Enhancement of Rabbit Meat Functionality by Replacing Traditional Feed Raw Materials with Alternative and More Sustainable Freshwater Cladophora glomerata Macroalgal Biomass in Their Diets





Abstract
:1. Introduction
2. Materials and Methods
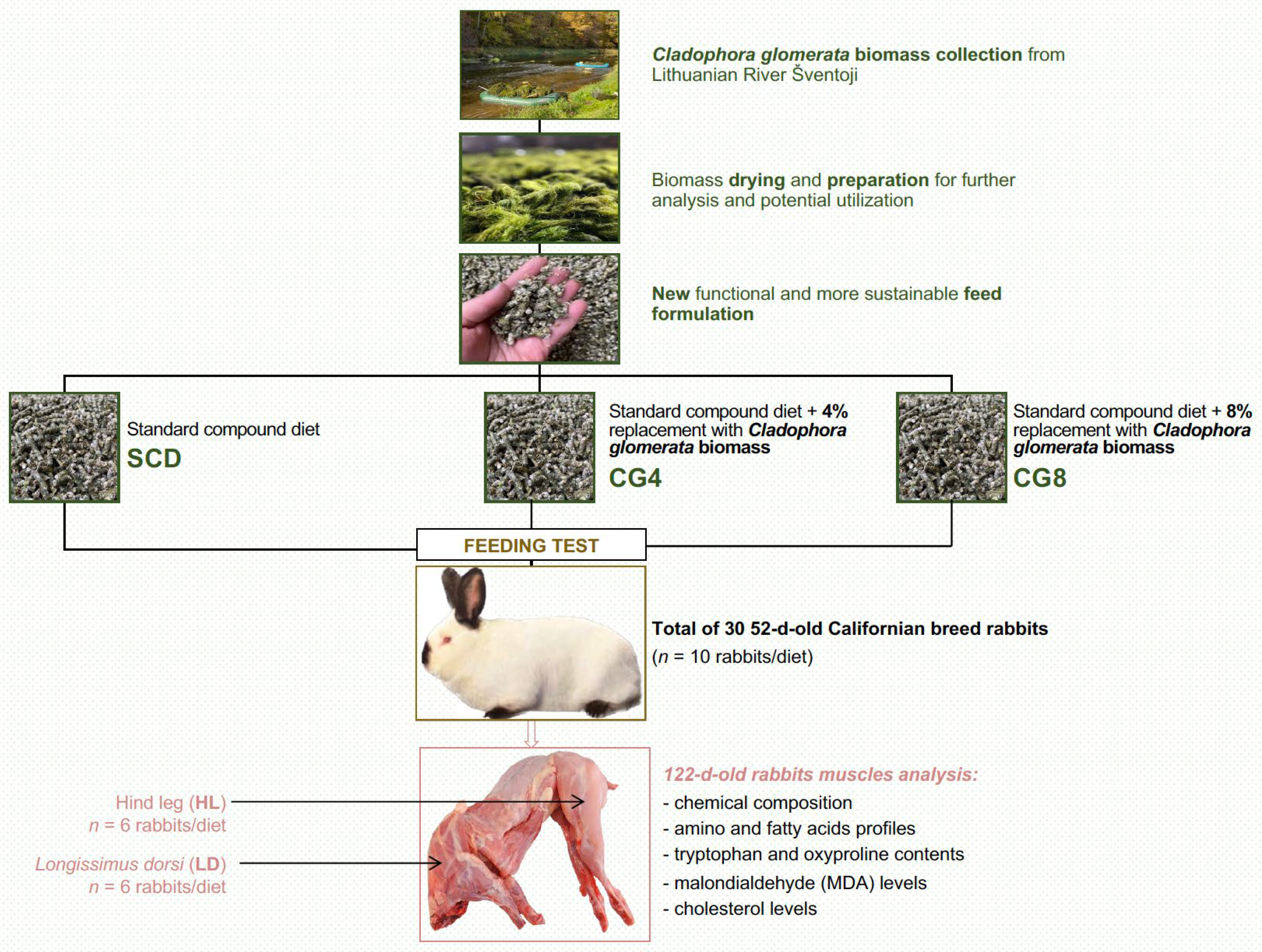
2.1. Animal and Experimental Treatments
2.2. Samples Collection
2.3. Reagents
2.4. Chemical Assay of Muscles
2.5. Amino Acid Profile
2.6. Fatty Acid Profile
2.7. Cholesterol and Lipid Oxidation Levels
2.8. Statistical Analysis
3. Results
3.1. Chemical Composition of Rabbit Muscles
3.2. Amino Acid Profile of Rabbit Muscles
3.3. Lipids in Rabbit Muscles
3.3.1. Fatty Acid Profile
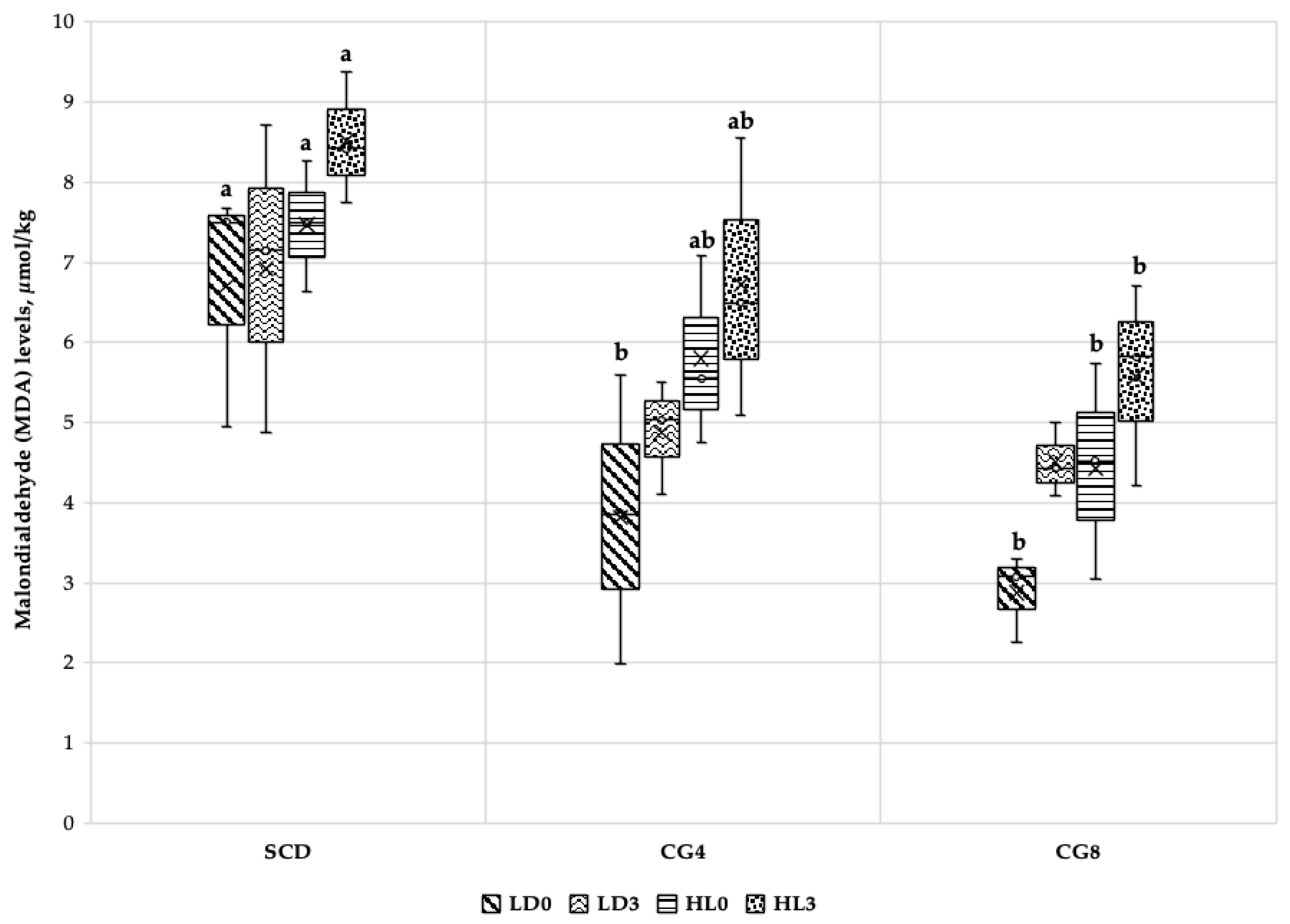
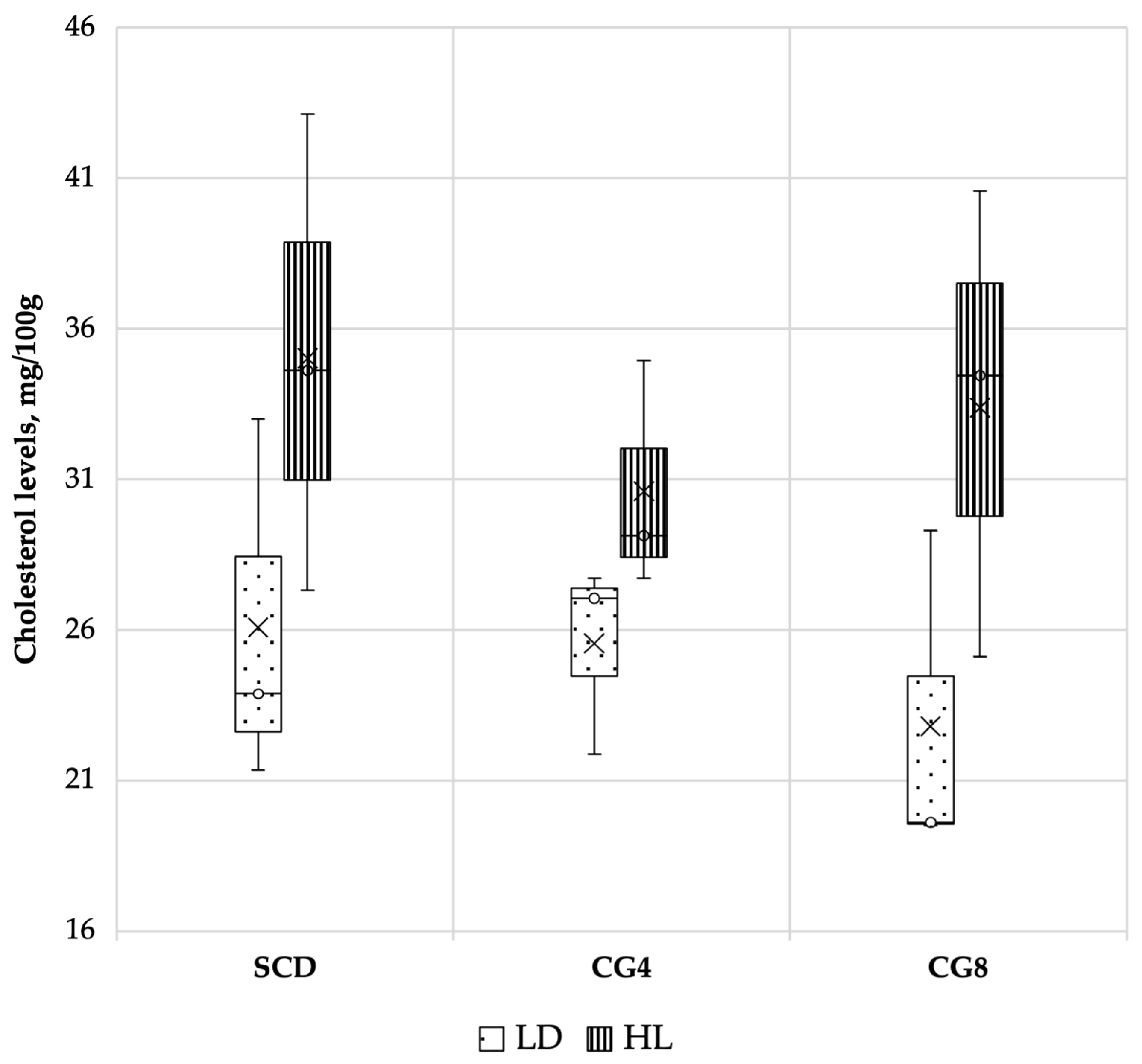
3.3.2. Malondialdehyde (MDA) and Cholesterol Levels
4. Discussion
4.1. Dry Matter of Rabbit Muscles
