





Screening and Activity Analysis of α-Glucosidase Inhibitory Peptides Derived from Coix Seed Prolamins Using Bioinformatics and Molecular Docking


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Coix Seed Prolamins
2.3. Preparation of Coix Seed Prolamins Hydrolysate (CHPs)
2.4. Degree of Hydrolysis (DH) Assay
2.5. α-Glucosidase Inhibitory Activity Assay
2.6. Identification of the Peptides (LC-MS/MS)
2.7. Bioinformatics Analysis of Bioactive Peptides (BPs) Screening from CHPs
2.8. Molecular Docking
2.9. Thermal Effects and pH Effects on the α-Glucosidase Inhibitory Activities of CHPs
2.10. Effects of Food Ingredients on the α-Glucosidase Inhibitory Activities of CHPs
2.11. Data Analysis
3. Results
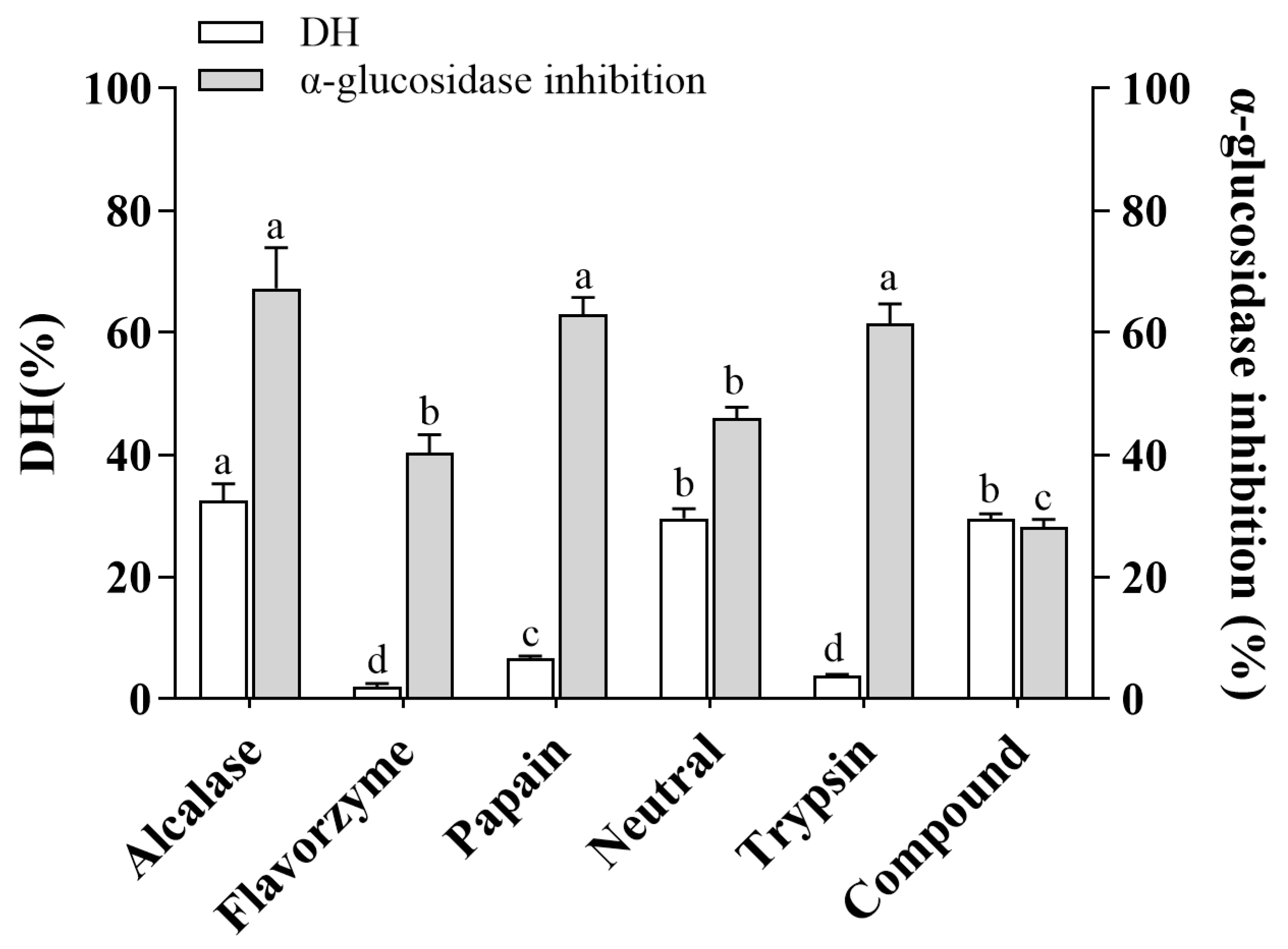
3.1. The DH and α-Glucosidase Inhibitory Activity of CHPs Generated by Different Proteases
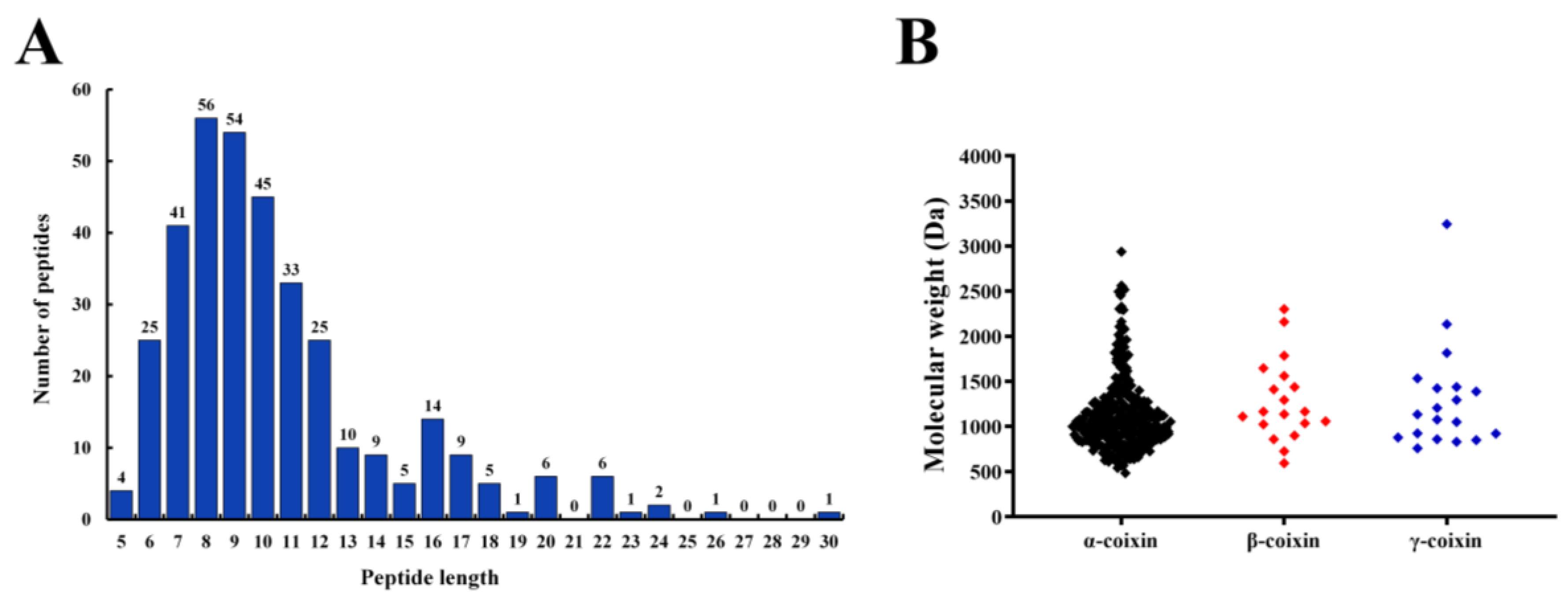
3.2. Identification of Peptides from CHPs-Alc
3.3. Screening of α-Glucosidase Inhibitory Peptides from CHPs-Alc via Bioinformatics
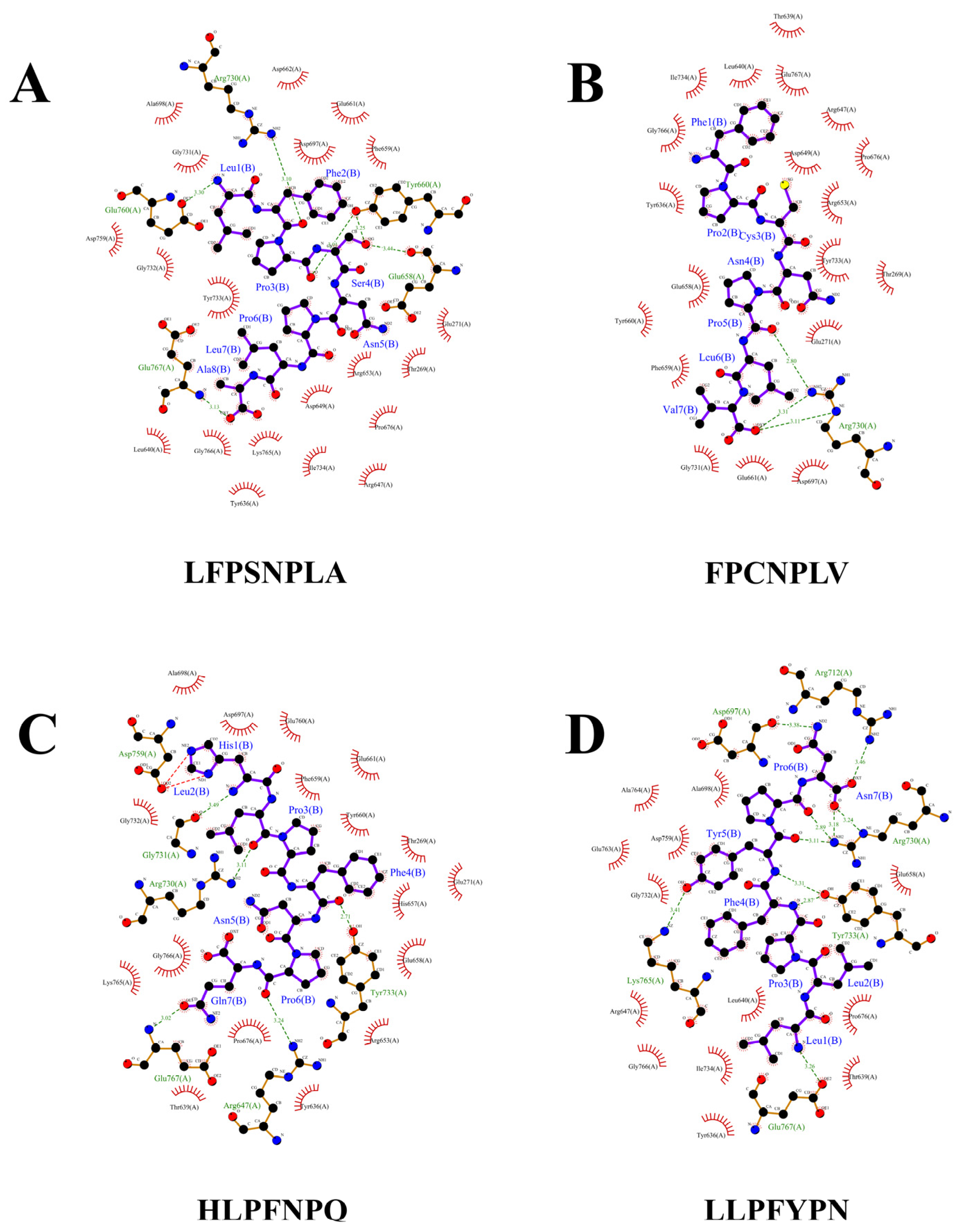
3.4. The Interaction Mechanism between α-Glucosidase Inhibitory Peptides and α-Glucosidase
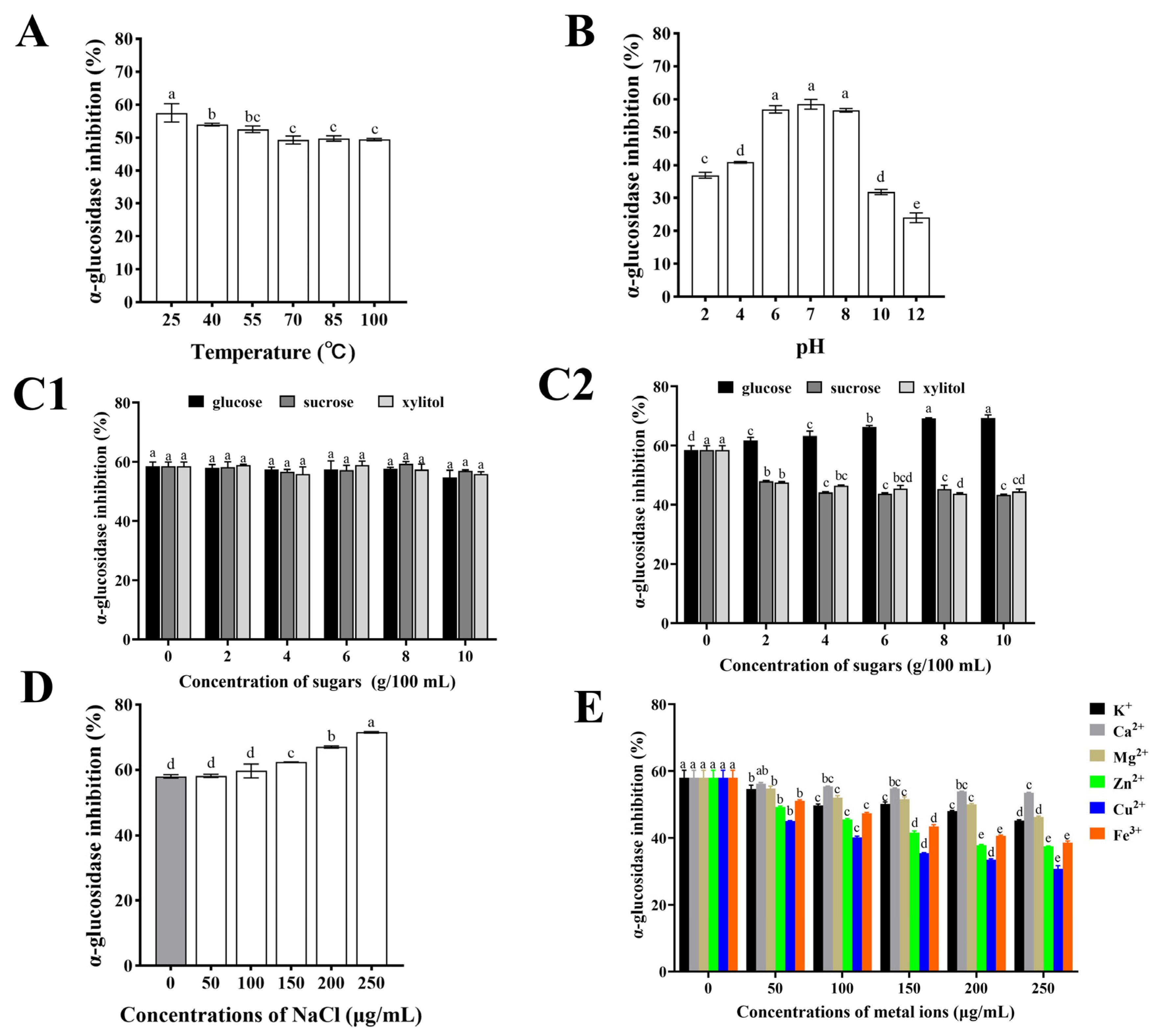
3.5. Effect of Food Processing on α-Glucosidase Activity of CHPs-Alc
3.5.1. Temperature and pH
3.5.2. Edible Sugar
3.5.3. NaCl
3.5.4. Metal Ions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lovic, D.; Piperidou, A.; Zografou, I.; Grassos, H.; Pittaras, A.; Manolis, A. The growing epidemic of diabetes mellitus. Curr. Vasc. Pharmacol. 2020, 18, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, T.; Fang, L.; Liu, C.; Liu, X.; Li, H.; Shi, J.; Min, W. Anti-diabetic effect by walnut (Juglans mandshurica Maxim.)-derived peptide LPLLR through inhibiting α-glucosidase and α-amylase, and alleviating insulin resistance of hepatic HepG2 cells. J. Funct. Foods 2020, 69, 103944. [Google Scholar] [CrossRef]
- Mushtaq, M.; Gani, A.; Noor, N.; Masoodi, F.A. Phenotypic and probiotic characterization of isolated LAB from Himalayan cheese (Kradi/Kalari) and effect of simulated gastrointestinal digestion on its bioactivity. LWT 2021, 149, 111669. [Google Scholar] [CrossRef]
