
Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry

Abstract
:1. Introduction
2. Materials and Methods
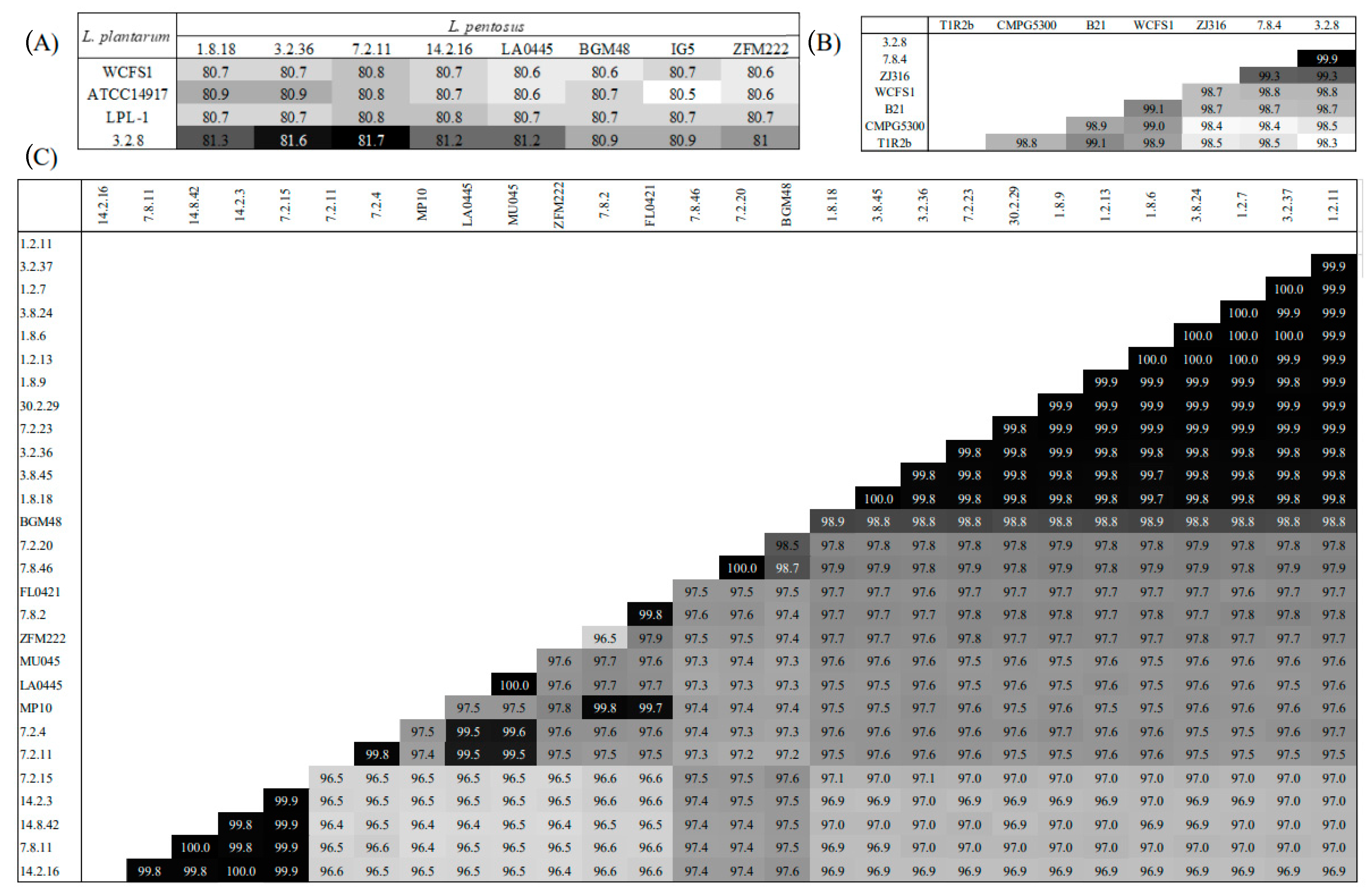
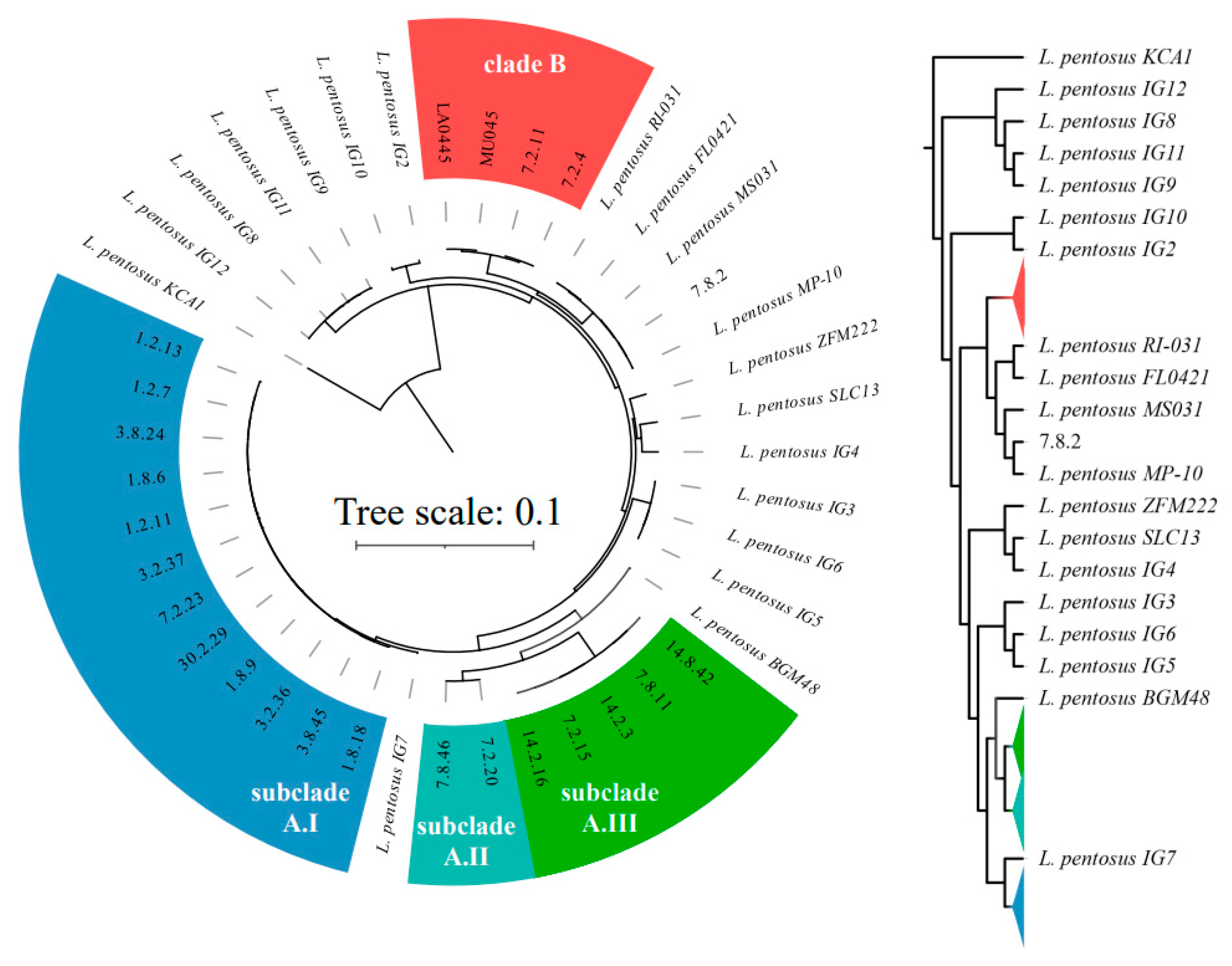
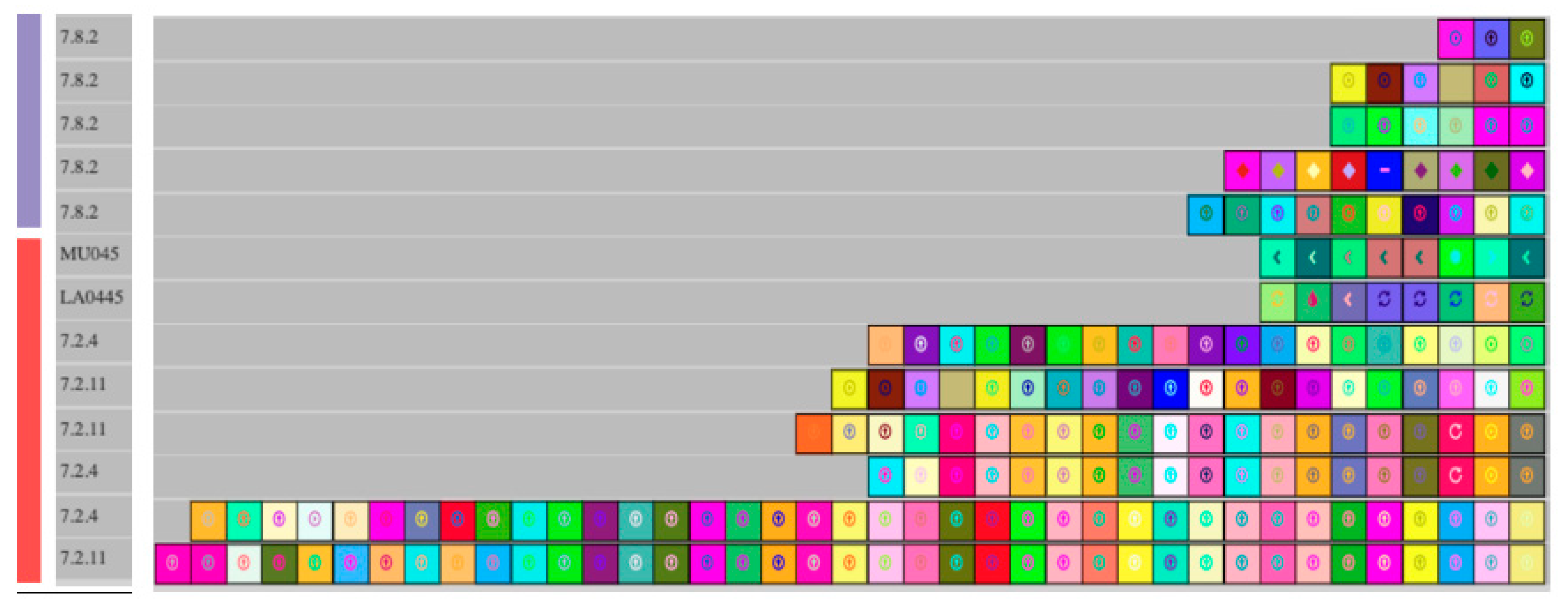
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Sahlany, S.T.; Niamah, A.K. Bacterial viability, antioxidant stability, antimutagenicity and sensory properties of onion types fermentation by using probiotic starter during storage. Nutr. Food Sci. 2022, 52, 901–916. [Google Scholar] [CrossRef]
- Benítez-Cabello, A.; Bautista-Gallego, J.; Garrido-Fernández, A.; Rantsiou, K.; Cocolin, L.; Jiménez-Díaz, R.; Arroyo-López, F.N. RT-PCR–DGGE Analysis to Elucidate the Dominant Bacterial Species of Industrial Spanish-Style Green Table Olive Fermentations. Front. Microbiol. 2016, 7, 1291. [Google Scholar] [CrossRef]
- Chen, K.-H.; McFEETERS, R.F.; Fleming, H.P. Stability of Mannitol to Lactobacillus plantarum Degradation in Green Beans Fermented with Lactobacillus cellobiosus. J. Food Sci. 1983, 48, 972–974. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Siragusa, S.; De Angelis, M.; Paradiso, A.; Minervini, F.; De Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic Analysis of Kimchi, a Traditional Korean Fermented Food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef]
- Kotzekidou, P.; Roukas, T. Characterization and Distribution of Lactobacilli during Lactic Fermentation of Okra (Hibiscus esculentus). J. Food Sci. 1986, 51, 623–625. [Google Scholar] [CrossRef]
- Lee, Y.G.; Cho, J.-Y.; Kim, Y.-M.; Moon, J.-H. Change in Flavonoid Composition and Antioxidative Activity during Fermentation of Onion (Allium cepa L.) by Leuconostoc mesenteroides with Different Salt Concentrations. J. Food Sci. 2016, 81, C1385–C1393. