide is a main factor inducing bacteria to enter viable but non-culturable state. Food Res. Int. 2022, 151, 110895. [Google Scholar] [CrossRef] [PubMed]
- Weichart, D.; McDougald, D.; Jacobs, D.; Kjelleberg, S. In situ analysis of nucleic acids in cold-induced nonculturable Vibrio vulnificus. Appl. Environ. Microbiol. 1997, 63, 2754–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masmoudi, S.; Denis, M.; Maalej, S. Inactivation of the gene katA or sodA affects the transient entry into the viable but non-culturable response of Staphylococcus aureus in natural seawater at low temperature. Mar. Pollut. Bull. 2010, 60, 2209–2214. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Santos, M.A.; Chambel, L. Thirty years of viable but nonculturable state research: Unsolved molecular mechanisms. Crit. Rev. Microbiol. 2013, 41, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Bi, X.; Hao, Y.; Liao, X. Induction of viable but nonculturable Escherichia coli O157:H7 by high pressure CO2 and its characteristics. PLoS ONE 2013, 8, e62388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichart, D.; Kjelleberg, S. Stress resistance and recovery potential of culturable and viable but nonculturable cells of Vibrio vulnificus. Microbiology 1996, 142, 845–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signoretto, C.; Lleo, M.; Canepari, P. Modification of the peptidoglycan of Escherichia coli in the viable but nonculturable state. Curr. Microbiol. 2002, 44, 125–131. [Google Scholar] [CrossRef]
- Yoon, J.H.; Lee, S.Y. Characteristics of viable-but-nonculturable Vibrio parahaemolyticus induced by nutrient-deficiency at cold temperature. Crit. Rev. Food Sci. Nutr. 2019, 60, 1302–1320. [Google Scholar] [CrossRef]
- Nilsson, L.; Oliver, J.D.; Kjelleberg, S. Resuscitation of Vibrio vulnificus from the viable but nonculturable state. J. Bacteriol. 1991, 173, 5054–5059. [Google Scholar] [CrossRef]
- del Mar Lleò, M.; Benedetti, D.; Tafi, M.C.; Signoretto, C.; Canepari, P. Inhibition of the resuscitation from the viable but non-culturable state in Enterococcus faecalis. Environ. Microbiol. 2007, 9, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Fera, M.T.; Maugeri, T.L.; Gugliandolo, C.; la Camera, E.; Lentini, V.; Favaloro, A.; Bonanno, D.; Carbone, M. Induction and resuscitation of viable nonculturable Arcobacter butzleri cells. Appl. Environ. Microbiol. 2008, 74, 3266–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maalej, S.; Gdoura, R.; Dukan, S.; Hammami, A.; Bouain, A. Maintenance of pathogenicity during entry into and resuscitation from viable but nonculturable state in Aeromonas hydrophila exposed to natural seawater at low temperature. J. Appl. Microbiol. 2004, 97, 557–565. [Google Scholar] [CrossRef]
- Nowakowska, J.; Oliver, J.D. Resistance to environmental stresses by Vibrio vulnificus in the viable but nonculturable state. FEMS Microbiol. Ecol. 2013, 84, 213–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, N.V.; Shashidhar, R.; Bandekar, J.R. Induction, resuscitation and quantitative real-time polymerase chain reaction analyses of viable but nonculturable Vibrio vulnificus in artificial sea water. World J. Microbiol. Biotechnol. 2014, 30, 2205–2212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ye, C.; Lin, H.; Lv, L.; Yu, X. UV disinfection induces a VBNC state in Escherichia coli and Pseudomonas aeruginosa. Environ. Sci. Technol. 2015, 49, 1721–1728. [Google Scholar] [CrossRef]
