
Differences in Bacterial Communities of Retail Raw Pork in Different Market Types in Hangzhou, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Culture-Dependent Bacterial Isolation and Identification
2.3. Antimicrobial Susceptibility Testing
2.4. NaClO Tolerance Determinations
2.5. DNA Extraction
2.6. PCR Amplification and Sequencing
2.7. Sequence Data Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Prevalence, Antibiotic and Chlorine Resistance of Foodborne Pathogens
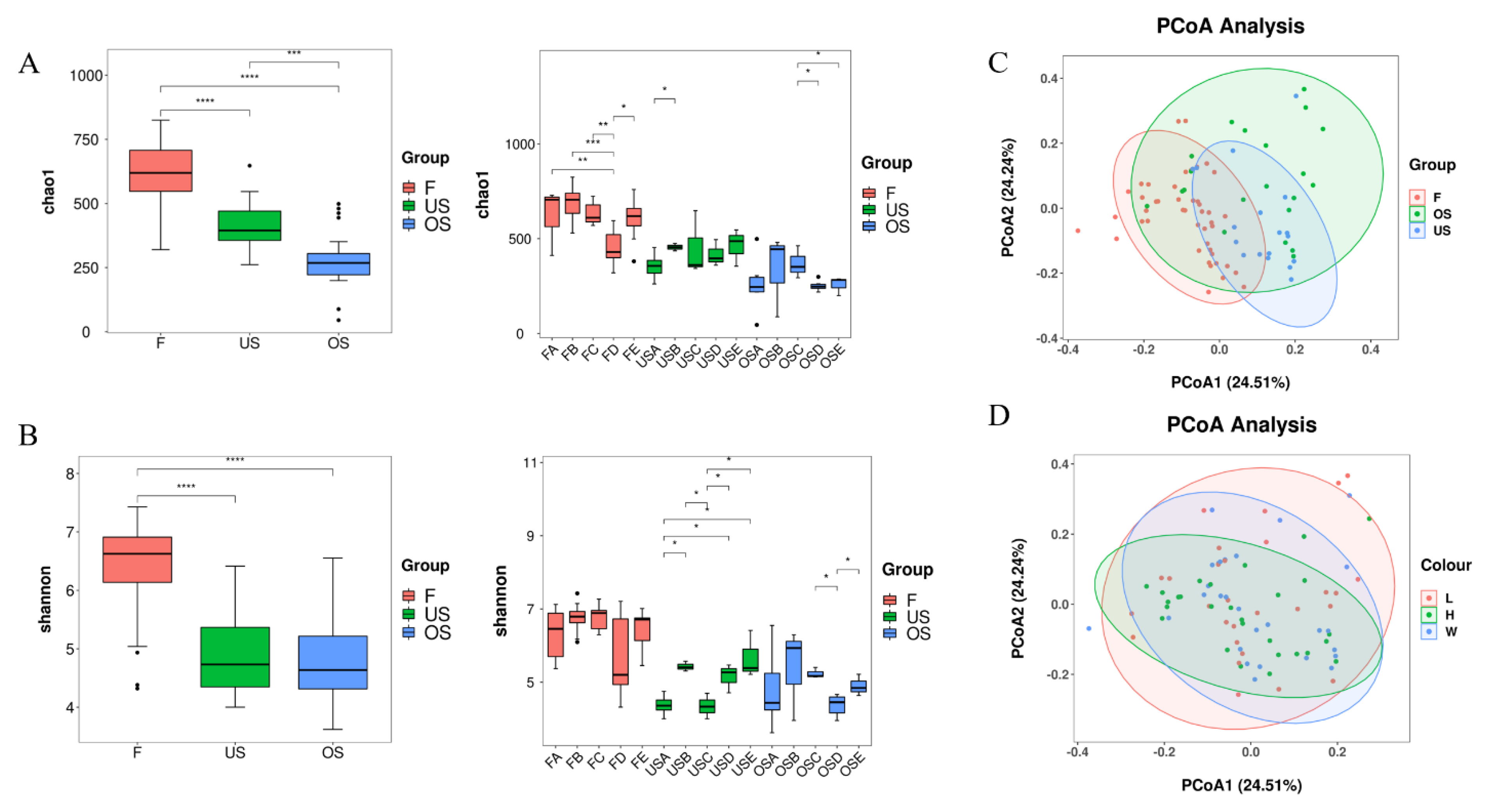
3.2. Bacterial Community Richness and Diversity
3.3. Comparison of Bacterial Communities of Different Types of Pork Markets
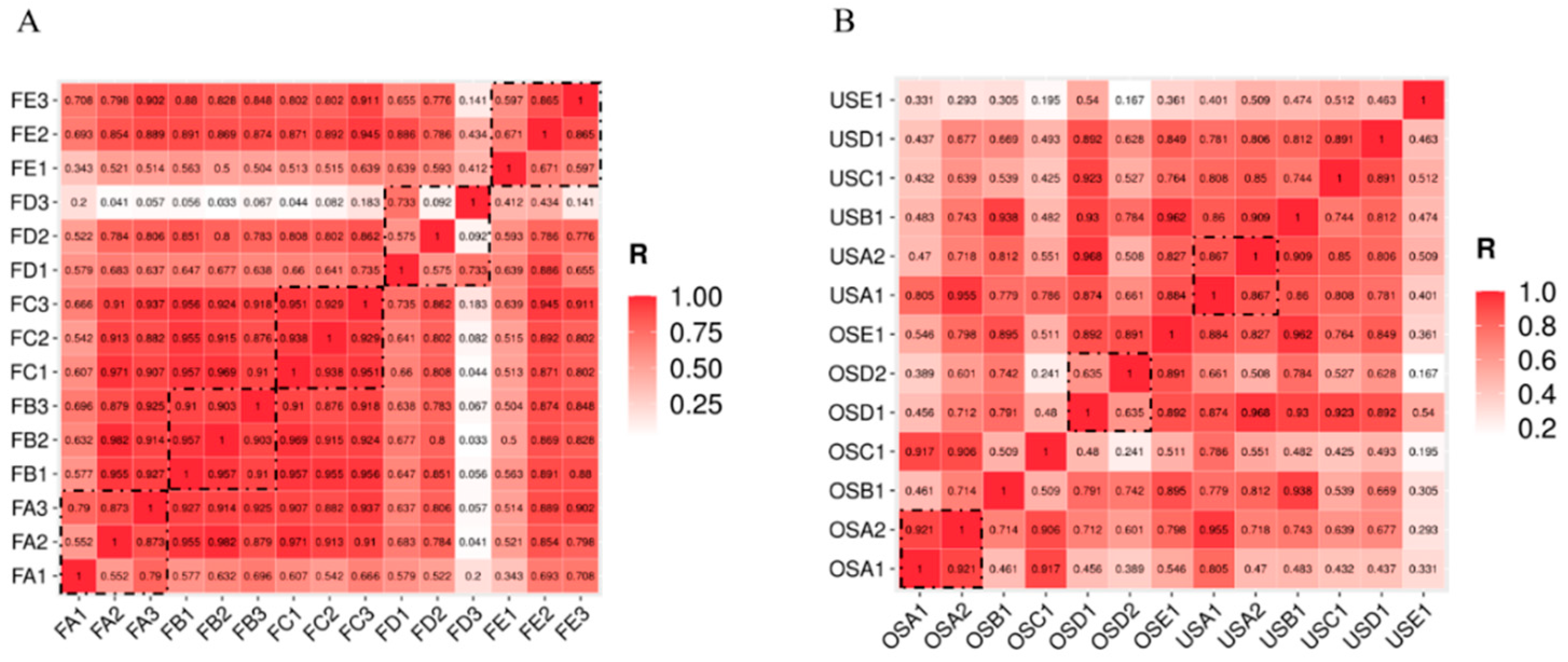
3.4. Bacterial Cross-Contamination in the Pork Market
3.5. Microbial Function Prediction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lambert, A.D.; Smith, J.P.; Dodds, K.L. Shelf life extension and microbiological safety of fresh meat—A review. Food Microbiol. 1991, 8, 267–297. [Google Scholar] [CrossRef]
- Liu, C.-X.; Xiao, Y.-P.; Hu, D.-W.; Liu, J.-X.; Chen, W.; Ren, D.-X. The safety evaluation of chilled pork from online platform in China. Food Control. 2019, 96, 244–250. [Google Scholar] [CrossRef]
- Zhu, Z.; Huang, Q.; Hong, X.; Chen, X.; Lu, Y.; Chen, Z.; Wang, C.; Meng, X.; Xu, Q.; Li, S. Isolation and characterization of Salmonella in pork samples collected from retail and wholesale markets in each season from 2016 to 2018 in Wuhan, China. J. Appl. Microbiol. 2019, 128, 875–883. [Google Scholar] [CrossRef]
- Hu, Z.; Peng, Z.; Zhang, X.; Li, Z.; Jia, C.; Li, X.; Wang, X. Prevalence and Molecular Characterization of Antimicrobial-Resistant Escherichia coli in Pig Farms, Slaughterhouses, and Terminal Markets in Henan Province of China. Foodborne Pathog. Dis. 2021, 18, 733–743. [Google Scholar] [CrossRef]
- Andritsos, N.D.; Mataragas, M.; Mavrou, E.; Stamatiou, A.; Drosinos, E.H. The microbiological condition of minced pork prepared at retail stores in Athens, Greece. Meat Sci. 2012, 91, 486–489. [Google Scholar] [CrossRef]
- Braley, C.; Fravalo, P.; Gaucher, M.-L.; Larivière-Gauthier, G.; Shedleur-Bourguignon, F.; Longpré, J.; Thibodeau, A. Similar Carcass Surface Microbiota Observed Following Primary Processing of Different Pig Batches. Front. Microbiol. 2022, 13, 849883. [Google Scholar] [CrossRef]
- Qiu, M.; Xiao, X.; Xiao, Y.; Ma, J.; Yang, H.; Jiang, H.; Dong, Q.; Wang, W. Dynamic Changes of Bacterial Communities and Microbial Association Networks in Ready-to-Eat Chicken Meat during Storage. Foods 2022, 11, 3733. [Google Scholar] [CrossRef]
