
Genetic Diversity of Aspergillus flavus on Maize in Guatemala

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Aspergillus favus from Grain Samples
2.2. Population Genetic Analyses
3. Results and Discussion
3.1. Grain Samples and Phenotypes of Isolates
3.2. Genetic Analysis
4. Conclusions and Next Steps
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tay, K. Guatemala’s Corn Sectors Struggles with Contraband and Low Prices. USDA Foreign Agricultural Service, GT19007, 26 June 2019. Available online: https://agriexchange.apeda.gov.in/marketreport/Reports/Grain_and_Feed_Annual_Guatemala_City_Guatemala_6-26-2019.pdf (accessed on 16 October 2023).
- Ministerio de Agricultura, Ganadería y Alimentación—MAGA. Plan Operativo Anual 2020—Reprogramación; MAGA: Guatemala City, Guatemala, 2020. [Google Scholar]
- Tay, K. Grain and Animal Feed. USDA Foreign Agricultural Service, GT2022-0004, Guatemala, 15 April 2022. Available online: https://apps.fas.usda.gov/newgainapi/api/Report/DownloadReportByFileName?fileName=Grain%20and%20Feed%20Annual_Guatemala%20City_Guatemala_GT2022-0004.pdf (accessed on 16 October 2023).
- Tay, K. Grain and Feed Annual. USDA Foreign Agricultural Service, GT2023-0001, Guatemala, 6 April 2023. Available online: https://apps.fas.usda.gov/newgainapi/api/Report/DownloadReportByFileName?fileName=Grain%20and%20Feed%20Annual_Guatemala%20City_Guatemala_GT2023-0001.pdf (accessed on 16 October 2023).
- IARC. Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In International Agency for Research on Cancer: Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Press: Lyon, France, 1993; Available online: http://monographs.iarc.fr/ENG/Monographs/vol56/index.php (accessed on 16 October 2023).
- Torres, O.; Matute, J.; Gelineau-van Waes, J.; Maddox, J.; Gregory, S.G.; Ashley-Koch, A.E.; Showker, J.L.; Voss, K.A.; Riley, R.T. Human health implications from co-exposure to aflatoxins and fumonisins in maize-based foods in Latin America: Guatemala as a case study. World Mycotoxin J. 2015, 8, 143–159. [Google Scholar] [CrossRef]
- Alvarez, C.S.; Hernández, E.; Escobar, K.; Villagrán, C.I.; Kroker-Lobos, M.F.; Rivera-Andrade, A.; Smith, J.W.; Egner, P.A.; Lazo, M.; Freedman, N.D.; et al. Aflatoxin B1 exposure and liver cirrhosis in Guatemala: A case–control study. BMJ Open Gastroenterol. 2020, 7, e000380. [Google Scholar] [CrossRef] [PubMed]
- Voth-Gaeddert, L.E.; Torres, O.; Maldonado, J.; Krajmalnik-Brown, R.; Rittmann, B.E.; Oerther, D.B. Aflatoxin exposure, child stunting, and dysbiosis in the intestinal microbiome among children in Guatemala. Environ. Eng. Sci. 2019, 36, 958–968. [Google Scholar] [CrossRef]
- Groopman, J.D.; Smith, J.W.; Rivera-Andrade, A.; Alvarez, C.S.; Kroker-Lobos, M.F.; Egner, P.A.; Gharzouzi, E.; Dean, M.; McGlynn, K.A.; Ramírez-Zea, M. Aflatoxin and the aetiology of liver cancer and its implications for Guatemala. World Mycotoxin J. 2021, 14, 305–317. [Google Scholar] [CrossRef]