- Ben Abdallah, F.; Lagha, R.; Bakhrouf, A. Resuscitation and morphological alterations of Salmonella bovismorbificans cells under starvation in soil. World J. Microbiol. Biotechnol. 2007, 24, 1507–1512. [Google Scholar] [CrossRef]
- del Mar Lleo, M.; Tafi, M.C.; Canepari, P. Nonculturable Enterococcus faecalis cells are metabolically active and capable of resuming active growth. Syst. Appl. Microbiol. 1998, 21, 333–339. [Google Scholar] [CrossRef]
- Dhiaf, A.; Bakhrouf, A.; Witzel, K.P. Resuscitation of eleven-year VBNC Citrobacter. J. Water Health 2008, 6, 565. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Delgado, M.; García-Amado, M.A.; Contreras, M.; Incani, R.N.; Chirinos, H.; Rojas, H.; Suárez, P. Survival, induction and resuscitation of Vibrio cholerae from the viable but non-culturable state in the Southern Caribbean Sea. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 21–26. [Google Scholar] [CrossRef]
- Lindbäck, T.; Rottenberg, M.E.; Roche, S.M.; Rørvik, L.M. The ability to enter into an avirulent viable but non-culturable (VBNC) form is widespread among Listeria monocytogenesisolates from salmon, patients and environment. Vet. Res. 2009, 41, 08. [Google Scholar] [CrossRef] [Green Version]
- Lleò, M.; Bonato, B.; Signoretto, C.; Canepari, P. Vancomycin resistance is maintained in enterococci in the viable but nonculturable state and after division is resumed. Antimicrob. Agents Chemother. 2003, 47, 1154–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magarinos, B.; Romalde, J.; Cid, A.; Toranzo, A. Viability of starved Pasteurella piscicida in seawater monitored by flow cytometry and the effect of antibiotics on its resuscitation. Lett. Appl. Microbiol. 1997, 24, 122–126. [Google Scholar] [CrossRef]
- Pawlowski, D.R.; Metzger, D.J.; Raslawsky, A.; Howlett, A.; Siebert, G.; Karalus, R.J.; Garrett, S.; Whitehouse, A.C. Entry of Yersinia pestis into the viable but nonculturable state in a low-temperature tap water microcosm. PLoS ONE 2011, 6, e17585. [Google Scholar] [CrossRef] [PubMed]
- Vattakaven, T.; Bond, P.; Bradley, G.; Munn, C.B. Differential effects of temperature and starvation on induction of the viable-but-nonculturable state in the coral pathogens Vibrio shiloi and Vibrio tasmaniensis. Appl. Environ. Microbiol. 2006, 72, 6508–6513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.C.; Jacobsen, C.S. Fate of Enterobacter cloacae JP120 and Alcaligenes eutrophus AEO106(pRO101) in soil during water stress: Effects on culturability and viability. Appl. Environ. Microbiol. 1993, 59, 1560–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolò, M.S.; Gioffrè, A.; Carnazza, S.; Platania, G.; Silvestro, I.D.; Guglielmino, S.P.P. Viable but nonculturable state of foodborne pathogens in grapefruit juice: A study of laboratory. Foodborne Pathog. Dis. 2011, 8, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Millet, V.; Lonvaud-Funel, A. The viable but non-culturable state of wine micro-organisms during storage. Lett. Appl. Microbiol. 2000, 30, 136–141. [Google Scholar] [CrossRef]
- Aurass, P.; Prager, R.; Flieger, A. EHEC/EAEC O104:H4 strain linked with the 2011 German outbreak of haemolytic uremic syndrome enters into the viable but non-culturable state in response to various stresses and resuscitates upon stress relief. Environ. Microbiol. 2011, 13, 3139–3148. [Google Scholar] [CrossRef]
- Kan, Y.; Jiang, N.; Xu, X.; Lyu, Q.; Gopalakrishnan, V.; Walcott, R.; Burdman, S.; Li, J.; Luo, L. 2019. Induction and resuscitation of the viable but non-culturable (VBNC) state in Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbitaceous crops. Front. Microbiol. 2019, 10, 1081. [Google Scholar] [CrossRef]
