
Rabbit Meat Extract Induces Browning in 3T3−L1 Adipocytes via the AMP−Activated Protein Kinase Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rabbit Meat Preparation
2.2. Cell Culture and Differentiation
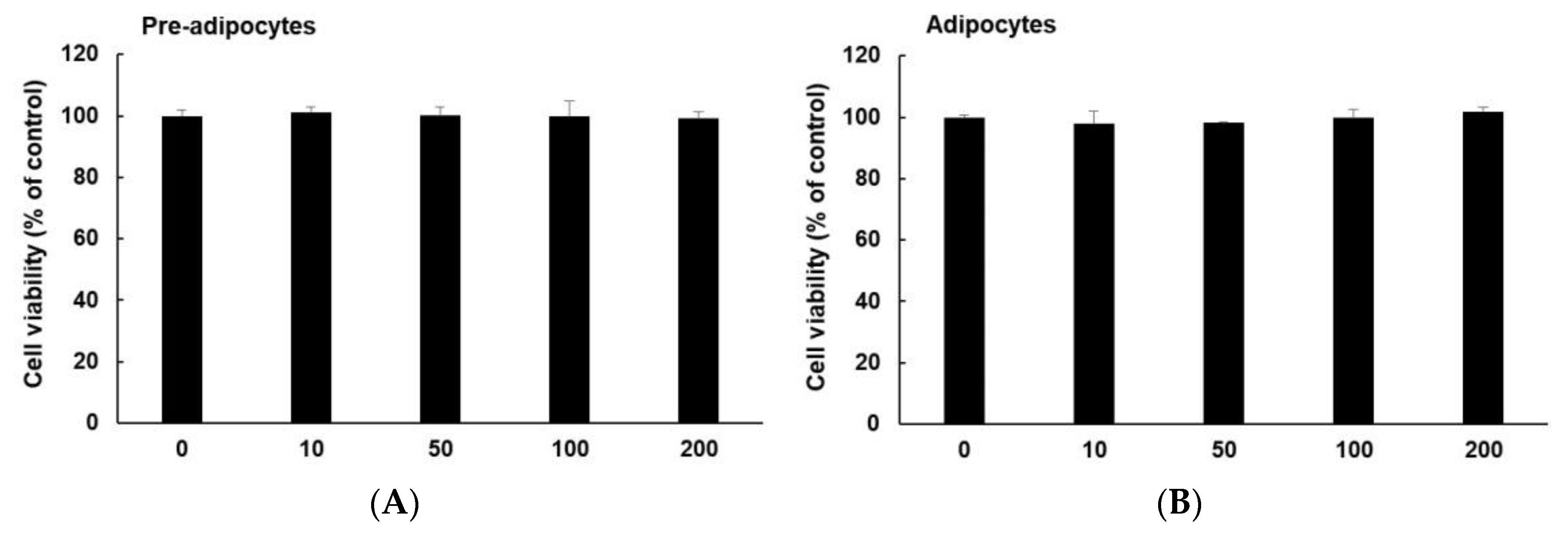
2.3. Cell Viability Assay
2.4. Oil Red O Staining
2.5. RNA Extraction and Quantitative Reverse−Transcription Polymerase Chain Reaction (qRT−PCR)
2.6. Western Blot Analysis
2.7. Glucose Uptake
2.8. Statistical Analysis
3. Results and Discussion
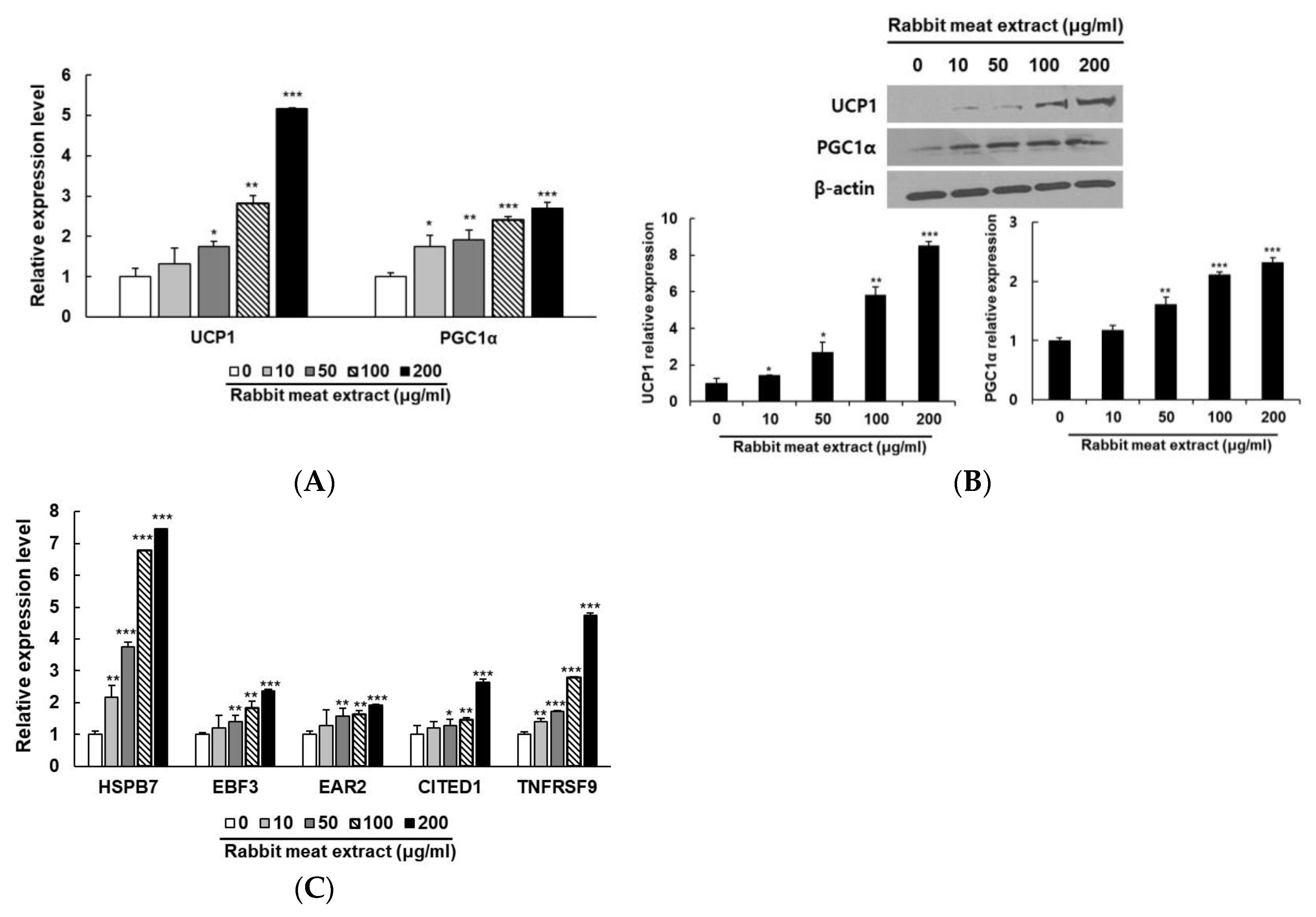
3.1. Rabbit Meat Extract Stimulated Browning in 3T3−L1 Adipocytes
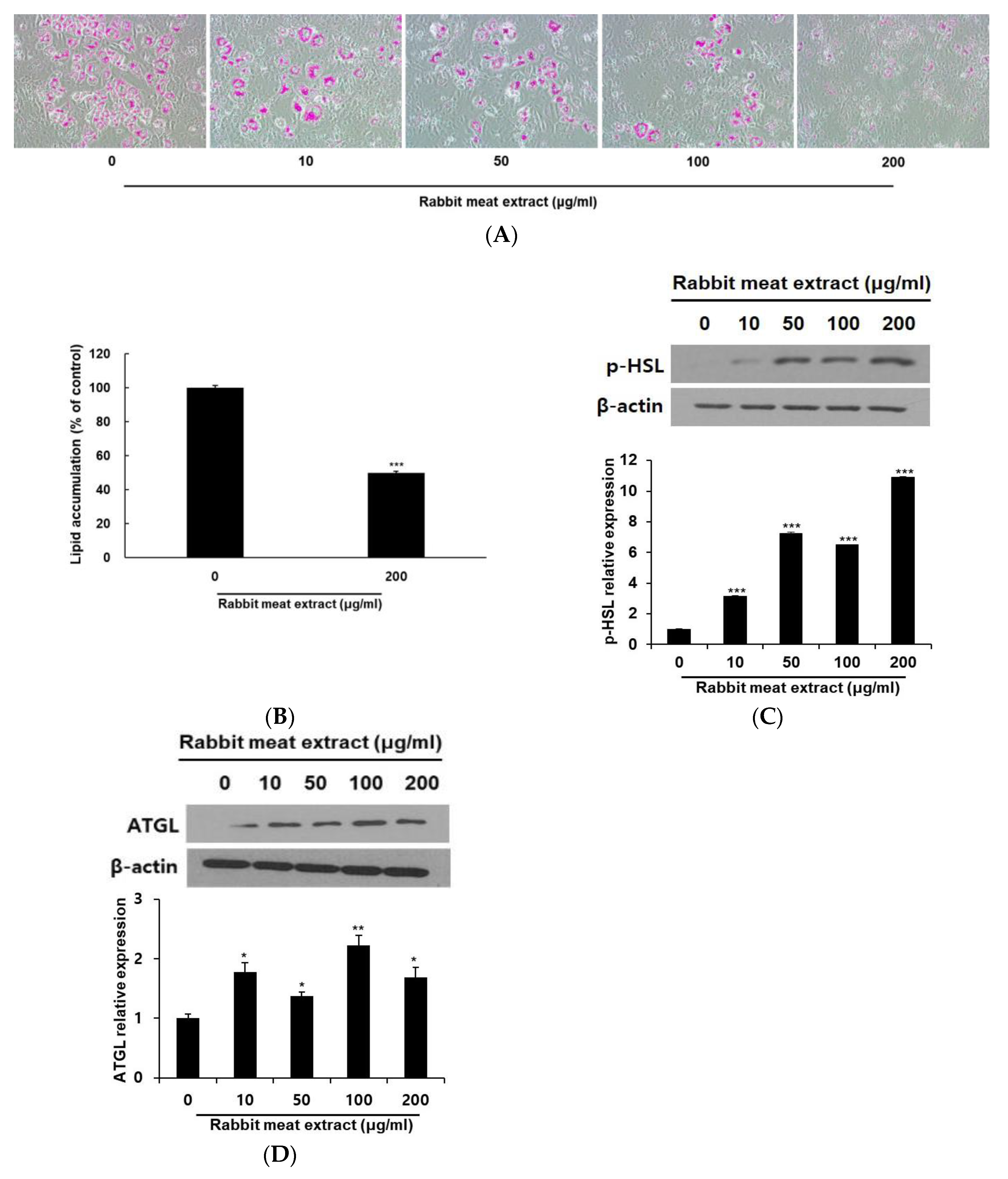
3.2. Rabbit Meat Extract Suppressed Intracellular Lipid Accumulation in Adipocytes
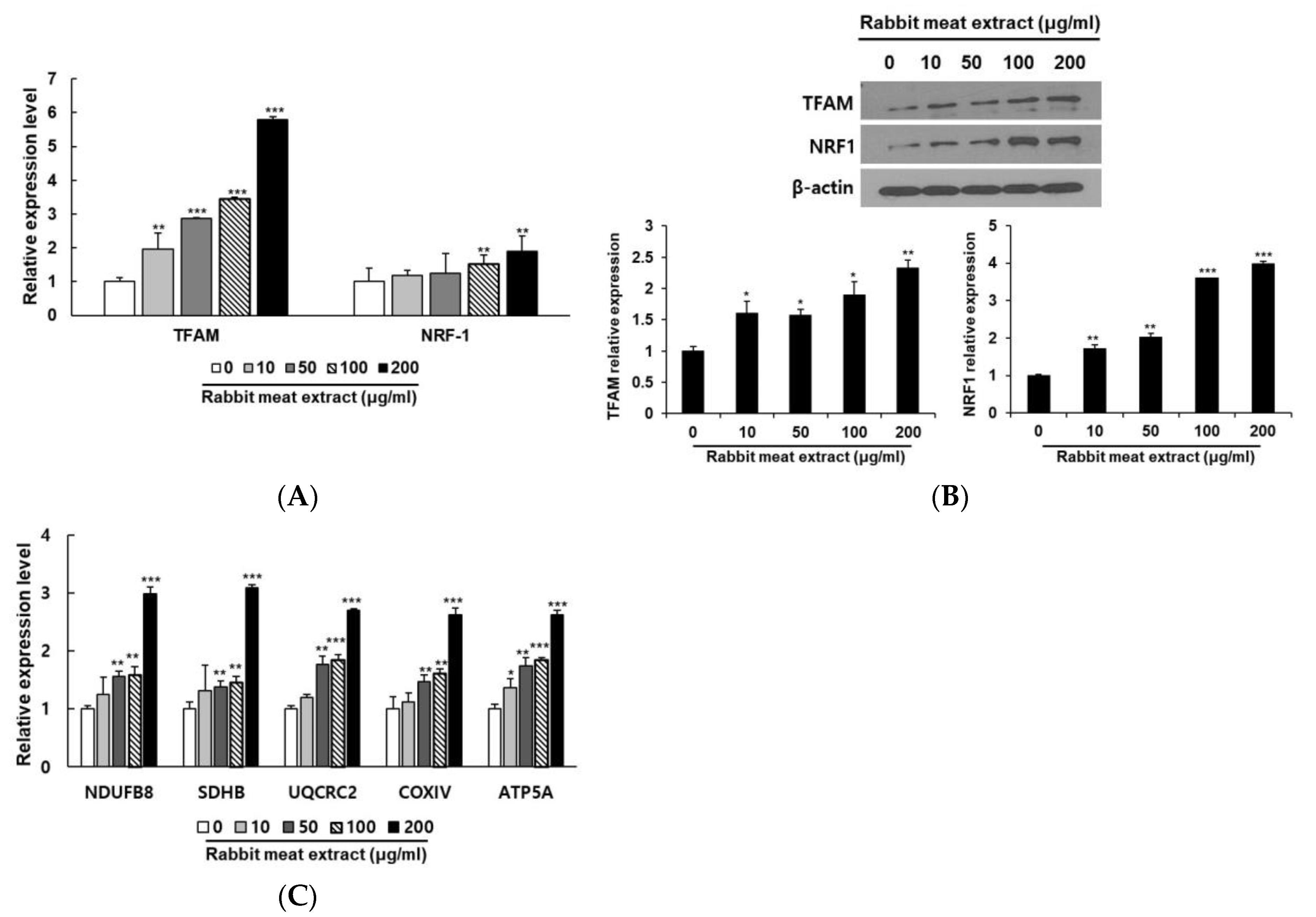
3.3. Rabbit Meat Extract Promoted Mitochondrial Biogenesis in 3T3−L1 Adipocytes
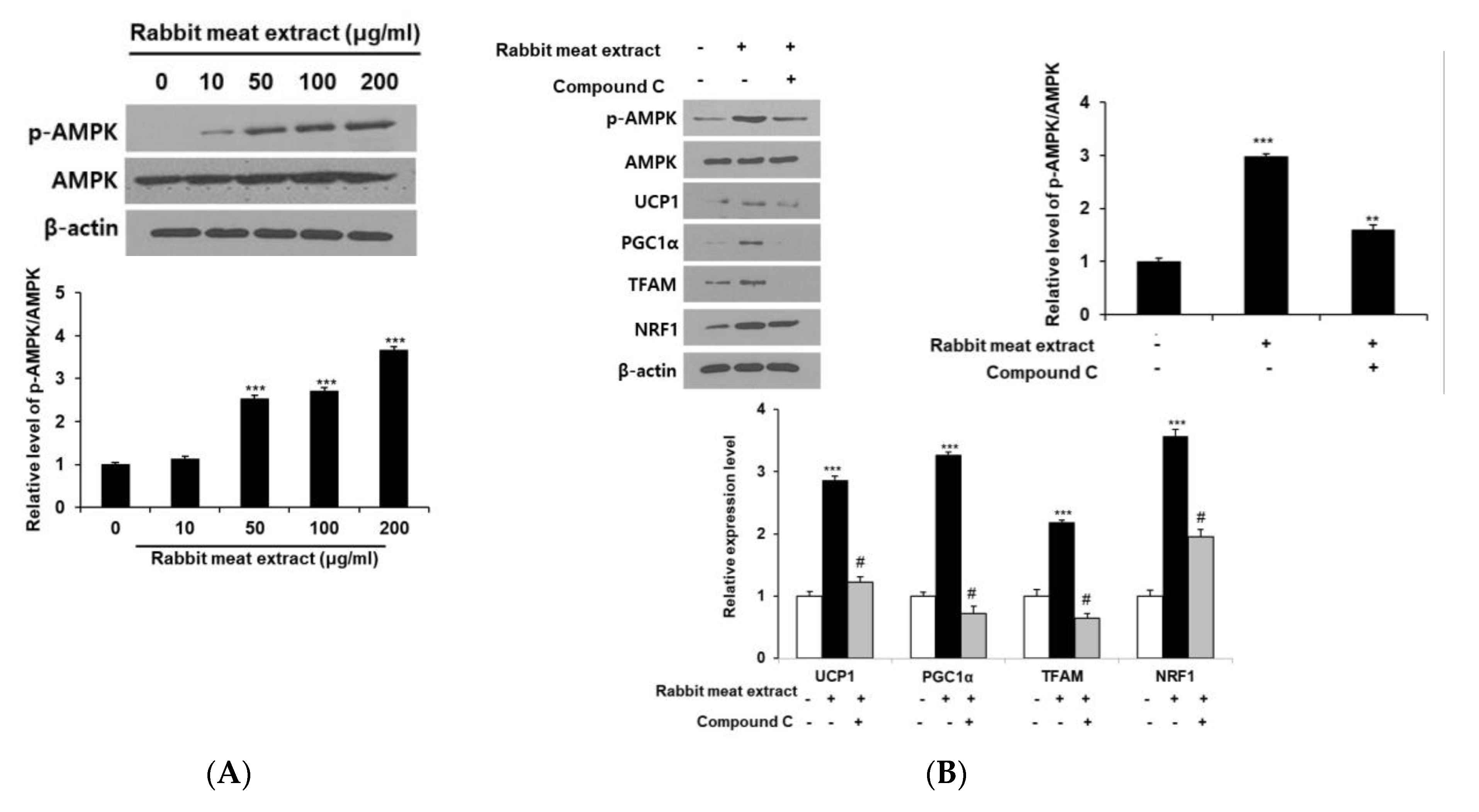
3.4. Effect of Rabbit Meat Extract on Browning of Adipocytes Is Mediated via AMPK Pathway
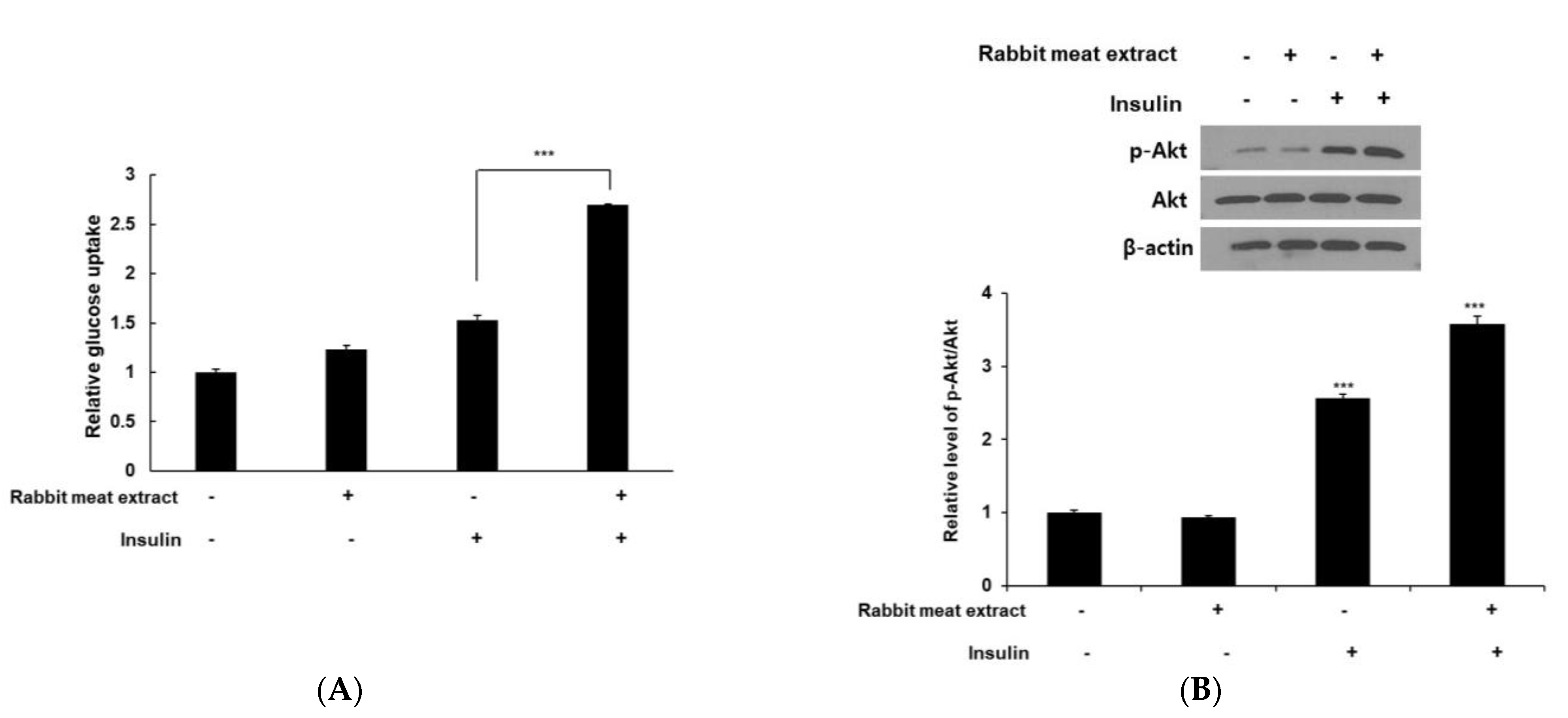
3.5. Rabbit Meat Extract Improved Insulin Resistance in 3T3−L1 Adipocytes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Wang, J.; Lin, Y.; Chen, J.; Liu, J.; Zhang, X. Nutritional activities of luteolin in obesity and associated metabolic disease: An eye on adipose tissues. Crit. Rev. Food Sci. Nutr. 2022, 1–15. [Google Scholar] [CrossRef]