- Mora, L.; González-Rogel, D.; Heres, A.; Toldrá, F. Iberian dry-cured ham as a potential source of α-glucosidase-inhibitory peptides. J. Funct. Foods 2020, 67, 103840. [Google Scholar] [CrossRef]
- Igbokwe, C.J.; Wei, M.; Feng, Y.; Duan, Y.; Ma, H.; Zhang, H. Coix seed: A review of its physicochemical composition, bioactivity, processing, application, functionality, and safety aspects. Food Rev. Int. 2022, 38, 921–939. [Google Scholar] [CrossRef]
- Meng, L.N. Adlay Seed Protein Dependent on IKK/NF-κB Pathway to Control Inflammation and Improve Insulin Resistance in Type 2 Diabetes Mellitus; Hefei University of Technology: Hefei, China, 2018. [Google Scholar]
- Watanabe, M.; Kato, M.; Ayugase, J. Anti-diabetic effects of adlay protein in type 2 diabetic db/db mice. Food Sci. Technol. Res. 2012, 18, 383–390. [Google Scholar] [CrossRef]
- Ulug, S.K.; Jahandideh, F.; Wu, J. Novel technologies for the production of bioactive peptides. Trends Food Sci. Technol. 2021, 108, 27–39. [Google Scholar] [CrossRef]
- Wang, L.; Yuan, J.; Zhang, X.; Qiao, Y. Protein composition analysis of Coix lacryma-jobi L. var. ma-yuen (Roman.) Stapf seeds. Prog. Mod. Biomed. 2012, 12, 4416–4432. [Google Scholar]
- Chen, P.; Li, L.; Huo, X.; Qiao, L.; Zhang, Y.; Chen, Z.; Wang, L. New angiotensin-converting enzyme inhibitory peptide from Coix prolamin and its influence on the gene expression of renin-angiotensin system in vein endothelial cells. J. Cereal Sci. 2020, 96, 103099. [Google Scholar] [CrossRef]
- Chen, L.C.; Zhang, S.Y.; Zi, Y.; Zhao, H.M.; Wang, H.Y.; Zhang, Y. Functional coix seed protein hydrolysates as a novel agent with potential hepatoprotective effect. Food Funct. 2020, 11, 9495–9502. [Google Scholar] [CrossRef]