- Ordax, M.; Biosca, E.G.; Wimalajeewa, S.C.; López, M.M.; Marco-Noales, E. Survival of Erwinia amylovorain mature apple fruit calyces through the viable but nonculturable (VBNC) state. J. Appl. Microbiol. 2009, 107, 106–116. [Google Scholar] [CrossRef]
- Purevdorj-Gage, L.; Nixon, B.; Bodine, K.; Xu, Q.; Doerrler, W.T. Differential effect of food sanitizers on formation of viable but nonculturable Salmonella enterica in poultry. J. Food Prot. 2018, 81, 386–393. [Google Scholar] [CrossRef]
- Divol, B.; Lonvaud-Funel, A. Evidence for viable but nonculturable yeasts in botrytis-affected wine. J. Appl. Microbiol. 2005, 99, 85–93. [Google Scholar] [CrossRef]
- Imazaki, I.; Nakaho, K. Temperature-upshift-mediated revival from the sodium-pyruvate-recoverable viable but nonculturable state induced by low temperature in Ralstonia solanacearum: Linear regression analysis. J. Gen. Plant Pathol. 2009, 75, 213–226. [Google Scholar] [CrossRef]
- Kolling, G.L.; Matthews, K.R. Examination of recovery in vitro and in vivo of nonculturable Escherichia coli O157: H7. Appl. Environ. Microbiol. 2001, 67, 3928–3933. [Google Scholar] [CrossRef] [Green Version]
- Lleo, M.M.; Bonato, B.; Tafi, M.C.; Signoretto, C.; Boaretti, M.; Canepari, P. Resuscitation rate in different enterococcal species in the viable but non-culturable state. J. Appl. Microbiol. 2001, 91, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Morishige, Y.; Fujimori, K.; Amano, F. Differential resuscitative effect of pyruvate and its analogues on VBNC (viable but non-culturable) Salmonella. Microbes Environ. 2013, 28, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishige, Y.; Koike, A.; Tamura-Ueyama, A.; Amano, F. Induction of viable but nonculturable salmonella in exponentially grown cells by exposure to a low-humidity environment and their resuscitation by catalase. J. Food Prot. 2017, 80, 288–294. [Google Scholar] [CrossRef]
- Pasquaroli, S.; Zandri, G.; Vignaroli, C.; Vuotto, C.; Donelli, G.; Biavasco, F. Antibiotic pressure can induce the viable but non-culturable state in Staphylococcus aureus growing in biofilms. J. Antimicrob. Chemother. 2013, 68, 1812–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.H.; Kong, H.G.; Bae, J.Y.; Lee, H.J.; Joo, H.J.; Jung, E.J.; Chung, E.; Lee, S.W. Induction of the viable but nonculturable state of Ralstonia solanacearum by low temperature in the soil microcosm and its resuscitation by Catalase. PLoS ONE 2014, 9, e109792. [Google Scholar]
- Zhong, L.; Chen, J.; Zhang, X.; Jiang, Y. Entry of Vibrio cincinnatiensis into viable but nonculturable state and its resuscitation. Lett. Appl. Microbiol. 2009, 48, 247–252. [Google Scholar] [CrossRef]
- Alleron, L.; Merlet, N.; Lacombe, C.; Frère, J. Long-term survival of Legionella pneumophila in the viable but nonculturable state after monochloramine treatment. Curr. Microbiol. 2008, 57, 497–502. [Google Scholar] [CrossRef]
- Asakura, H.; Igimi, S.; Kawamoto, K.; Yamamoto, S.; Makino, S. Role of in vivo passage on the environmental adaptation of enterohemorrhagic Escherichia coli O157:H7: Cross-induction of the viable but nonculturable state by osmotic and oxidative stresses. FEMS Microbiol. Lett. 2005, 253, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Baffone, W.; Casaroli, A.; Citterio, B.; Pierfelici, L.; Campana, R.; Vittoria, E.; Guaglianone, E.; Donelli, G. Campylobacter jejuni loss of culturability in aqueous microcosms and ability to resuscitate in a mouse model. Int. J. Food Microbiol. 