- Shin, S. Regulation of adipose tissue biology by long−chain fatty acids: Metabolic effects and molecular mechanisms. J. Obes. Metab. Syndr. 2022, 31, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity−associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef]
- Shinde, A.B.; Song, A.; Wang, Q.A. Brown adipose tissue heterogeneity, energy metabolism, and beyond. Front. Endocrinol. 2021, 12, 651763. [Google Scholar] [CrossRef]
- Jung, S.M.; Sanchez−Gurmaches, J.; Guertin, D.A. Brown adipose tissue development and metabolism. Handb. Exp. Pharmacol. 2019, 251, 3–36. [Google Scholar] [PubMed]
- Maurer, S.; Harms, M.; Boucher, J. The colorful versatility of adipocytes: White−to−brown transdifferentiation and its therapeutic potential in humans. FEBS J. 2021, 288, 3628–3646. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Hu, W. Molecular and cellular regulation of thermogenic fat. Front. Endocrinol. 2023, 14, 1215772. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yao, J.; Lu, C.; Yang, N.; Han, X.; Lin, H.; Yin, Y. Cold−inducible PPA1 is critical for the adipocyte browning in mice. Biochem. Biophys. Res. Commun. 2023, 677, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Han, H.S.; Seong, J.K.; Ko, Y.G.; Koo, S.H. Involvement of a novel cAMP signaling mediator for beige adipogenesis. Metabolism 2023, 143, 155536. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, E.M.; Steinberg, G.R. Emerging role of AMPK in brown and beige adipose tissue (BAT): Implications for obesity, insulin resistance, and type 2 diabetes. Curr. Diab Rep. 2018, 18, 80. [Google Scholar] [CrossRef]