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Kouretas, K.; Drosinos, E.H. Effect of ripening stage on the development of the microbial community during spontaneous fermentation of green tomatoes. J. Sci. Food Agric. 2014, 94, 1600–1606. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Dickey, A.; Fitria, R.; Ravishankar, N.; Hayes, J.S.; Campbell, K.; Arritt, F. Modulation of the bacterial population in commercial cucumber fermentations by brining salt type. J. Appl. Microbiol. 2020, 128, 1678–1693. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Hayes, J.; Medina, E.; Anekella, K.; Daughtry, K.; Dieck, S.; Levi, M.; Price, R.; Butz, N.; Lu, Z.; et al. Reassessment of the succession of lactic acid bacteria in commercial cucumber fermentations and physiological and genomic features associated with their dominance. Food Microbiol. 2017, 63, 217–227. [Google Scholar] [CrossRef]
- Plengvidhya, V.; Breidt, F.; Lu, Z.; Fleming, H.P. DNA Fingerprinting of Lactic Acid Bacteria in Sauerkraut Fermentations. Appl. Environ. Microbiol. 2007, 73, 7697–7702. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Reale, A. A Holistic Review on Euro-Asian Lactic Acid Bacteria Fermented Cereals and Vegetables. Microorganisms 2020, 8, 1176. [Google Scholar] [CrossRef] [PubMed]
- De Castro, A.; Montaño, A.; Sánchez, A.H.; Rejano, L. Lactic acid fermentation and storage of blanched garlic. Int. J. Food Microbiol. 1998, 39, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Kim, S.J.; Lee, J.Y.; Yoon, S.R.; You, S.Y.; Kim, S.H. Multifunctional properties of Lactobacillus plantarum strains WiKim83 and WiKim87 as a starter culture for fermented food. Food Sci. Nutr. 2019, 7, 2505–2516. [Google Scholar] [CrossRef] [PubMed]