2006, 107, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, K.F.; Eaton, K.A.; Fontaine, C.; Brewster, R.; Wu, J.; Eisenberg, J.N.S.; Valdivieso, M.; Baker, L.H.; Xi, C. Reduced infectivity of waterborne viable but nonculturable Helicobacter pylori strain SS1 in mice. Helicobacter 2017, 22, e12391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappelier, J.; Minet, J.; Magras, C.; Colwell, R.; Federighi, M. Recovery in embryonated eggs of viable but nonculturable Campylobacter jejuni cells and maintenance of ability to adhere to HeLa cells after resuscitation. Appl. Environ. Microbiol. 1999, 65, 5154–5157. [Google Scholar] [CrossRef] [Green Version]
- Cappelier, J.M.; Besnard, V.; Roche, S.; Garrec, N.; Zundel, E.; Velge, P.; Federighi, M. Avirulence of viable but non-culturable Listeria monocytogenes cells demonstrated by in vitro and in vivo models. Vet. Res. 2005, 36, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappelier, J.M.; Besnard, V.; Roche, S.M.; Velge, P.; Federigh, M. Avirulent viable but non culturable cells of Listeria monocytogenes need the presence of an embryo to be recovered in egg yolk and regain virulence after recovery. Vet. Res. 2007, 38, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.; Brayton, P.; Herrington, D.; Tall, B.; Huq, A.; Levine, M. Viable but non-culturable Vibrio cholerae O1 revert to a cultivable state in the human intestine. World J. Microbiol. Biotechnol. 1996, 12, 28–31. [Google Scholar] [CrossRef]
- Epalle, T.; Girardot, F.; Allegra, S.; Maurice-Blanc, C.; Garraud, O.; Riffard, S. Viable but not culturable forms of Legionella pneumophila generated after heat shock treatment are infectious for macrophage-like and alveolar epithelial cells after resuscitation on Acanthamoeba polyphaga. Microb. Ecol. 2014, 69, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Forsman, M.; Henningson, E.W.; Larsson, E.; Johansson, T.; Sandström, G. Francisella tularensis does not manifest virulence in viable but non-culturable state. FEMS Microbiol. Ecol. 2000, 31, 217–224. [Google Scholar] [CrossRef] [PubMed]
- García, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Kwaik, Y.A. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277. [Google Scholar] [CrossRef]
- Highmore, C.J.; Warner, J.C.; Rothwell, S.D.; Wilks, S.A.; Keevil, C.W. Viable-but-nonculturable Listeria monocytogenes and Salmonella enterica serovar Thompson induced by chlorine stress remain infectious. mBio 2018, 9, e00540-e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klančnik, A.; Guzej, B.; Jamnik, P.; Vučković, D.; Abram, M.; Možina, S.S. Stress response and pathogenic potential of Campylobacter jejuni cells exposed to starvation. Res. Microbiol. 2009, 160, 345–352. [Google Scholar] [CrossRef]
- Mustapha, P.; Epalle, T.; Allegra, S.; Girardot, F.; Garraud, O.; Riffard, S. Monitoring of Legionella pneumophila viability after chlorine dioxide treatment using flow cytometry. Res. Microbiol. 2015, 166, 215–219. [Google Scholar] [CrossRef]
- Pruzzo, C.; Tarsi, R.; Lleò, M.D.M.; Signoretto, C.; Zampini, M.; Colwell, R.R.; Canepari, P. In vitro adhesion to human cells by viable but nonculturable Enterococcus faecalis. Curr. Microbiol. 2002, 45, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Senoh, M.; Ghosh-Banerjee, J.; Ramamurthy, T.; Colwell, R.R.; Miyoshi, S.; Nair, G.B.; Takeda, Y. Conversion of viable but nonculturable enteric bacteria to culturable by co-culture with eukaryotic cells. Microbiol. Immunol. 2012, 56, 342–345. [Google Scholar] [CrossRef]