- Ray, S. Sensory Properties of Foods and Their Measurement Methods. In Techniques to Measure Food Safety and Quality; Springer: Berlin/Heidelberg, Germany, 2021; pp. 345–381. [Google Scholar] [CrossRef]
- Li, H.; Li, W.; Dai, Y.; Jiang, Y.; Liang, J.; Wang, S.; Zhuang, M.; Huang, Z.; Xu, L.; Xue, B.; et al. Characteristics of Settings and Etiologic Agents of Foodborne Disease Outbreaks—China, 2020. China CDC Wkly. 2021, 3, 889–893. [Google Scholar] [CrossRef]
- Van Cuong, N.; Nhung, N.T.; Nghia, N.H.; Hoa, N.T.M.; Trung, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial Consumption in Medicated Feeds in Vietnamese Pig and Poultry Production. EcoHealth 2016, 13, 490–498. [Google Scholar] [CrossRef]
- Nhung, N.T.; Van, N.T.B.; Van Cuong, N.; Duong, T.T.Q.; Nhat, T.T.; Hang, T.T.T.; Nhi, N.T.H.; Kiet, B.T.; Hien, V.B.; Ngoc, P.T.; et al. Antimicrobial residues and resistance against critically important antimicrobials in non-typhoidal Salmonella from meat sold at wet markets and supermarkets in Vietnam. Int. J. Food Microbiol. 2018, 266, 301–309. [Google Scholar] [CrossRef]
- Boss, S.; Stephan, R.; Horlbog, J.A.; Magouras, I.; Colon, V.A.; Lugsomya, K.; Nüesch-Inderbinen, M. Serotypes, Antimicrobial Resistance Profiles, and Virulence Factors of Salmonella Isolates in Chinese Edible Frogs (Hoplobatrachus rugulosus) Collected from Wet Markets in Hong Kong. Foods 2023, 12, 2245. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Boto, D.; D’arrigo, M.; García-Lafuente, A.; Bravo, D.; Pérez-Baltar, A.; Gaya, P.; Medina, M.; Arqués, J.L. Staphylococcus aureus in the Processing Environment of Cured Meat Products. Foods 2023, 12, 2161. [Google Scholar] [CrossRef] [PubMed]
- Templeton, M.R.; Oddy, F.; Leung, W.-K.; Rogers, M. Chlorine and UV disinfection of ampicillin-resistant and trimethoprim-resistantEscherichia coli. J. Environ. Eng. Sci. 2013, 8, 231–237. [Google Scholar] [CrossRef]
- Wu, G.; Yang, Q.; Long, M.; Guo, L.; Li, B.; Meng, Y.; Zhang, A.; Wang, H.; Liu, S.; Zou, L. Evaluation of agar dilution and broth microdilution methods to determine the disinfectant susceptibility. J. Antibiot. 2015, 68, 661–665. [Google Scholar] [CrossRef]