- Lorn, D.; Nguyen, T.-K.-C.; Ho, P.-H.; Tan, R.; Licandro, H.; Waché, Y. Screening of lactic acid bacteria for their potential use as aromatic starters in fermented vegetables. Int. J. Food Microbiol. 2021, 350, 109242. [Google Scholar] [CrossRef] [PubMed]
- Maoloni, A.; Ferrocino, I.; Milanović, V.; Cocolin, L.; Corvaglia, M.R.; Ottaviani, D.; Bartolini, C.; Talevi, G.; Belleggia, L.; Cardinali, F.; et al. The Microbial Diversity of Non-Korean Kimchi as Revealed by Viable Counting and Metataxonomic Sequencing. Foods 2020, 9, 1568. [Google Scholar] [CrossRef]
- Montaño, A.; Sánchez, A.H.; Rejano, L.; de Castro, A. Processing and storage of lye-treated carrots fermented by a mixed starter culture. Int. J. Food Microbiol. 1997, 35, 83–90. [Google Scholar] [CrossRef]
- Oguntoyinbo, F.A.; Cho, G.S.; Trierweiler, B.; Kabisch, J.; Rösch, N.; Neve, H.; Bockelmann, W.; Frommherz, L.; Nielsen, D.S.; Krych, L.; et al. Fermentation of African kale (Brassica carinata) using L. plantarum BFE 5092 and L. fermentum BFE 6620 starter strains. Int. J. Food Microbiol. 2016, 238, 103–112. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Paradiso, A.; De Angelis, M.; Salmon, J.-C.; Buchin, S.; De Gara, L.; Gobbetti, M. Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 2009, 128, 473–483. [Google Scholar] [CrossRef]
- Cheng, L.; Luo, J.; Li, P.; Yu, H.; Huang, J.; Luo, L. Microbial diversity and flavor formation in onion fermentation. Food Funct. 2014, 5, 2338–2347. [Google Scholar] [CrossRef]
- Zhai, Y.; Pérez-Díaz, I.M. Identification of potential causative agents of the CO2-mediated bloater defect in low salt cucumber fermentation. Int. J. Food Microbiol. 2021, 344, 109115. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Fleming, H.P.; Hassan, H.M. Acid Tolerance of Leuconostoc mesenteroides and Lactobacillus plantarum. Appl. Environ. Microbiol. 