- Dorner, J.W. Biological control of aflatoxin contamination of crops. J. Toxicol. Toxin Rev. 2004, 23, 425–450. [Google Scholar] [CrossRef]
- Cotty, P.J.; Mellon, J.E. Ecology of aflatoxin producing fungi and biocontrol of aflatoxin contamination. Mycotoxin Res. 2006, 22, 110–117. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.L.; Adhikari, B.N.; Cotty, P.J. Biological control of aflatoxins in Africa: Current status and potential challenges in the face of climate change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef]
- Horn, B.W.; Dorner, J.W. Soil populations of Aspergillus species from section Flavi along a transect through peanut-growing regions of the United States. Mycologia 1998, 90, 767–776. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Weaver, M.A.; Horn, B.W.; Xie, W.; Shier, W.T. Comparison of cultural and analytical methods for determination of aflatoxin production by Mississippi Delta Aspergillus isolates. Can. J. Microbiol. 2004, 50, 193–199. [Google Scholar] [CrossRef]
- Weaver, M.A.; Abbas, H.K.; Brewer, M.J.; Pruter, L.S.; Little, N.S. Integration of biological control and transgenic insect protection for mitigation of mycotoxins in corn. Crop Prot. 2017, 98, 108–115. [Google Scholar] [CrossRef]
- Weaver, M.A.; Callicott, K.A.; Mehl, H.L.; Opoku, J.; Park, L.C.; Fields, K.S.; Mandel, J.R. Characterization of the Aspergillus flavus Population from Highly Aflatoxin-Contaminated Corn in the United States. Toxins 2022, 14, 755. [Google Scholar] [CrossRef] [PubMed]
- Sweany, R.R.; Damann, K.E., Jr.; Kaller, M.D. Comparison of soil and corn kernel Aspergillus flavus populations: Evidence for niche specialization. Phytopathology 2011, 101, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Callicott, K.A.; Kachapalula, P.; Edmunds, D.; Singh, P.; Jaime, R.; Islam, M.S.; Shenge, K.; Adhikari, B.N.; Arone-Maxwell, L.; Ching’anda, C.; et al. Brief Protocols for Research on Management of Aflatoxin-Producing Fungi, 2nd ed.; Phoenix, A.Z., Ed.; ARS: Beltsville, MD, USA; USDA: Washington, DC, USA, 2018. [Google Scholar]
- Grubisha, L.C.; Cotty, P.J. Twenty-four microsatellite markers for the aflatoxin-producing fungus Aspergillus flavus. Mol. Ecol. Resour. 2009, 9, 264–267. [Google Scholar] [CrossRef]
- Islam, M.S.; Callicott, K.A.; Mutegi, C.; Bandyopadhyay, R.; Cotty, P.J. Aspergillus flavus resident in Kenya: High genetic diversity in an ancient population primarily shaped by clonal reproduction and mutation-driven evolution. Fungal Ecol. 2018, 35, 20–33. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.H.; Moore, G.G.; Horn, B.W.; Carbone, I. Characterization and population analysis of the mating-type genes in Aspergillus flavus and Aspergillus parasiticus. Fungal Genet. Biol. 2008, 45, 1292–1299. [Google Scholar] [CrossRef]