- Senoh, M.; Ghosh-Banerjee, J.; Ramamurthy, T.; Hamabata, T.; Kurakawa, T.; Takeda, M.; Colwell, R.R.; Nair, G.B.; Takeda, Y. Conversion of viable but nonculturable Vibrio cholerae to the culturable state by co-culture with eukaryotic cells. Microbiol. Immunol. 2010, 54, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Stern, N.; Jones, D.; Wesley, I.; Rollins, D. Colonization of chicks by non-culturable Campylobacter spp. Lett. Appl. Microbiol. 1994, 18, 333–336. [Google Scholar] [CrossRef]
- Liao, H.; Zhong, X.; Xu, L.; Ma, Q.; Wang, Y.; Cai, Y.; Guo, X. Quorum-sensing systems trigger catalase expression to reverse the oxyR deletion-mediated VBNC state in Salmonella typhimurium. Res. Microbiol. 2019, 170, 65–73. [Google Scholar] [CrossRef]
- Liu, Y.; Kumblathan, T.; Uppal, G.K.; Zhou, A.; Moe, B.; Hrudey, S.E.; Li, X.F. A hidden risk: Survival and resuscitation of Escherichia coli O157:H7 in the viable but nonculturable state after boiling or microwaving. Water Res. 2020, 183, 116102. [Google Scholar] [CrossRef] [PubMed]
- Ayrapetyan, M.; Williams, T.C.; Oliver, J.D. Interspecific quorum sensing mediates the resuscitation of viable but nonculturable vibrios. Appl. Environ. Microbiol. 2014, 80, 2478–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, D.; Bagirova, M.; Allahverdiyev, A.M.; Abamor, E.S.; Safarov, T.; Kocazeybek, B.S. Resuscitation of the Helicobacter pylori coccoid forms by resuscitation promoter factor obtained from Micrococcus luteus. Curr. Microbiol. 2020, 77, 2093–2103. [Google Scholar] [CrossRef] [PubMed]
- Kana, B.D.; Gordhan, B.G.; Downing, K.J.; Sung, N.; Vostroktunova, G.; Machowski, E.E.; Tsenova, L.; Young, M.; Kaprelyants, A.; Kaplan, G.; et al. The resuscitation-promoting factors of Mycobacterium tuberculosisare required for virulence and resuscitation from dormancy but are collectively dispensable for growth in vitro. Mol. Microbiol. 2008, 67, 672–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Su, X.; Guo, L.; Ding, L.; Qu, K.; Shen, C. Induction of viable but nonculturable state in Rhodococcus and transcriptome analysis using RNA-seq. PLoS ONE 2016, 11, e0147593. [Google Scholar]
- Nikitushkin, V.D.; Demina, G.R.; Kaprelyants, A.S. Rpf proteins are the factors of reactivation of the dormant forms of actinobacteria. Biochem. Biokhimiia 2016, 81, 1719–1734. [Google Scholar] [CrossRef]
- Shleeva, M.; Mukamolova, G.V.; Young, M.; Williams, H.D.; Kaprelyants, A.S. Formation of ‘non-culturable’cells of Mycobacterium smegmatis in stationary phase in response to growth under suboptimal conditions and their Rpf-mediated resuscitation. Microbiology 2004, 150, 1687–1697. [Google Scholar] [CrossRef]
- Shleeva, M.O.; Kudykina, Y.K.; Vostroknutova, G.N.; Suzina, N.E.; Mulyukin, A.L.; Kaprelyants, A.S. Dormant ovoid cells of Mycobacterium tuberculosis are formed in response to gradual external acidification. Tuberculosis 2011, 91, 146–154. [Google Scholar] [CrossRef]
- Su, X.; Li, S.; Xie, M.; Tao, L.; Zhou, Y.; Xiao, Y.; Lin, H.; Chen, J.; Sun, F. Enhancement of polychlorinated biphenyl biodegradation by resuscitation promoting factor (Rpf) and Rpf-responsive bacterial community. Chemosphere 2021, 263, 128283. [Google Scholar] [CrossRef]
- Su, X.; Sun, F.; Wang, Y.; Hashmi, M.Z.; Guo, L.; Ding, L.; Shen, C. Identification, characterization and molecular analysis of the viable but nonculturable Rhodococcus biphenylivorans. Sci. Rep. 2015, 5, 18590. [Google Scholar] [CrossRef]