- Van der Vaart, J.I.; Boon, M.R.; Houtkooper, R.H. The role of AMPK signaling in brown adipose tissue activation. Cells 2021, 10, 1122. [Google Scholar] [CrossRef]
- Lee, H.Y.; Lee, G.H.; Kim, H.J.; Lim, Y.J.; Ko, B.M.; Kim, D.S.; Kim, T.W.; Kim, H.K.; Kim, T.Y.; Hwang, D.I.; et al. Combination of panax ginseng and Diospyrus kaki leaf inhibits white adipocyte differentiation and browning process through AMP−activated protein kinase (AMPK) activation in vitro and in vivo. Nutrients 2023, 15, 2276. [Google Scholar] [CrossRef]
- Yoon, Y.; Park, M.K.; Kim, K.H.; Lee, G.H. Black wheat extracts (arriheuk) regulate adipogenesis and lipolysis via adenosine monophosphate (AMP) activated protein kinase (AMPK)/sirtuin 1 (SIRT1) signaling pathways. Foods 2023, 12, 2727. [Google Scholar] [CrossRef] [PubMed]
- Forbes−Hernández, T.Y.; Cianciosi, D.; Ansary, J.; Mezzetti, B.; Bompadre, S.; Quiles, J.L.; Giampieri, F.; Battino, M. Strawberry (Fragaria x ananassa cv. Romina) methanolic extract promotes browning in 3T3−L1 cells. Food Funct. 2021, 11, 297–304. [Google Scholar] [CrossRef]
- Day, E.A.; Ford, R.J.; Steinberg, G.R. AMPK as a therapeutic target for treating metabolic diseases. Trends Endocrino Metab. 2017, 28, 545–560. [Google Scholar] [CrossRef]