- Higgins, D.; Pal, C.; Sulaiman, I.M.; Jia, C.; Zerwekh, T.; Dowd, S.E.; Banerjee, P. Application of high-throughput pyrosequencing in the analysis of microbiota of food commodities procured from small and large retail outlets in a U.S. metropolitan area – A pilot study. Food Res. Int. 2018, 105, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; Park, H.J.; Jang, H.I.; Kim, S.A.; Imm, J.Y.; Hwang, I.G.; Rhee, M.S. Changes in microbial contamination levels of porcine carcasses and fresh pork in slaughterhouses, processing lines, retail outlets, and local markets by commercial distribution. Res. Vet. Sci. 2013, 94, 413–418. [Google Scholar] [CrossRef]
- Shiningeni, D.; Chimwamurombe, P.; Shilangale, R.; Misihairabgwi, J. Prevalence of pathogenic bacteria in street vended ready-to-eat meats in Windhoek, Namibia. Meat Sci. 2019, 148, 223–228. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Alvarez-Molina, A.; Alexa, E.A.; Walsh, C.J.; Mencía-Ares, O.; Puente-Gómez, P.; Likotrafiti, E.; Fernández-Gómez, P.; Prieto, B.; Crispie, F.; et al. Microbial colonization and resistome dynamics in food processing environments of a newly opened pork cutting industry during 1.5 years of activity. Microbiome 2021, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Jin, X.; Zheng, H.; Li, J.; Meng, C.; Yin, K.; Xie, X.; Huang, C.; Lei, T.; Sun, X.; et al. The prevalence and load of Salmonella, and key risk points of Salmonella contamination in a swine slaughterhouse in Jiangsu province, China. Food Control. 2018, 87, 153–160. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, J.; Li, R.; Ye, K. High-throughput sequencing analysis of the bacterial community for assessing the differences in extraction methods of bacteria separation from chilled pork. LWT 2020, 134, 110213. [Google Scholar] [CrossRef]
- Jang, M.J.; Kim, S.Y.; Ricke, S.C.; Rhee, M.S.; Kim, S.A. Microbial ecology of alfalfa, radish, and rapeseed sprouts based on culture methods and 16S rRNA microbiome sequencing. Food Res. Int. 2021, 144, 110316. [Google Scholar] [CrossRef]
- Bassey, A.P.; Chen, Y.; Zhu, Z.; Odeyemi, O.A.; Frimpong, E.B.; Ye, K.; Li, C.; Zhou, G. Assessment of quality characteristics and bacterial community of modified atmosphere packaged chilled pork loins using 16S rRNA amplicon sequencing analysis. Food Res. Int. 2021, 145, 110412. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, W.; Zhai, Z.; Shi, H.; Luo, Y.; Chen, Z.; Huang, K. Universal primer-multiplex PCR approach for simultaneous detection of Escherichia coli, Listeria monocytogenes, and Salmonella spp. in food samples. J. Food. Sci. 2009, 74, M446–M452. [Google Scholar] [CrossRef]