- Li, Z.M.; Zhang, S.; Bai, L.; Tang, H.C.; Zhang, G.F.; Zhang, J.Y.; Meng, W.M.; Zhang, D.J. Flexible processing technology of coix seed prolamins by combined heat-ultrasound: Effects on their enzymatic hydrolysis characteristics and the hypoglycemic activities of derived peptides. Ultrason. Sonochem. 2023, 98, 106526. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Z.M.; Feng, Y.C.; Wang, C.Y.; Zhang, D.J. Processing Enhances Coix Seed Prolamins Structure and Releases Functional Peptides after Digestion: In Silico and In Vitro Studies. Foods 2023, 12, 2500. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.J.; Wang, Z.W. Structure characteristics of Coix seeds prolamins and physicochemical and mechanical properties of their films. J. Cereal Sci. 2018, 79, 233–239. [Google Scholar] [CrossRef]
- Mojica, L.; De Mejía, E.G. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Ma, Y.L.; Sun, P.; Thakur, K.; Wang, S.; Zhang, J.G.; Wei, Z.J. Purification and characterisation of α-glucosidase inhibitory peptides from defatted camellia seed cake. Int. J. Food Sci. Technol. 2021, 56, 138–147. [Google Scholar] [CrossRef]
- Huang, Q.Q.; Tian, Y.P. α-Glucosidase and dipeptidyl peptidase IV inhibitory activity and peptide composition of Porphyra yezoensis protein hydrolysate. Food Sci. 2020, 41, 110–116. [Google Scholar] [CrossRef]
- Singh, P.P.; Gupta, V.; Prakash, B. Recent advancement in functional properties and toxicity assessment of plant-derived bioactive peptides using bioinformatic approaches. Crit. Rev. Food Sci. Nutr. 2023, 63, 4503–4521. [Google Scholar] [CrossRef]
- Liu, W.W.; Li, H.Y.; Wen, Y.T.; Liu, Y.L.; Wang, J.; Sun, B.G. Molecular mechanism for the α-glucosidase inhibitory effect of wheat germ peptides. J. Agric. Food Chem. 2021, 69, 15231–15239. [Google Scholar] [CrossRef]
- Wong, F.C.; Xiao, J.; Michelle, G.L.O.; Pang, M.J.; Wong, S.J.; The, L.K.; Chai, T.T. Identification and characterization of antioxidant peptides from hydrolysate of blue-spotted stingray and their stability against thermal, pH and simulated gastrointestinal digestion treatments. Food Chem. 2019, 271, 614–622. [Google Scholar] [CrossRef]
- Nourmohammadi, E.; SadeghiMahoonak, A.; Alami, M.; Ghorbani, M. Amino acid composition and antioxidative properties of hydrolysed pumpkin (Cucurbita pepo L.) oil cake protein. Int. J. Food Prop. 2017, 20, 3244–3255. [Google Scholar] [CrossRef]