- Lyons, C.L.; Roche, H.M. Nutritional modulation of AMPK−impact upon metabolic−inflammation. Int. J. Mol. Sci. 2018, 19, 3092. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Kim, S.; Lee, Y.; Oh, J.; Yoon, Y. Effects on goat meat extracts on α−glucosidase inhibitory activity, expression of Bcl−2−associated X (BAX), p53, and p21 in cell line and expression of atrogin−1, muscle atrophy F−box (MAFbx), muscle RING−finger protein−1 (MuRF−1) and myosin heavy chain 7 (MYH−7) in C2C12 myoblasts. Food Sci. Anim. Resour. 2023, 43, 359–373. [Google Scholar] [PubMed]
- Ni, L.; Zhuge, F.; Yang, S.; Ma, L.; Zheng, A.; Zhao, Y.; Hu, L.; Fu, Z.; Ni, Y. Hydrolyzed chicken meat extract attenuates neuroinflammation and cognitive impairment in middle−aged mouse by regulating M1/M2 microglial polarization. J. Agric. Food Chem. 2021, 69, 9800–9812. [Google Scholar] [CrossRef]
- Ni, Y.; Zhou, K.; Zhang, L.; Nan, S.; Fu, Z. Hydrolyzed chicken meat extract boosts the immunoregulatory effect by regulating M1/M2 macrophage polarization. J. Funct. Foods 2022, 95, 105194. [Google Scholar] [CrossRef]
- Tsai, S.F.; Chang, C.Y.; Yong, S.M.; Lim, A.L.; Nakao, Y.; Chen, S.J.; Kuo, Y.M. A hydrolyzed chicken extract CMI−168 enhances learning and memory in middle−aged mice. Nutrients 2018, 11, 27. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, D.; Lee, M.; Jang, A. Anti−inflammatory effect of dietary pork extract on proliferation and cytokine secretion using mouse primary splenocytes. Food Res. Int. 2017, 102, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Sawano, S.; Baba, K.; Sonoda, Y.; Wakamatsu, J.I.; Tomonaga, S.; Furuse, M.; Sato, Y.; Tatsumi, R.; Ikeuchi, Y.; Mizunoya, W.C. Beef extract supplementation promotes myoblast proliferation and myotube growth in C2C12 cells. Eur. J. Nutr. 2020, 59, 3735–3743. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Yamada, T. UCP1 dependent and independent thermogenesis in brown and beige adipocytes. Front. Endocrinol. 2020, 11, 498. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.A.; La Merrill, M.A. An emerging role for epigenetic regulation of PGC−1α expression in environmentally stimulated brown adipose thermogenesis. Environ. Epigenet. 2017, 3, dvx009. [Google Scholar] [CrossRef]
- Kim, S.H.; Plutzky, J. Brown fat and browning for the treatment of obesity and related metabolic disorders. Diabetes Metab. J. 2016, 40, 12–21. [Google Scholar] [CrossRef]
- Cheng, C.F.; Ku, H.C.; Lin, H. PGC−1α as a pivotal factor in lipid and metabolic regulation. Int. J. Mol. Sci. 2018, 19, 3447. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Kazak, L.; Spiegelman, B.M. New advances in adaptive thermogenesis: UCP1 and beyond. Cell Metab. 2019, 29, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, M.; Schreiber, R.; Haemmerle, G.; Lass, A.; Fledelius, C.; Jacobsen, P.; Tornqvist, H.; Zechner, R.; Zimmermann, R. Adipose triglyceride lipase and hormone−sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol. Chem. 2006, 281, 40236–40241. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.Y.; Choi, J.W.; Choi, H.J.; Im, S.W.; Park, G.H.; Jeong, J.B. Anti−obesity effects of Hovenia dulcis branches in mouse adipocytes, 3T3−L1 cells, and high−fat diet obese mice. Pharmazie 2023, 78, 89–92. [Google Scholar] [PubMed]
- Zhao, Y.; Qin, R. Vitamin D3 affects browning of white adipocytes by regulating autophagy via PI3K/Akt/mTOR/p53 signaling in vitro and in vivo. Apoptosis 2022, 27, 992–1003. [Google Scholar] [CrossRef]
- Vliora, M.; Grillo, E.; Corsini, M.; Ravelli, C.; Nintou, E.; Karligiotou, E.; Flouris, A.D.; Mitola, S. Irisin regulates thermogenesis and lipolysis in 3T3−L1 adipocytes. Biochim. Biophys. Acta Gen. Subj. 2022, 1866, 130085. [Google Scholar] [CrossRef] [PubMed]
- Matravadia, S.; Herbst, E.A.; Jain, S.S.; Mutch, D.M.; Holloway, G.P. Both linoleic acid α–linolenic acid prevent insulin resistance but have divergent impacts on skeletal muscle mitochondrial bioenergetics in obese Zucker rats. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E102–E114. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, W.; Chen, J.; Wang, X.; Wang, Y. AMP−activated protein kinase is required for the anti−adipogenic effects of alpha−linolenic acid. Nutr. Metab. 