- Zhang, L.; Fu, Y.; Xiong, Z.; Ma, Y.; Wei, Y.; Qu, X.; Zhang, H.; Zhang, J.; Liao, M. Highly Prevalent Multidrug-Resistant Salmonella from Chicken and Pork Meat at Retail Markets in Guangdong, China. Front. Microbiol. 2018, 9, 2104. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, Approved Standard, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Xiao, X.; Bai, L.; Wang, S.; Liu, L.; Qu, X.; Zhang, J.; Xiao, Y.; Tang, B.; Li, Y.; Yang, H.; et al. Chlorine Tolerance and Cross-Resistance to Antibiotics in Poultry-Associated Salmonella Isolates in China. Front. Microbiol. 2021, 12, 833743. [Google Scholar] [CrossRef]
- Niamah, A.K. Detected of Aero Gene in Aeromonas Hydrophila Isolates from Shrimp and Peeled Shrimp Samples in Local Markets. J. Microbiol. Biotechnol. Food Sci. 2012, 2, 634. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- De Filippis, F.; La Storia, A.; Villani, F.; Ercolini, D. Strain-Level Diversity Analysis of Pseudomonas fragi after In Situ Pangenome Reconstruction Shows Distinctive Spoilage-Associated Metabolic Traits Clearly Selected by Different Storage Conditions. Appl. Environ. Microbiol. 2018, 85. [Google Scholar] [CrossRef]
- Lai, Z.; Lin, L.; Zhang, J.; Mao, S. Effects of high-grain diet feeding on mucosa-associated bacterial community and gene expression of tight junction proteins and inflammatory cytokines in the small intestine of dairy cattle. J. Dairy Sci. 2022, 105, 6601–6615. [Google Scholar] [CrossRef]
- Cai, Y.; Tao, J.; Jiao, Y.; Fei, X.; Zhou, L.; Wang, Y.; Zheng, H.; Pan, Z.; Jiao, X. Phenotypic characteristics and genotypic correlation between Salmonella isolates from a slaughterhouse and retail markets in Yangzhou, China. Int. J. Food Microbiol. 2016, 222, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Anwar, T.M.; Peng, X.; Biswas, S.; Elbediwi, M.; Li, Y.; Fang, W.; Yue, M. Prevalence and antimicrobial resistance of Salmonella recovered from pig-borne food products in Henan, China. Food Control. 2021, 121, 107535. [Google Scholar] [CrossRef]
- Fang, J.; Shen, Y.; Qu, D.; Han, J. Antimicrobial resistance profiles and characteristics of integrons in Escherichia coli strains isolated from a large-scale centralized swine slaughterhouse and its downstream markets in Zhejiang, China. Food Control. 2019, 95, 215–222. [Google Scholar] [CrossRef]
- Aregbe, A.Y.; Mu, T.; Sun, H. Effect of different pretreatment on the microbial diversity of fermented potato revealed by high-throughput sequencing. Food Chem. 2019, 290, 125–134. [Google Scholar] [CrossRef]