- Mokni Ghribi, A.; Maklouf Gafsi, I.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Yang, R.; Zhao, W. Effect of acid deamidation-alcalase hydrolysis induced modification on functional and bitter-masking properties of wheat gluten hydrolysates. Food Chem. 2019, 277, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Du, M.; Shen, M.; Wu, T.; Lin, L. Physico-chemical properties, antioxidant activities and angiotensin-I converting enzyme inhibitory of protein hydrolysates from Mung bean (Vigna radiate). Food Chem. 2019, 270, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.C.; Zhao, H.X.; Pan, X.X.; Orfila, C.; Lu, W.H.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef]
- Qiu, L.Y.; Deng, Z.Y.; Zhao, C.D.; Xiao, T.; Weng, C.; Li, J.; Zheng, L.F. Nutritional composition and proteomic analysis of soft-shelled turtle (Pelodiscus sinensis) egg and identification of oligopeptides with alpha-glucosidase inhibitory activity. Food Res. Int. 2021, 145, 110414. [Google Scholar] [CrossRef]
- Beverly, R.L.; Huston, R.K.; Markell, A.M.; McCulley, E.A.; Martin, R.L.; Dallas, D.C. Milk peptides survive in vivo gastrointestinal digestion and are excreted in the stool of infants. J. Nutr. 2020, 150, 712–721. [Google Scholar] [CrossRef]
- Xu, Q.B.; Hong, H.; Wu, J.P.; Yan, X.H. Bioavailability of bioactive peptides derived from food proteins across the intestinal epithelial membrane: A review. Trends Food Sci. Technol. 2019, 86, 399–411. [Google Scholar] [CrossRef]
- González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, α-amylase, and α-glucosidase enzymes. Int. J. Mol. Sci. 2018, 19, 2883. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef]
- Mudgil, P.; Kilari, B.P.; Kamal, H.; Olalere, O.A.; FitzGerald, R.J.; Gan, C.Y.; Maqsood, S. Multifunctional bioactive peptides derived from quinoa protein hydrolysates: Inhibition of α-glucosidase, dipeptidyl peptidase-IV and angiotensin I converting enzymes. J. Cereal Sci. 2020, 96, 103130. [Google Scholar] [CrossRef]