2015, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Bargut, T.C.L.; Martins, F.F.; Santos, L.P.; Aguila, M.B.; Mandarim−de−Lacerda, C.A. Administration of eicosapentaenoic and docosahexaenoic acids may improve the remodeling and browning in subcutaneous white adipose tissue and thermogenic markers in brown adipose tissue in mice. Mol. Cel. Endocrinol. 2019, 482, 18–27. [Google Scholar] [CrossRef]
- Pahlavani, M.; Razafimanjato, F.; Ramalingam, L.; Kalupahana, N.S.; Moussa, H.; Scoggin, S.; Moustaid−Moussa, N. Eicosapentaenoic acid regulates brown adipose tissue metabolism in high−fat−fed mice and in clonal brown adipocytes. J. Nutr. Biochem. 2017, 39, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Hu, M.; Huang, J.; Yu, S.; Li, X.; Li, Y.; Mao, L. Anti−obesity effects of DHA and EPA in high fat−induced insulin resistant mice. Food Funct. 2021, 12, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Major, G.C.; Chaput, J.P.; Ledoux, M.; St−Pierre, S.; Anderson, G.H.; Zemel, M.B.; Tremblay, A. Recent developments in calcium−related obesity research. Obes. Rev. 2008, 9, 428–445. [Google Scholar]
- Schrager, S. Dietary calcium intake and obesity. J. Am. Board. Fam. Pract. 2005, 18, 205–210. [Google Scholar] [CrossRef]
- Ayoub, J.J.; Samra, M.J.; Hlais, S.A.; Bassil, M.S.; Obeid, O.A. Effect of phosphorus supplementation on weight gain and waist circumference of overweight/obese adults: A randomized clinical trial. Nutr. Diabetes 2015, 5, e189. [Google Scholar] [CrossRef]