- Li, N.; Zhang, Y.; Wu, Q.; Gu, Q.; Chen, M.; Zhang, Y.; Sun, X.; Zhang, J. High-throughput sequencing analysis of bacterial community composition and quality characteristics in refrigerated pork during storage. Food Microbiol. 2019, 83, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wu, H.; Zhang, C.; Jie, J.; Liu, Z.; Zeng, M.; Wang, C. Spoilage of refrigerated Litopenaeus vannamei: Eavesdropping on Acinetobacter acyl-homoserine lactones promotes the spoilage potential of Shewanella baltica. J. Food Sci. Technol. 2018, 55, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Moen, B.; Heir, E.; Hansen, A.Å.; Langsrud, S. Contamination of salmon fillets and processing plants with spoilage bacteria. Int. J. Food Microbiol. 2016, 237, 98–108. [Google Scholar] [CrossRef]
- Li, X.; Xiong, Q.; Xu, B.; Wang, H.; Zhou, H.; Sun, Y. Bacterial community dynamics during different stages of processing of smoked bacon using the 16S rRNA gene amplicon analysis. Int. J. Food Microbiol. 2021, 351, 109076. [Google Scholar] [CrossRef] [PubMed]
- Stellato, G.; La Storia, A.; De Filippis, F.; Borriello, G.; Villani, F.; Ercolini, D. Overlap of spoilage microbiota between meat and meat processing environment in small-scale vs large-scale retail distribution. Appl. Environ. Microbiol. 2016, 82, 4045–4054. [Google Scholar] [CrossRef]
- Van Ba, H.; Seo, H.-W.; Seong, P.-N.; Kang, S.-M.; Cho, S.-H.; Kim, Y.-S.; Park, B.-Y.; Moon, S.-S.; Kang, S.-J.; Choi, Y.-M.; et al. The fates of microbial populations on pig carcasses during slaughtering process, on retail cuts after slaughter, and intervention efficiency of lactic acid spraying. Int. J. Food Microbiol. 2019, 294, 10–17. [Google Scholar] [CrossRef]
- Stanborough, T.; Fegan, N.; Powell, S.M.; Tamplin, M.; Chandry, P.S. Insight into the Genome of Brochothrix thermosphacta, a Problematic Meat Spoilage Bacterium. Appl. Environ. Microbiol. 2017, 83, e02786-16. [Google Scholar] [CrossRef]
- Remenant, B.; Jaffrès, E.; Dousset, X.; Pilet, M.-F.; Zagorec, M. Bacterial spoilers of food: Behavior, fitness and functional properties. Food Microbiol. 2015, 45, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Wei, Z.; Zhou, G.; Kristiansen, K.; Wang, C. Effects of Different Storage Temperatures on Bacterial Communities and Functional Potential in Pork Meat. Foods 2022, 11, 2307. [Google Scholar] [CrossRef]
- Moi, I.M.; Roslan, N.N.; Leow, A.T.C.; Ali, M.S.M.; Rahman, R.N.Z.R.A.; Rahimpour, A.; Sabri, S. The biology and the importance of Photobacterium species. Appl. Microbiol. Biotechnol. 2017, 101, 4371–4385. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, T.T.; Dalgaard, P.; Björkroth, J. Volatile organic compounds and Photobacterium phosphoreum associated with spoilage of modified-atmosphere-packaged raw pork. Int. J. Food Microbiol. 2016, 218, 86–95. [Google Scholar] [CrossRef]