- Di Stefano, E.; Oliviero, T.; Udenigwe, C.C. Functional significance and structure–activity relationship of food-derived α-glucosidase inhibitors. Curr. Opin. Food Sci. 2018, 20, 7–12. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, Y.Y.; Yu, T.T.; He, J.T.; Cui, J.; Wang, J.N.; Cheng, X.N.; Fan, J. Oat globulin peptides regulate antidiabetic drug targets and glucose transporters in Caco-2 cells. J. Funct. Foods. 2018, 42, 12–20. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef]
- Sadri, H.; Larki, N.N.; Kolahian, S. Hypoglycemic and hypolipidemic effects of leucine, zinc, and chromium, alone and in combination, in rats with type 2 diabetes. Biol. Trace Elem. Res. 2017, 180, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2018, 91, 370–379. [Google Scholar] [CrossRef]
- Li, Z.M.; Zhang, S.; Meng, W.H.; Zhang, J.Y.; Zhang, D.J. Food-Derived α-Glucosidase Inhibitory Peptides: Research Progress on Structure-Activity Relationship, Safety and Bioavailability. Food Sci. 2023, 44, 298–309. [Google Scholar] [CrossRef]
- Mojica, L.; Luna-Vital, D.A.; González de Mejía, E. Characterization of peptides from common bean protein isolates and their potential to inhibit markers of type-2 diabetes, hypertension and oxidative stress. J. Sci. Food Agric. 2017, 97, 2401–2410. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Wu, F.H.; He, Z.P.; Fang, X.Z.; Liu, X.Q. Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis. Foods 2023, 12, 393. [Google Scholar] [CrossRef]
- Kamdem, J.P.; Tsopmo, A. Reactivity of peptides within the food matrix. J. Food Biochem. 2019, 43, e12489. [Google Scholar] [CrossRef]
- Chai, T.T.; Xiao, J.B.; Dass, S.M.; Teoh, J.Y.; Ee, K.Y.; Ng, W.J.; Wong, F.C. Identification of antioxidant peptides derived from tropical jackfruit seed and investigation of the stability profiles. Food Chem. 2021, 340, 127876. [Google Scholar] [CrossRef]
- Liu, L.; Li, X.D.; Du, L.L.; Zhang, X.X.; Yang, W.S.; Zhang, H.D. Effect of ultrasound assisted heating on structure and antioxidant activity of whey protein peptide grafted with galactose. LWT 2019, 109, 130–136. [Google Scholar] [CrossRef]