1990, 56, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Ucar, R.A.; Pérez-Díaz, I.M.; Dean, L.L. Content of xylose, trehalose and l-citrulline in cucumber fermentations and utilization of such compounds by certain lactic acid bacteria. Food Microbiol. 2020, 91, 103454. [Google Scholar] [CrossRef] [PubMed]
- Fleming, H.P.; Daeschel, M.A.; McFeeters, R.F.; Pierson, M.D. Butyric Acid Spoilage of Fermented Cucumbers. J. Food Sci. 1989, 54, 636–639. [Google Scholar] [CrossRef]
- Franco, W.; Pérez-Díaz, I.M.; Johanningsmeier, S.D.; McFeeters, R.F. Characteristics of Spoilage-Associated Secondary Cucumber Fermentation. Appl. Environ. Microbiol. 2012, 78, 1273–1284. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Medina, E.; Page, C.A.; Johanningsmeier, S.D.; Daughtry, K.V.; Moeller, L. Prevention of microbes-induced spoilage in sodium chloride–free cucumber fermentations employing preservatives. J. Food Sci. 2022, 87, 5054–5069. [Google Scholar] [CrossRef]
- Fred, E.B.; Peterson, W.H.; Anderson, J.A. The characteristics of certain pentose-destroying bacteria, especially as it concerns their action on arabinose. J. Biol. Chem. 1921, 48, 385–412. [Google Scholar] [CrossRef]
- Zanoni, P.; Farrow, J.A.E.; Phillips, B.A.; Collins, M.D. Lactobacillus pentosus (Fred, Peterson, and Anderson) sp. nov., nom. rev. Int. J. Syst. Bacteriol. 1987, 37, 339–341. [Google Scholar] [CrossRef]
- Bringel, F.; Curk, M.-C.; Hubert, J.-C. Characterization of Lactobacilli by Southern-Type Hybridization with a Lactobacillus plantarum pyrDFE Probe. Int. J. Syst. Bacteriol. 1996, 46, 588–594. [Google Scholar] [CrossRef]