- Su, X.; Wang, Y.; Xue, B.; Zhang, Y.; Mei, R.; Zhang, Y.; Hashmi, M.Z.; Lin, H.; Chen, J.; Sun, F. Resuscitation of functional bacterial community for enhancing biodegradation of phenol under high salinity conditions based on Rpf. Bioresour. Technol. 2018, 261, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Xue, B.; Wang, Y.; Hashmi, M.Z.; Lin, H.; Chen, J.; Mei, R.; Wang, Z.; Sun, F. Bacterial community shifts evaluation in the sediments of Puyang River and its nitrogen removal capabilities exploration by resuscitation promoting factor. Ecotoxicol. Environ. Saf. 2019, 179, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Li, H.; Jia, Y.; Fan, J.; Wan, J.; Guo, L.; Su, X.; Zhang, Y.; Wu, W.M.; Shen, C. Supplementing resuscitation-promoting factor (Rpf) enhanced biodegradation of polychlorinated biphenyls (PCBs) by Rhodococcus biphenylivorans strain TG9T. Environ. Pollut. 2020, 263, 114488. [Google Scholar] [CrossRef] [PubMed]
- Giagnoni, L.; Arenella, M.; Galardi, E.; Nannipieri, P.; Renella, G. Bacterial culturability and the viable but non-culturable (VBNC) state studied by a proteomic approach using an artificial soil. Soil Biol. Biochem. 2018, 118, 51–58. [Google Scholar] [CrossRef]
- Pan, H.; Dong, K.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. The association of cell division regulated by DicC with the formation of viable but non-culturable Escherichia coli O157:H7. Front. Microbiol. 2019, 10, 2850. [Google Scholar] [CrossRef] [PubMed]
- Santander, R.D.; Figas-Segura, A.; Biosca, E.G. Erwinia amylovora catalases KatA and KatG are virulence factors and delay the starvation-induced viable but nonculturable (VBNC) response. Mol. Plant Pathol. 2017, 19, 922–934. [Google Scholar] [CrossRef] [Green Version]
- Mukamolova, G.V.; Kaprelyants, A.S.; Kell, D.B. Secretion of an antibacterial factor during resuscitation of dormant cells in Micrococcus luteus cultures held in an extended stationary phase. Antonie Leeuwenhoek 1995, 67, 289–295. [Google Scholar] [CrossRef]
- Mukamolova, G.V.; Kaprelyants, A.S.; Young, D.I.; Young, M.; Kell, D.B. A bacterial cytokine. Proc. Natl. Acad. Sci. USA 1998, 95, 8916–8921. [Google Scholar] [CrossRef] [Green Version]
- Mukamolova, G.V.; Murzin, A.G.; Salina, E.G.; Demina, G.R.; Kell, D.B.; Kaprelyants, A.S.; Young, M. Muralytic activity of Micrococcus luteus Rpf and its relationship to physiological activity in promoting bacterial growth and resuscitation. Mol. Microbiol. 2006, 59, 84–98. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Chen, X.; Hu, J.; Shen, C.; Ding, L. Exploring the potential environmental functions of viable but non-culturable bacteria. World J. Microbiol. Biotechnol. 2013, 29, 2213–2218. [Google Scholar] [CrossRef]
- Nikitushkin, V.D.; Demina, G.R.; Shleeva, M.O.; Guryanova, S.V.; Ruggiero, A.; Berisio, R.; Kaprelyants, A.S. A Product of RpfB and RipA joint enzymatic action promotes the resuscitation of dormant mycobacteria. FEBS J. 2015, 282, 2500–2511. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, D.J.; Pavelka, M.S., Jr. The RipA and RipB Peptidoglycan Endopeptidases Are Individually Nonessential to Mycobacterium smegmatis. J. Bacteriol. 2016, 198, 1464–1475. [Google Scholar] [CrossRef] [PubMed]
- Hett, E.C.; Chao, M.C.; Deng, L.L.; Rubin, E.J. A mycobacterial enzyme essential for cell division synergizes with resuscitation-promoting factor. PLoS Pathog. 2008, 4, e1000001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hett, E.C.; Chao, M.C.; Rubin, E.J. Interaction and modulation of two antagonistic cell wall enzymes of mycobacteria. PLoS Pathog. 2010, 6, e1001020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signoretto, C.; Lleo, M.M.; Tafi, M.C.; Canepari, P. Cell wall chemical composition of Enterococcus faecalis in the viable but nonculturable state. Appl. Environ. Microbiol. 2000, 66, 1953–1959. [Google Scholar] [CrossRef] [Green Version]