- Meirmans, P.G.; Van Tienderen, P.H. GENOTYPE and GENODIVE: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, J. Optimum branchings. J. Res. Natl. Bur. Stand. B 1967, 71, 233–240. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef]
- Variety Release Announcement. Available online: https://cgspace.cgiar.org/bitstream/handle/10568/123590/ProjectInnovationSummary-Maize-P1870-I1967.pdf?sequence=1&isAllowed=y (accessed on 28 August 2023).
- Bentley, K.E.; Mauricio, R. High degree of clonal reproduction and lack of large-scale geographic patterning mark the introduced range of the invasive vine, kudzu (Pueraria montana var. lobata), in North America. Am. J. Bot. 2016, 103, 1499–1507. [Google Scholar] [CrossRef]
- Dorner, J.W.; Cole, R.J.; Wicklow, D.T. Aflatoxin reduction in corn through field application of competitive fungi. J. Food Prot. 1999, 62, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Weaver, M.A.; Abbas, H.K.; Falconer, L.L.; Allen, T.W.; Pringle, H.L., III; Sciumbato, G.L. Biological control of aflatoxin is effective and economical in Mississippi field trials. Crop Prot. 2015, 69, 52–55. [Google Scholar] [CrossRef]
- Molo, M.S.; White, J.B.; Cornish, V.; Gell, R.M.; Baars, O.; Singh, R.; Carbone, M.A.; Isakeit, T.; Wise, K.A.; Woloshuk, C.P.; et al. Asymmetrical lineage introgression and recombination in populations of Aspergillus flavus: Implications for biological control. PLoS ONE 2022, 17, e0276556. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
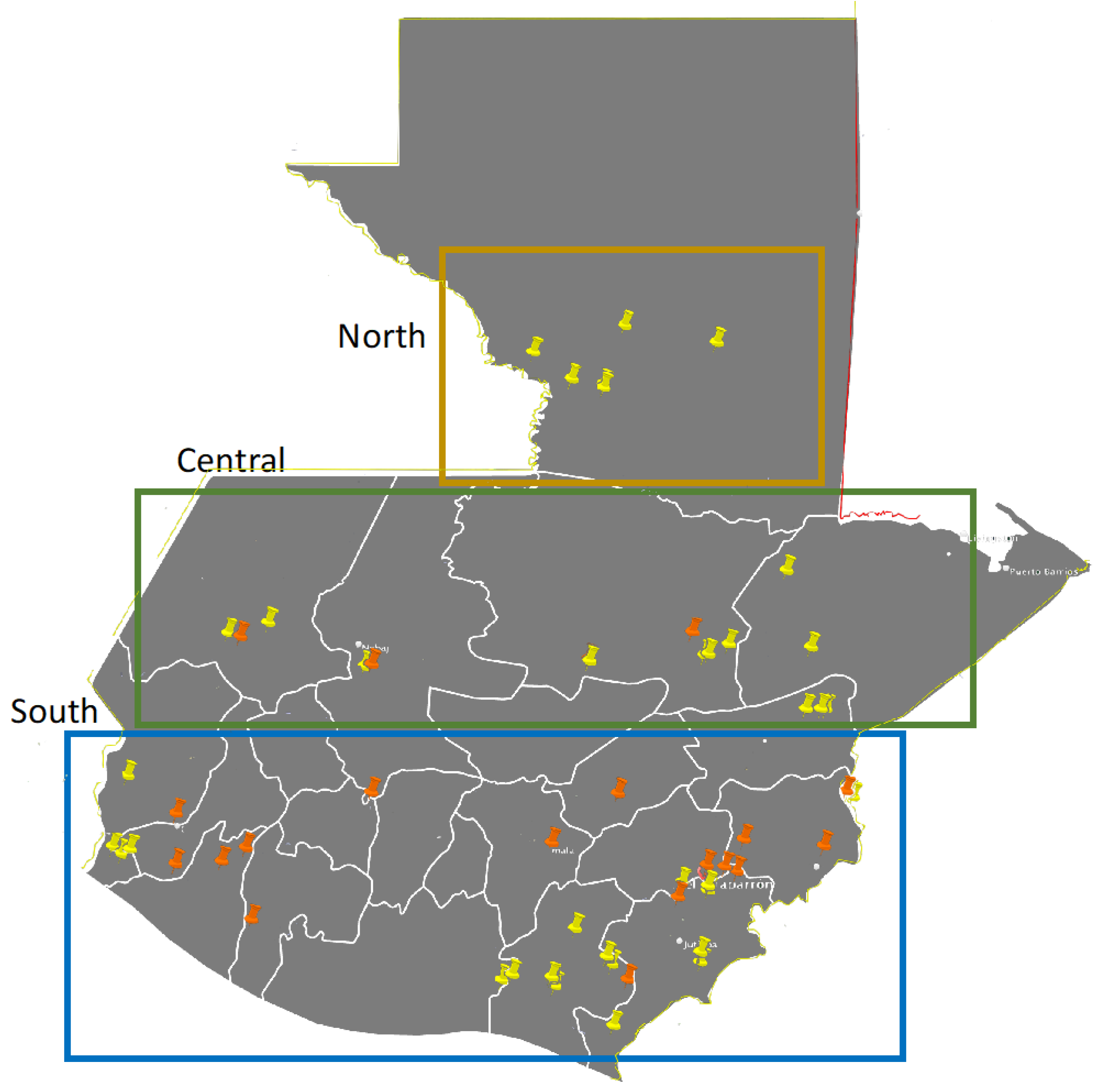
| 2021 Grain Samples | Cultivar | Elevation | Aldea | Municipality | Department |
| 1 | HB 83 | Midlands | Palos abrazodos | Moyuta | Jutiapa |
| 2 | JC-24 | Lowlands | Piedra Grande | Chiquimulilla | Santa Rosa |
| 3 | JC-24 | Lowlands | Piedra Grande | Chiquimulilla | Santa Rosa |
| 4 | Fortaleza F3 | Lowlands | Placetas | Chiquimulilla | Santa Rosa |
| 5 | JC-24 | Lowlands | Piedra Grande | Chiquimulilla | Santa Rosa |
| 6 | Unknown | Midlands | Obrajuelo | Agua Blanca | Jutiapa |
| 7 | Unknown | Midlands | Ipala | Chiquimula | |
| 8 | Fortaleza F3 | Midlands | Poza Verde | S. M. Chaparron | Jalapa |
| 9 | H5 SG | Midlands | Las Animas | S. M. Chaparron | Jalapa |
| 10 | Fortaleza F3 | Midlands | Jalapa | Jalapa | |
| 11 | Unknown | Midlands | Seamay | Senahu | Alta Verapaz |
| 12 | Unknown | Midlands | Paija | S.M. Tucuru | Alta Verapaz |
| 13 | Unknown | Midlands | Panhoma | Tamahu | Alta Verapaz |
| 14 | Unknown | Midlands | La herradura Chijul | S.M. Tucuru | Alta Verapaz |
| 15 | Unknown | Midlands | Peniel | Tacuru | Alta Verapaz |
| 16 | Fortaleza F3 | Highlands | Payushyec | Sacapulas | Quiche |
| 17 | Unknown | Highlands | Pueblo Viejo | San Sebastian | Huehuetenango |