- Dellaglio, F.; Bottazzi, V.; Vescovo, M. Deoxyribonucleic Acid Homology Among Lactobacillus Species of the Subgenus Streptobacterium Orla-Jensen. Int. J. Syst. Bacteriol. 1975, 25, 160–172. [Google Scholar] [CrossRef]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA Gene Sequence Analysis and Multiplex PCR Assay with recA Gene-Derived Primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Díaz, I.M.; Johanningsmeier, S.D.; Anekella, K.; Pagán-Medina, C.G.; Méndez-Sandoval, L.; Arellano, C.; Price, R.; Daughtry, K.V.; Borges, M.; Bream, C.; et al. Genotypic and phenotypic diversity among Lactobacillus plantarum and Lactobacillus pentosus isolated from industrial scale cucumber fermentations. Food Microbiol. 2021, 94, 103652. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Beaz-Hidalgo, R.; Hossain, M.J.; Liles, M.R. Taxonomic Affiliation of New Genomes Should Be Verified Using Average Nucleotide Identity and Multilocus Phylogenetic Analysis. Genome Announc. 2014, 2, e00927-14. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
- Anekella, K.; Pérez-Díaz, I.M. Characterization of robust Lactobacillus plantarum and Lactobacillus pentosus starter cultures for environmentally friendly low-salt cucumber fermentations. J. Food Sci. 2020, 85, 3487–3497. [Google Scholar] [CrossRef]
- Fleming, H.P.; McFeeters, R.F.; Daeschel, M.A.; Humphries, E.G.; Thompson, R.L. Fermentation of Cucumbers in Anaerobic Tanks. J. Food Sci. 1988, 53, 127–133. [Google Scholar] [CrossRef]
- McDonald, L.C.; Shieh, D.-H.; Fleming, H.P.; McFeeters, R.F.; Thompson, R.L. Evaluation of malolactic-deficient strains of Lactobacillus plantarum for use in cucumber fermentations. Food Microbiol. 1993, 10, 489–499. [Google Scholar] [CrossRef]
- Fideler Moore, J. Generation of Bioactive Peptides and ɣ-Aminobutyric Acid (GABA) during Natural Lactic Acid Fermentation of Cucumber. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2021. Available online: https://www.lib.ncsu.edu/resolver/1840.20/38713 (accessed on 1 June 2023).