- Nikitushkin, V.D.; Demina, G.R.; Shleeva, M.O.; Kaprelyants, A.S. Peptidoglycan fragments stimulate resuscitation of “non-culturable” mycobacteria. Antonie Leeuwenhoek 2013, 103, 37–46. [Google Scholar] [CrossRef]
- Panutdaporn, N.; Kawamoto, K.; Asakura, H.; Makino, S.-I. Resuscitation of the viable but non-culturable state of Salmonella enterica serovar Oranienburg by recombinant resuscitation-promoting factor derived from Salmonella Typhimurium strain LT2. Int. J. Food Microbiol. 2006, 106, 241–247. [Google Scholar] [CrossRef]
- Mir, M.; Asong, J.; Li, X.; Cardot, J.; Boons, G.J.; Husson, R.N. The extracytoplasmic domain of the Mycobacterium tuberculosis Ser/Thr kinase PknB binds specific muropeptides and is required for PknB localization. PLoS Pathog. 2011, 7, e1002182. [Google Scholar] [CrossRef] [Green Version]
- Aydin, I.; Saijo-Hamano, Y.; Namba, K.; Thomas, C.; Roujeinikova, A. Structural analysis of the essential resuscitation promoting factor YeaZ suggests a mechanism of nucleotide regulation through dimer reorganization. PLoS ONE 2011, 6, e23245. [Google Scholar] [CrossRef] [Green Version]
- Handford, J.I.; Ize, B.; Buchanan, G.; Butland, G.P.; Greenblatt, J.; Emili, A.; Palmer, T. Conserved network of proteins essential for bacterial viability. J. Bacteriol. 2009, 191, 4732–4749. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, J.; Zhao, M.; Yang, Z.; Yue, L.; Zhang, X. Promoting resuscitation of viable but nonculturable cells of Vibrio harveyi by a resuscitation-promoting factor-like protein YeaZ. J. Appl. Microbiol. 2017, 122, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Vecchietti, D.; Ferrara, S.; Rusmini, R.; Macchi, R.; Milani, M.; Bertoni, G. Crystal structure of YeaZ from Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2016, 470, 460–465. [Google Scholar] [CrossRef]
- Zhao, R.; Chen, J.; Wang, Y.; Li, Y.; Kong, X.; Han, Y. Proteolytic activity of Vibrio harveyi YeaZ is related with resuscitation on the viable but non-culturable state. Lett. Appl. Microbiol. 2020, 71, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial Quorum Sensing and Microbial Community Interactions. mBio 2018, 9, e02331-e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassett, D.J.; Ma, J.F.; Elkins, J.G.; McDermott, T.R.; Ochsner, U.A.; West, S.E.; Huang, C.T.; Fredericks, J.; Burnett, S.; Stewart, P.; et al. Quorum sensing in Pseudomonas aeruginosa controls expression of catalase and superoxide dismutase genes and mediates biofilm susceptibility to hydrogen peroxide. Mol. Microbiol. 1999, 34, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Kang, M.; Kim, S.H.; Lee, H.; Lim, J.; Choi, S.H.; Park, S.; Lee, K. Isolation and characterization of rpoS from a pathogenic bacterium, Vibrio vulnificus: Role of σs in survival of exponential-phase cells under oxidative stress. J. Bacteriol. 2004, 186, 3304–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, X. Effects of quorum sensing on the biofilm formation and viable but non-culturable state. Food Res. Int. 2020, 137, 109742. [Google Scholar] [CrossRef]