- Enciso, M.; Schütte, C.; Delle Site, L. Influence of pH and sequence in peptide aggregation via molecular simulation. J. Chem. Phys. 2015, 143, 243130. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Mirdamadi, S.; Safavi, M.; Soleymanzadeh, N. The stability of antioxidant and ACE-inhibitory peptides as influenced by peptide sequences. LWT 2020, 130, 109710. [Google Scholar] [CrossRef]
- Wang, X.Q.; Yu, H.H.; Xing, R.G.; Chen, X.L.; Liu, S.; Li, P.C. Optimization of the extraction and stability of antioxidative peptides from mackerel (Pneumatophorus japonicus) protein. BioMed Res. Int. 2017, 2017, 6837285. [Google Scholar] [CrossRef]
- Pereira, A.M.; Lisboa, C.R.; Santos, T.D.; Costa, J.A.V. Bioactive stability of microalgal protein hydrolysates under food processing and storage conditions. J. Food Sci. Technol. 2019, 56, 4543–4551. [Google Scholar] [CrossRef]
- Hmidet, N.; Balti, R.; Nasri, R.; Sila, A.; Bougatef, A.; Nasri, M. Improvement of functional properties and antioxidant activities of cuttlefish (Sepia officinalis) muscle proteins hydrolyzed by Bacillus mojavensis A21 proteases. Food Res. Int. 2011, 44, 2703–2711. [Google Scholar] [CrossRef]
- Liu, L.; Li, S.S.; Zheng, J.X.; Bu, T.T.; He, G.Q.; Wu, J.P. Safety considerations on food protein-derived bioactive peptides. Trends Food Sci. Technol. 2020, 96, 199–207. [Google Scholar] [CrossRef]
- Shen, Q.; Ou, A.; Liu, S.; Elango, J.; Wang, S.J.; Henriques da Silva, T.; Wu, W.; Robinson, J.; Bao, B. Effects of ion concentrations on the hydroxyl radical scavenging rate and reducing power of fish collagen peptides. J. Food Biochem. 2019, 43, e12789. [Google Scholar] [CrossRef]
- Zhu, C.Z.; Zhang, W.G.; Kang, Z.L.; Zhou, G.H.; Xu, X.L. Stability of an antioxidant peptide extracted from Jinhua ham. Meat Sci. 2014, 96, 783–789. [Google Scholar] [CrossRef]