- Page, C.A.; Pérez-Díaz, I.M. Whole-Genome Sequencing and Annotation of Selected Lactobacillales Isolated from Commercial Cucumber Fermentation. Microbiol. Resour. Announc. 2021, 10, e0062521. [Google Scholar] [CrossRef]
- Markowitz, V.M.; Chen, I.-M.A.; Palaniappan, K.; Chu, K.; Szeto, E.; Grechkin, Y.; Ratner, A.; Jacob, B.; Huang, J.; Williams, P.; et al. IMG: The integrated microbial genomes database and comparative analysis system. Nucleic Acids Res. 2012, 40, D115–D122. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2017, 9, 5114. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Meth. Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Alanjary, M.; Steinke, K.; Ziemert, N. AutoMLST: An automated web server for generating multi-locus species trees highlighting natural product potential. Nucleic Acids Res. 2019, 47, W276–W282. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding data and analysis capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Russel, J.; Pinilla-Redondo, R.; Mayo-Muñoz, D.; Shah, S.A.; Sørensen, S.J. CRISPRCasTyper: Automated Identification, Annotation, and Classification of CRISPR-Cas Loci. CRISPR J. 2020, 3, 462–469. [Google Scholar] [CrossRef]
- Nethery, M.A.; Barrangou, R. Predicting and visualizing features of CRISPR–Cas systems. Methods Enzymol. 2019, 616, 323–362. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef]
- Davidson, B.E.; Kordias, N.; Dobos, M.; Hillier, A.J. Genomic organization of lactic acid bacteria. Antonie Leeuwenhoek 1996, 70, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Yampolsky, L.Y. Mutation and genome evolution. In Encyclopedia of Evolutionary Biology; Kliman, E.D., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 77–83. [Google Scholar] [CrossRef]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Bobay, L.-M.; Ochman, H. The Evolution of Bacterial Genome Architecture. Front. Genet. 2017, 8, 72. [Google Scholar] [CrossRef]
- Sela, I.; Wolf, Y.I.; Koonin, E.V. Theory of prokaryotic genome evolution. Proc. Natl. Acad. Sci. USA 2016, 113, 11399–11407. [Google Scholar] [CrossRef]
- Westoby, M.; Aagren, D.; Gillings, M.R.; Litchman, E.; Madin, J.S.; Paulsen, I.T.; Tetu, S.G. Cell size, genome size, and maximum growth rate are near-independent dimensions of ecological variation across bacteria and archaea. Ecol. Evol. 2021, 11, 3956–3976. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Daeschel, M.A.; McFeeters, R.F.; Fleming, H.P. Modification of lactic acid bacteria for cucumber fermentations: Elimination of carbon dioxide production from malate. Devel. Ind. Microbiol. 1985, 26, 339–346. [Google Scholar]
- Suo, C.; Yin, Y.; Wang, X.; Lou, X.; Song, D.; Wang, X.; Gu, Q. Effects of Lactobacillus plantarum ZJ316 on pig growth and pork quality. BMC Vet. Res. 2012, 8, 89. [Google Scholar] [CrossRef]
- Parlindungan, E.; May, B.K.; Jones, O.A.H. Metabolic Insights into the Effects of Nutrient Stress on Lactobacillus plantarum B21. Front. Mol. Biosci. 2019, 6, 75. [Google Scholar] [CrossRef]
- Golneshin, A.; Gor, M.-C.; Williamson, N.; Vezina, B.; Van, T.T.H.; May, B.K.; Smith, A.T. Discovery and characterisation of circular bacteriocin plantacyclin B21AG from Lactiplantibacillus plantarum B21. Heliyon 2020, 6, e04715. [Google Scholar] [CrossRef]