| 18 | Unknown | Highlands | Piol | San Sebastian | Huehuetenango |
| 19 | Unknown | Highlands | Chumuxuquim | Sacapulas | Quiche |
| 20 | Unknown | Highlands | Payushyec | Sacapulas | Quiche |
| 21 | DK 410 | Lowlands | San Vicente 1 | Coatepeque | Quetzaltenango |
| 22 | DK 6018 | Lowlands | Caballo Blanco | Caballo Blanco | Retalhuleu |
| 23 | DK 390 | Lowlands | San Rafael | Coatepeque | Quetzaltenango |
| 24 | DK 6018 | Lowlands | Caballo Blanco | Caballo Blanco | Retalhuleu |
| 25 | Unknown | Lowlands | Los Palomos | Los Palomos | Retalhuleu |
| 26 | Fortaleza F3 | Midlands | Caulotes | Esquipulas | Chiquimula |
| 27 | Fortaleza F3 | Midlands | Caulotes | Esquipulas | Chiquimula |
| 28 | Fortaleza F3 | Lowlands | Linea C-8 | S. A. Villa Seca | Retalhuleu |
| 29 | Fortaleza F3 | Lowlands | Linea C-8 | S. A. Villa Seca | Retalhuleu |
| 30 | Fortaleza F3 | Midlands | Olopita | Esquipulas | Chiquimula |
| 31 | Fortaleza F3 | Midlands | Olopita | Esquipulas | Chiquimula |
| 32 | Fortaleza F3 | Lowlands | Linea C-18 | S. A. Villa Seca | Retalhuleu |
| 33 | Unknown | Lowlands | Linea A-3 | La maquina | Suchitepequez |
| 34 | Unknown | Midlands | El ovejero | Progreso | Jutiapa |
| 2023 Grain Samples | Cultivar | Elevation | Latitude | Longitude | |
| 1 | DK 390 | Lowlands | 14.05702 | −90.36782 | Santa Rosa |
| 2 | Unknown | Lowlands | 14.2781 | −90.28683 | Santa Rosa |
| 3 | HB 83 | Lowlands | 14.08254 | −90.37903 | Santa Rosa |
| 4 | DK 390 | Lowlands | 16.58299 | −90.09404 | Peten |
| 5 | HB 83 | Lowlands | 16.35779 | −90.17787 | Peten |
| 6 | Criollo | Midlands | 14.84926 | −92.06439 | San Marcos |
| 7 | Fortaleza F3 | Highlands | 15.27847 | −91.11817 | Quiche |
| 8 | Criollo | Midlands | 14.57462 | −92.12152 | San Marcos |
| 9 | Amarillo | Highlands | 15.44222 | −91.50228 | Huehuetenango |
| 10 | DK 390 | Midlands | 14.56577 | −92.05198 | San Marcos |
| 11 | DK 390 | Midlands | 14.5472 | −92.08604 | San Marcos |
| 12 | Criollo Blanco | Highlands | 15.40105 | −91.66328 | Huehuetenango |
| 13 | NK 102 | Lowlands | 16.48032 | −90.45605 | Peten |
| 14 | DK 390 | Lowlands | 16.58241 | −90.09541 | Peten |
| 15 | Criollo | Lowlands | 16.37866 | −90.30373 | Peten |
| 16 | Criollo | Lowlands | 16.34505 | −90.17684 | Peten |
| 17 | Fortaleza F5 | Lowlands | 14.07803 | −90.57511 | Santa Rosa |
| 18 | Fortaleza F3 | Lowlands | 14.09022 | −90.37938 | Santa Rosa |
| 19 | Fortaleza F5 | Midlands | 13.90407 | −90.13655 | Jutiapa |
| 20 | Fortaleza F5 | Lowlands | 14.09958 | −90.53562 | Santa Rosa |
| 21 | Unknown | Midlands | 15.32417 | −89.77944 | Alta Verapaz |
| 22 | Unknown | Midlands | 15.35361 | −89.35777 | Alta Verapaz |
| 23 | Unknown | Midlands | 15.32578 | −89.75277 | Alta Verapaz |
| 24 | Unknown | Midlands | 16.5175 | −89.72888 | Alta Verapaz |