- Chu, N.; Chan, T.Y.; Chu, Y.K.; Ling, J.; He, J.; Leung, K.; Ma, R.C.W.; Chan, J.C.N.; Chow, E. Higher dietary magnesium and potassium intake are associated with lower body fat in people with impaired glucose tolerance. Front. Nutr. 2023, 10, 1169705. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, I.-S.; Lee, J.A.; Cho, S.-H.; Kim, H.-W.; Kim, Y.; Seo, K.; Cho, H.-W.; Lee, M.Y.; Chun, J.L.; Kim, K.H. Rabbit Meat Extract Induces Browning in 3T3−L1 Adipocytes via the AMP−Activated Protein Kinase Pathway. Foods 2023, 12, 3671. https://doi.org/10.3390/foods12193671
Bae I-S, Lee JA, Cho S-H, Kim H-W, Kim Y, Seo K, Cho H-W, Lee MY, Chun JL, Kim KH. Rabbit Meat Extract Induces Browning in 3T3−L1 Adipocytes via the AMP−Activated Protein Kinase Pathway. Foods. 2023; 12(19):3671. https://doi.org/10.3390/foods12193671
Chicago/Turabian StyleBae, In-Seon, Jeong Ah Lee, Soo-Hyun Cho, Hyoun-Wook Kim, Yunseok Kim, Kangmin Seo, Hyun-Woo Cho, Min Young Lee, Ju Lan Chun, and Ki Hyun Kim. 2023. "Rabbit Meat Extract Induces Browning in 3T3−L1 Adipocytes via the AMP−Activated Protein Kinase Pathway" Foods 12, no. 19: 3671. https://doi.org/10.3390/foods12193671
APA StyleBae, I. -S., Lee, J. A., Cho, S. -H., Kim, H. -W., Kim, Y., Seo, K., Cho, H. -W., Lee, M. Y., Chun, J. L., & Kim, K. H. (2023). Rabbit Meat Extract Induces Browning in 3T3−L1 Adipocytes via the AMP−Activated Protein Kinase Pathway. Foods, 12(19), 3671. https://doi.org/10.3390/foods12193671