- Zhang, Y.; He, S.D.; Bonneil, É.; Simpson, B.K. Generation of antioxidative peptides from Atlantic sea cucumber using alcalase versus trypsin: In vitro activity, de novo sequencing, and in silico docking for in vivo function prediction. Food Chem. 2020, 306, 125581. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Shi, P.Q.; Li, Y.; Zhuang, Y.L.; You, L.Z.; Liu, L.; Wang, W. A novel ACE-inhibitory hexapeptide from camellia glutelin-2 hydrolysates: Identification, characterization and stability profiles under different food processing conditions. LWT 2021, 147, 111682. [Google Scholar] [CrossRef]
- Sahni, P.; Sharma, S.; Surasani, V.K.R. Influence of processing and pH on amino acid profile, morphology, electrophoretic pattern, bioactive potential and functional characteristics of alfalfa protein isolates. Food Chem. 2020, 333, 127503. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Wang, S.K.; Zhu, X.; Li, Q.Q.; Fan, Y.; Cheng, D.; Li, B.F. A novel calcium-binding peptide from Antarctic krill protein hydrolysates and identification of binding sites of calcium-peptide complex. Food Chem. 2018, 243, 389–395. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent | Blank Group/μL | Blank Control Group/μL | Sample Group/μL | Sample Control Group/μL |
|---|---|---|---|---|
| The solution of CHPs | / | / | 50 | 50 |
| α-glucosidase (0.1 U/mL) | 50 | / | 50 | / |
| Phosphate Buffered Saline (PBS) (0.1 mol/L, pH 6.8) | 50 | 100 | / | 50 |
| Mix well and incubate at 37 °C for 10 min | ||||
| pNPG (2.5 mmol/L) | 50 | 50 | 50 | 50 |
| Mix well and incubate at 37 °C for 30 min | ||||
| Na2CO3 (0.5 mol/L) | 100 | 100 | 100 | 100 |
| Absorbance was measured at 405 nm | ||||
| NO. | Sequence | Length | MW/Da | Peptide Ranker Score | Affinity (kcal/mol) | pI | Net Charge at pH7.0 | Hydrophobicity /(Kcal/mol) | Toxicy |
|---|---|---|---|---|---|---|---|---|---|
| 1 | LFPSNPLA | 8 | 857.4647 | 0.648003 | −8.9 | 5.6 | 0 | 5.78 | - |
| 2 | FPCNPLV | 7 | 845.41056 | 0.852117 | −8.8 | 5.24 | 0 | 5.59 | - |
| 3 | HLPFNPQ | 7 | 851.42899 | 0.639342 | −8.7 | 7.69 | 0 | 9.17 | - |
| 4 | LLPFYPN | 7 | 862.45889 | 0.81539 | −8.7 | 5.39 | 0 | 4.11 | - |
| 5 | FLSPF | 5 | 609.31625 | 0.964433 | −8.5 | 5.41 | 0 | 3.83 | - |
| 6 | FPCNPLVA | 8 | 916.44767 | 0.745656 | −8.4 | 5.21 | 0 | 6.09 | - |