- Samelis, J.; Kakouri, A.; Rementzis, J. Selective effect of the product type and the packaging conditions on the species of lactic acid bacteria dominating the spoilage microbial association of cooked meats at 4 °C. Food Microbiol. 2000, 17, 329–340. [Google Scholar] [CrossRef]
- Paudyal, N.; Anihouvi, V.; Hounhouigan, J.; Matsheka, M.I.; Sekwati-Monang, B.; Amoa-Awua, W.; Atter, A.; Ackah, N.B.; Mbugua, S.; Asagbra, A.; et al. Prevalence of foodborne pathogens in food from selected African countries—A meta-analysis. Int. J. Food Microbiol. 2017, 249, 35–43. [Google Scholar] [CrossRef]
- Asiegbu, C.V.; Lebelo, S.L.; Tabit, F.T. The food safety knowledge and microbial hazards awareness of consumers of ready-to-eat street-vended food. Food Control. 2016, 60, 422–429. [Google Scholar] [CrossRef]
- Cocolin, L.; Alessandria, V.; Dolci, P.; Gorra, R.; Rantsiou, K. Culture independent methods to assess the diversity and dynamics of microbiota during food fermentation. Int. J. Food Microbiol. 2013, 167, 29–43. [Google Scholar] [CrossRef]
- Duthoo, E.; Rasschaert, G.; Leroy, F.; Weckx, S.; Heyndrickx, M.; De Reu, K. The Microbiota of Modified-Atmosphere-Packaged Cooked Charcuterie Products throughout Their Shelf-Life Period, as Revealed by a Complementary Combination of Culture-Dependent and Culture-Independent Analysis. Microorganisms 2021, 9, 1223. [Google Scholar] [CrossRef]
- García-Patiño, M.G.; García-Contreras, R.; Licona-Limón, P. The Immune Response against Acinetobacter baumannii, an Emerging Pathogen in Nosocomial Infections. Front. Immunol. 2017, 8, 441. [Google Scholar] [CrossRef]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R., Jr.; Chen, Z.-S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updat. 2015, 18, 1–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Supermarket Type | Sampling Sites | Sample Code | Sample Size |
|---|---|---|---|
| Farmers’ Market | Farmers’ Market A | FA1 | 3 |
| FA2 | 3 | ||
| FA3 | 3 | ||
| Farmers’ Market B | FB1 | 3 | |
| FB2 | 3 | ||
| FB3 | 3 | ||
| Farmers’ Market C | FC1 | 3 | |
| FC2 | 3 | ||
| FC3 | 3 | ||
| Farmers’ Market D | FD1 | 3 | |
| FD2 | 3 | ||
| FD3 | 3 | ||
| Farmers’ Market E | FE1 | 3 | |
| FE2 | 3 | ||
| FE3 | 3 | ||
| Upscale Supermarket | Upscale Supermarket A | USA1 | 3 |
| USA2 | 3 | ||
| Upscale Supermarket B | USB | 3 | |