- Barrangou, R.; Dudley, E.G. CRISPR-Based Typing and Next-Generation Tracking Technologies. Annu. Rev. Food Sci. Technol. 2016, 7, 395–411. [Google Scholar] [CrossRef]
- Horvath, P.; Romero, D.A.; Coûté-Monvoisin, A.C.; Richards, M.; Deveau, H.; Moineau, S.; Boyaval, P.; Fremaux, C.; Barrangou, R. Diversity, Activity, and Evolution of CRISPR Loci in Streptococcus thermophilus. J. Bacteriol. 2008, 190, 1401–1412. [Google Scholar] [CrossRef]
- Shariat, N.; DiMarzio, M.J.; Yin, S.; Dettinger, L.; Sandt, C.H.; Lute, J.R.; Barrangou, R.; Dudley, E.G. The combination of CRISPR-MVLST and PFGE provides increased discriminatory power for differentiating human clinical isolates of Salmonella enterica subsp. enterica serovar Enteritidis. Food Microbiol. 2013, 34, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Toro, M.; Cao, G.; Ju, W.; Allard, M.; Barrangou, R.; Zhao, S.; Brown, E.; Meng, J. Association of Clustered Regularly Interspaced Short Palindromic Repeat (CRISPR) Elements with Specific Serotypes and Virulence Potential of Shiga Toxin-Producing Escherichia coli. Appl. Environ. Microbiol. 2014, 80, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Pérez-Díaz, I.M.; Hayes, J.S.; Breidt, F. Bacteriophage Ecology in a Commercial Cucumber Fermentation. Appl. Environ. Microbiol. 2012, 78, 8571–8578. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Breidt, F.; Plengvidhya, V.; Fleming, H.P. Bacteriophage Ecology in Commercial Sauerkraut Fermentations. Appl. Environ. Microbiol. 2003, 69, 3192–3202. [Google Scholar] [CrossRef]
- Ramisetty, B.C.M.; Sudhakari, P.A. Bacterial “Grounded” Prophages: Hotspots for Genetic Renovation and Innovation. Front. Genet. 2019, 10, 65. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Accession Number | Strain | Accession Number |
|---|---|---|---|
| Lactiplantibacillus plantarum | |||
| CMPG5300 | GCA_000762955.1 | JDM1 | GCA_000023085.1 |
| B21 | GCA_000931425.2 | DOMLa | GCA_000604105.1 |
| WCFS1 | GCA_000203855. | 16 | GCA_000412205.1 |
| ZJ316 | GCA_000338115.2 | P-8 | GCA_000392485.2 |
| ATCC14917 | GCA_000143745.1 | 7.8.4 | GCA_018993285.1 |
| LPL-1 | GCA_002205775.2 | 3.2.8 | GCA_018991195.1 |
| ST-III | GCA_000148815.2 | T1R2b | GCA_018993325.1 |
| CGMCC1.557 | GCA_001272315.2 | ||
| Lactiplantibacillus pentosus | |||
| BGM48 | GCA_002850015.1 | 1.8.6 | GCA_018991355.1 |
| FL0421 | GCA_001188985.1 | 1.8.9 | GCA_018993945.1 |
| ZFM222 | GCA_003627295.1 | 1.8.18 | GCA_018993635.1 |
| MP-10 | GCA_900092635.1 | 3.2.37 | GCA_018993495.1 |
| KCA1 | GCA_000271445.1 | 3.2.36 | GCA_018993725.1 |
| RI-031 | GCA_002751855.1 | 3.8.24 | GCA_018993345.1 |
| MS031 | GCA_016804305.1 | 3.8.45 | GCA_018991565.1 |
| SLC13 | GCA_002211885.1 | 7.2.4 | GCA_018991185.1 |
| IG2 | GCA_002993465.1 | 7.2.11 | GCA_018991375.1 |
| IG3 | GCA_002993425.1 | 7.2.15 | GCA_018993445.1 |
| IG4 | GCA_002993385.1 | 7.2.20 | GCA_018993665.1 |
| IG5 | GCA_002993435.1 | 7.2.23 | GCA_018993485.1 |
| IG6 | GCA_002993485.1 | 7.8.2 | GCA_018991535.1 |
| IG7 | GCA_002993395.1 | 7.8.11 | GCA_018993275.1 |
| IG8 | GCA_003702625.1 | 7.8.46 | GCA_018991465.1 |
| IG9 | GCA_003702665.1 | 14.8.42 | GCA_018993525.1 |
| IG10 | GCA_003702635.1 | 14.2.3 | GCA_018991425.1 |
| IG11 | GCA_003702605.1 | 14.2.16 | GCA_018993625.1 |
| IG12 | GCA_003702565.1 | 30.2.29 | GCA_018991255.1 |