| 7 | LPPFLPS | 7 | 769.43743 | 0.850373 | −8.4 | 5.45 | 0 | 4.57 | - |
| 8 | QLFPSNPLA | 9 | 985.52328 | 0.585702 | −8.4 | 5.49 | 0 | 6.55 | - |
| 9 | QQHLPFNPQ | 9 | 1107.5461 | 0.574762 | −8.4 | 7.59 | 0 | 10.71 | - |
| 10 | SWQQPIVGR | 9 | 1069.5669 | 0.703379 | −8.4 | 10.85 | +1 | 9.33 | - |
| NO. | Sequence | Binding Energy/kcal mol−1 | Residues Formed Hydrogen Bonds with the Ligand | Number of Hydrogen Bonds | Distance of Hydrogen Bond | Number of Hydrophobic Amino Acid Residues |
|---|---|---|---|---|---|---|
| 1 | LFPSNPLA | −8.9 | Glu658, Tyr660, Arg730, Glu760, Glu767 | 5 | 3.44Å, 3.25Å, 3.10Å, 3.30Å, 3.13Å | 19 |
| 2 | FPCNPLV | −8.8 | Arg730 | 3 | 3.11Å, 3.31Å, 2.80Å | 19 |
| 3 | HLPFNPQ | −8.7 | Arg647, Tyr733, Gly731, Arg730, Glu767 | 5 | 3.24Å, 2.71Å, 3.49Å, 3.11Å, 3.02Å | 17 |
| 4 | LLPFYPN | −8.7 | Glu767, Tyr733, Arg730, Arg712, Asp697, Lys765 | 10 | 3.26Å, 2.87Å, 3.31Å, 3.11Å, 2.89Å, 3.18Å, 3.24Å, 3.46Å, 3.38Å, 3.41Å | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhang, S.; Meng, W.; Zhang, J.; Zhang, D. Screening and Activity Analysis of α-Glucosidase Inhibitory Peptides Derived from Coix Seed Prolamins Using Bioinformatics and Molecular Docking. Foods 2023, 12, 3970. https://doi.org/10.3390/foods12213970
Li Z, Zhang S, Meng W, Zhang J, Zhang D. Screening and Activity Analysis of α-Glucosidase Inhibitory Peptides Derived from Coix Seed Prolamins Using Bioinformatics and Molecular Docking. Foods. 2023; 12(21):3970. https://doi.org/10.3390/foods12213970
Chicago/Turabian StyleLi, Zhiming, Shu Zhang, Weihong Meng, Jiayu Zhang, and Dongjie Zhang. 2023. "Screening and Activity Analysis of α-Glucosidase Inhibitory Peptides Derived from Coix Seed Prolamins Using Bioinformatics and Molecular Docking" Foods 12, no. 21: 3970. https://doi.org/10.3390/foods12213970
APA StyleLi, Z., Zhang, S., Meng, W., Zhang, J., & Zhang, D. (2023). Screening and Activity Analysis of α-Glucosidase Inhibitory Peptides Derived from Coix Seed Prolamins Using Bioinformatics and Molecular Docking. Foods, 12(21), 3970. https://doi.org/10.3390/foods12213970