| 25 | Unknown | Midlands | 15.36583 | −89.68333 | Alta Verapaz |
| 26 | Unknown | Midlands | 15.30222 | −90.23189 | Alta Verapaz |
| 27 | Unknown | Midlands | 15.32861 | −89.76222 | Alta Verapaz |
| 28 | DK 390 | ||||
| 29 | DK 390 | ||||
| 30 | HS 23 | ||||
| 31 | HB 83 | ||||
| 32 | Fortaleza F5 | ||||
| 33 | Fortaleza F5 | ||||
| 34 | HB 83 | Lowlands | 15.11846 | −89.31867 | Zacapa |
| 35 | HB 83 | Lowlands | 15.11846 | −89.37867 | Zacapa |
| 36 | Criollo | Midlands | 15.64639 | −89.45346 | Chiquimula |
| 37 | Criollo | Midlands | 14.44197 | −89.76379 | Jutiapa |
| 38 | Fortaleza F3 | Midlands | 14.4236 | −89.76408 | Jutiapa |
| 39 | Fortaleza F3 | Midlands | 14.79819 | −89.21239 | Chiquimula |
| 40 | Fortaleza F3 | Midlands | 14.78059 | −89.19426 | Chiquimula |
| 41 | DK 390 | Midlands | 14.16575 | −90.15913 | Jutiapa |
| 42 | DK 390 | Lowlands | 14.17018 | −90.1626 | Santa Rosa |
| 43 | H-5 | Midlands | 14.45488 | −89.86585 | Jalapa |
| 44 | H-5 | Midlands | 14.14853 | −89.79325 | Jutiapa |
| 45 | JC-24 | Lowlands | 14.16575 | −90.15913 | Santa Rosa |
| 46 | Fortaleza F3 | Midlands | 14.18684 | −89.79283 | Jutiapa |
| 47 | Fortaleza F3 | Lowlands | 14.14105 | −90.14312 | Santa Rosa |
| 48 | HB 83 | Lowlands | 15.119 | −89.3 | Zacapa |
| Aflatoxigenic Status of Isolates | N | Na | Ne | I | h | uh |
|---|---|---|---|---|---|---|
| Zero | 40 | 5.7 (0.57) a | 2.8 (0.46) a | 1.1 (0.14) a | 0.51 (0.06) a | 0.51 (0.06) a |
| Low | 20 | 2.8 (0.56) b | 2.1 (0.32) b | 0.61 (0.17) c | 0.33 (0.08) c | 0.34 (0.09) c |
| Medium | 46 | 5.1 (0.69) a | 2.5 (0.47) a | 0.90 (0.17) b | 0.42 (0.07) b | 0.43 (0.08) b |
| High | 20 | 2.7 (0.53) b | 2.1 (0.35) b | 0.61 (0.17) c | 0.32 (0.08) c | 0.34 (0.09) c |
| Clone only | 126 | 4.1 (0.33) | 2.3 (0.20) | 0.80 (0.08) | 0.39 (0.04) | 0.41 (0.04) |
| Total | 272 | 7.4 (0.74) | 2.3 (0.36) | 0.86 (0.15) | 0.40 (0.07) | 0.40 (0.07) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weaver, M.A.; Bowen, C.; Park, L.C.; Bastidas, A.; Drewry, S.G.; Mandel, J.R. Genetic Diversity of Aspergillus flavus on Maize in Guatemala. Foods 2023, 12, 3864. https://doi.org/10.3390/foods12203864
Weaver MA, Bowen C, Park LC, Bastidas A, Drewry SG, Mandel JR. Genetic Diversity of Aspergillus flavus on Maize in Guatemala. Foods. 2023; 12(20):3864. https://doi.org/10.3390/foods12203864
Chicago/Turabian StyleWeaver, Mark A., Curt Bowen, Lilly C. Park, Angela Bastidas, Samantha G. Drewry, and Jennifer R. Mandel. 2023. "Genetic Diversity of Aspergillus flavus on Maize in Guatemala" Foods 12, no. 20: 3864. https://doi.org/10.3390/foods12203864
APA StyleWeaver, M. A., Bowen, C., Park, L. C., Bastidas, A., Drewry, S. G., & Mandel, J. R. (2023). Genetic Diversity of Aspergillus flavus on Maize in Guatemala. Foods, 12(20), 3864. https://doi.org/10.3390/foods12203864