| Upscale Supermarket C | USC | 3 | |
| Upscale Supermarket D | USD | 3 | |
| Upscale Supermarket E | USE | 3 | |
| Ordinary Supermarket | Ordinary Supermarket A | OSA1 | 3 |
| OSA2 | 3 | ||
| Ordinary Supermarket B | OSB | 3 | |
| Ordinary Supermarket C | OSC | 3 | |
| Ordinary Supermarket D | OSD1 | 3 | |
| OSD2 | 3 | ||
| Ordinary Supermarket E | OSE | 3 |
| Sample Code | Sample Size | E. coli | Salmonella | |||||
|---|---|---|---|---|---|---|---|---|
| Prevalence (%) * | MDR (%) # | NaClO Tolerance (%) # | Prevalence (%) * | MDR (%) # | NaClO Tolerance (%) # | |||
| Farmers’ Market | FA | 9 | 0 (0%) | - | - | 1 (11.1%) | 0 | - |
| FB | 9 | 2 (22.2%) | 0 | - | 1 (11.1%) | 1 (100%) | 1 (100%) | |
| FC | 9 | 4 (44.4%) | 0 | - | 4 (44.4%) | 2 (100%) | 4 (100%) | |
| FD | 9 | 1 (11.1%) | 1 (100%) | 1 (100%) | 3 (30%) | 2 (66.7%) | 3 (100%) | |
| FE | 3 | 2 (22.2%) | 2 (100%) | 1 (50%) | 3 (30%) | 1 (33.3%) | 3 (100%) | |
| Upscale Supermarket | USA | 6 | 3 (50%) | 2 (66.7%) | 2 (66.7%) | 0 (0%) | - | - |
| USB | 3 | 2 (66.7%) | 0 | - | 0 (0%) | - | - | |
| USC | 3 | 2 (66.7%) | 1 (50%) | - | 0 (0%) | - | - | |
| USD | 3 | 0 (0%) | - | - | 0 (0%) | - | - | |
| USE | 3 | 1 (33.3%) | 1 (100%) | 0 (0%) | - | - | ||
| Ordinary Supermarket | OSA | 6 | 2 (33.3%) | 1 (50%) | - | 1 (16.7%) | 1 (100%) | 1 (100%) |
| OSB | 3 | 0 (0%) | - | - | 0 (0%) | - | - | |
| OSC | 3 | 1 (33.3%) | 0 | 1 (33.3%) | 0 (0%) | - | - | |
| OSD | 6 | 6 (100%) | 6 (100%) | 3 (50%) | 0 (0%) | - | - | |
| OSE | 3 | 1 (33.3%) | 0 | 1 (100%) | 0 (0%) | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Yi, Z.; Cai, W.; Ma, J.; Yang, H.; Zhou, M.; Xiao, X. Differences in Bacterial Communities of Retail Raw Pork in Different Market Types in Hangzhou, China. Foods 2023, 12, 3357. https://doi.org/10.3390/foods12183357
Wang W, Yi Z, Cai W, Ma J, Yang H, Zhou M, Xiao X. Differences in Bacterial Communities of Retail Raw Pork in Different Market Types in Hangzhou, China. Foods. 2023; 12(18):3357. https://doi.org/10.3390/foods12183357
Chicago/Turabian StyleWang, Wen, Zhengkai Yi, Wei Cai, Jiele Ma, Hua Yang, Min Zhou, and Xingning Xiao. 2023. "Differences in Bacterial Communities of Retail Raw Pork in Different Market Types in Hangzhou, China" Foods 12, no. 18: 3357. https://doi.org/10.3390/foods12183357
APA StyleWang, W., Yi, Z., Cai, W., Ma, J., Yang, H., Zhou, M., & Xiao, X. (2023). Differences in Bacterial Communities of Retail Raw Pork in Different Market Types in Hangzhou, China. Foods, 12(18), 3357. https://doi.org/10.3390/foods12183357