- McDonald, L.C.; Hackney, C.R.; Ray, B. Enhanced recovery of injured Escherichia coli by compounds that degrade hydrogen peroxide or block its formation. Appl. Environ. Microbiol. 1983, 45, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Mizunoe, Y.; Wai, S.N.; Ishikawa, T.; Takade, A.; Yoshida, S. Resuscitation of viable but nonculturable cells of Vibrio parahaemolyticus induced at low temperature under starvation. FEMS Microbiol. Lett. 2000, 186, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Mizunoe, Y.; Wai, S.N.; Takade, A.; Yoshida, S. Restoration of culturability of starvation-stressed and low-temperature-stressed Escherichia coli O157 cells by using H2O2-degrading compounds. Arch. Microbiol. 1999, 172, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Vilhena, C.; Kaganovitch, E.; Grünberger, A.; Motz, M.; Forné, I.; Kohlheyer, D.; Jung, K. Importance of pyruvate sensing and transport for the resuscitation of viable but nonculturable Escherichia coli K-12. J. Bacteriol. 2019, 201, e00610–e00618. [Google Scholar] [CrossRef] [PubMed]
- Behr, S.; Fried, L.; Jung, K. Identification of a novel nutrient-sensing histidine kinase/response regulator network in Escherichia coli. J. Bacteriol. 2014, 196, 2023–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behr, S.; Kristoficova, I.; Witting, M.; Breland, E.J.; Eberly, A.R.; Sachs, C.; Schmitt-Kopplin, P.; Hadjifrangiskou, M. Identification of a high-affinity pyruvate receptor in Escherichia coli. Sci. Rep. 2017, 7, 1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, Y.; Lyu, Q.; Jiang, N.; Han, S.; Li, J.; Burdman, S.; Luo, L. iTRAQ-based proteomic analyses of the plant-pathogenic bacterium Acidovorax citrulli during entrance into and resuscitation from the viable but nonculturable state. J. Proteom. 2020, 211, 103547. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Wang, B.; Wang, J.; Liu, Y.; Fang, X.; Liao, Z. Global proteomic analysis of the resuscitation state of Vibrio parahaemolyticus compared with the normal and viable but non-culturable state. Front. Microbiol. 2019, 10, 1045. [Google Scholar] [CrossRef]
- Debnath, A.; Mizuno, T.; Miyoshi, S.I. Comparative proteomic analysis to characterize temperature-induced viable but non-culturable and resuscitation states in Vibrio cholerae. Microbiology 2019, 165, 737–746. [Google Scholar] [CrossRef]
- Lopez Marin, M.A.; Strejcek, M.; Junkova, P.; Suman, J.; Santrucek, J.; Uhlik, O. Exploring the potential of Micrococcus luteus culture supernatant with resuscitation-promoting factor for enhancing the culturability of soil bacteria. Front. Microbiol. 2021, 12, 685263. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Tang, L.; Zhang, Y.; Zhang, Y.; Wang, X.; Zhang, X. Evaluation of Rpf protein of Micrococcus luteus for cultivation of soil actinobacteria. Syst. Appl. Microbiol. 2021, 44, 126234. [Google Scholar] [CrossRef]
- Su, X.; Zhang, S.; Mei, R.; Zhang, Y.; Hashmi, M.Z.; Liu, J.; Lin, H.; Ding, L.; Sun, F. Resuscitation of viable but non-culturable bacteria to enhance the cellulose-degrading capability of bacterial community in composting. Microb. Biotechnol. 2018, 11, 527–536. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, H.; Ren, Q. Wake Up! Resuscitation of Viable but Nonculturable Bacteria: Mechanism and Potential Application. Foods 2023, 12, 82. https://doi.org/10.3390/foods12010082
Pan H, Ren Q. Wake Up! Resuscitation of Viable but Nonculturable Bacteria: Mechanism and Potential Application. Foods. 2023; 12(1):82. https://doi.org/10.3390/foods12010082
Chicago/Turabian StylePan, Hanxu, and Qing Ren. 2023. "Wake Up! Resuscitation of Viable but Nonculturable Bacteria: Mechanism and Potential Application" Foods 12, no. 1: 82. https://doi.org/10.3390/foods12010082
APA StylePan, H., & Ren, Q. (2023). Wake Up! Resuscitation of Viable but Nonculturable Bacteria: Mechanism and Potential Application. Foods, 12(1), 82. https://doi.org/10.3390/foods12010082