| 1.2.7 | GCA_018991365.1 | LA0445 | GCA_018993965.1 |
| 1.2.11 | GCA_018991285.1 | MU045 | GCA_018993585.1 |
| 1.2.13 | GCA_018991345.1 | ||
| Weissella spp. | Leuconostoc spp. | ||
| W. paramesenteroides 3.2.24 | GCA_018991665.1 | Leu. citreum 3.8.12 | GCA_018991475.1 |
| W. cibaria 7.8.34 | GCA_025770435.1 | Leu. lactis 1.2.50 | GCA_018993865.1 |
| W. cibaria 3.8.44 | GCA_018993825.1 | Leu. fallax 1.2.22 | GCA_018993745.1 |
| W. sagaensis 1.2.50 | GCA_018993865.1 | Leu. mesenteroides 1.2.47 | GCA_018991785.1 |
| Pediococcus spp. | Lactococcus spp. | ||
| P. ethanolidurans 90.8.30 | GCA_018993685.1 | Lc. lactis LA0312 | GCA_018991605.1 |
| P. damnosus DSM 20331 | GCA_001437255.1 | Lc. lactis 1.8.12 | GCA_018991645.1 |
| Levilactobacillus spp. | |||
| Lev. brevis 14.2.24 | GCA_018993885.1 | Lactiplantibacillus paraplantarum L-ZS9 | GCA_001443645.1 |
| Lev. brevis 14.2.10 | GCA_018991505.1 | ||
| Lev. brevis 3.2.41 | GCA_018991675.1 | ||
| Genomic DNA Sequence Identification | ||||||||
|---|---|---|---|---|---|---|---|---|
| L. plantarum | L. pentosus | |||||||
| 3.2.8 | 7.8.4 | T1R2b | Subclade A.I | Subclade A.II | Subclade A.III | Clade B | 7.8.2 | |
| CRISPR loci | 1 | 1 | 0 | 4 ± 1 | 4 | 4 ± 1 | 2 ± 1 | 5 |
| Type I-E | 0 | 0 | 0 | 2 | 3 | 2 | 2 ± 1 | 4 |
| Type II-A | 1 | 1 | 0 | 2 ± 1 | 1 | 1 ± 1 | 0 | 1 |
| Closest Homolog Identity (>95%) | L. plantarum Isolates Autochthonous to Cucumber Fermentation | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3.2.8 | 7.8.4 | T1R2b | ||||||||||||||||||||||
| Siphoviridae sp. ctu0P1 | X | |||||||||||||||||||||||
| Siphoviridae sp. ctyPQ2 | X | |||||||||||||||||||||||
| Siphoviridae sp. ct97I3 | X | |||||||||||||||||||||||
| Siphoviridae sp. ctk5O4 | ||||||||||||||||||||||||
| Siphoviridae sp. ctUZT2 | X | |||||||||||||||||||||||
| Closest Homolog Identity (>95%) | L. pentosus Isolates Autochthonous to Cucumber Fermentation | |||||||||||||||||||||||
| Subclade A.I | Subclade A.II | Subclade A.III | Clade B | |||||||||||||||||||||
| 1.2.7 | 1.2.13 | 3.8.24 | 1.8.6 | 1.2.11 | 3.2.37 | 30.2.29 | 1.8.9 | 7.2.23 | 3.2.36 | 3.8.45 | 1.8.18 | 7.8.46 | 7.2.20 | 14.2.3 | 7.2.15 | 14.2.16 | 14.8.42 | 7.8.11 | 7.2.11 | 7.2.4 | LA0445 | MU0045 | 7.8.2 | |
| Siphoviridae ctu0P1 | X | X | X | X | X | X | ||||||||||||||||||
| Siphoviridae ctyPQ2 | ||||||||||||||||||||||||
| Siphoviridae ct97I3 | ||||||||||||||||||||||||
| Siphoviridae ctk5O4 | X | |||||||||||||||||||||||
| Siphoviridae ctUZT2 | ||||||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Page, C.A.; Pérez-Díaz, I.M.; Pan, M.; Barrangou, R. Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry. Foods 2023, 12, 2455. https://doi.org/10.3390/foods12132455
Page CA, Pérez-Díaz IM, Pan M, Barrangou R. Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry. Foods. 2023; 12(13):2455. https://doi.org/10.3390/foods12132455
Chicago/Turabian StylePage, Clinton A., Ilenys M. Pérez-Díaz, Meichen Pan, and Rodolphe Barrangou. 2023. "Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry" Foods 12, no. 13: 2455. https://doi.org/10.3390/foods12132455
APA StylePage, C. A., Pérez-Díaz, I. M., Pan, M., & Barrangou, R. (2023). Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry. Foods, 12(13), 2455. https://doi.org/10.3390/foods12132455