4.2. Proteins and Amino Acid Profiles of Rabbit Muscles
4.3. Lipids and Fatty Acid Profile of Rabbit Muscles
5. Conclusions
- A 4% inclusion can significantly increase protein and total amino acid levels in rabbits’ muscles while also increasing levels of essential amino acids (threonine, valine, methionine, lysine, and isoleucine) and tryptophan but decreasing hydroxyproline. As a result of their improved and balanced essential amino acid content, rabbit muscle proteins have a higher biological value, resulting in simpler digestion.
- Synergism between alternative freshwater C. glomerata inclusion in rabbit feed can lower the fat content of rabbits’ muscles; the greater the dosage of biomass in the diet, the lower the fat accumulation. Since biomass has a reduced fat content, it can reduce lipid oxidation levels in both fresh and stored muscles.
- Inclusions of 4% and 8% can gradually decrease SFA and MUFA levels in rabbit muscles while significantly enhancing PUFA, which indicates a more nutritionally advantageous quality of lipids. Rabbit meat under both C. glomerata treatments can result in increased heart disease prevention abilities, as observed PUFA/SFA and h/H values were greater, and TI and AI were lower.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Korczyński, M.; Witkowska, Z.; Opaliński, S.; Świniarska, M.; Dobrzański, Z. Algae Extract as a Potential Feed Additive. In Marine Algae Extracts; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2015; pp. 603–626. [Google Scholar]
- Michalak, I.; Messyasz, B. Concise review of Cladophora spp.: Macroalgae of commercial interest. J. Appl. Phycol. 2021, 33, 133–166. [Google Scholar] [CrossRef]
- Laungsuwon, R.; Chulalaksananukul, W. Chemical composition and antibacterial activity of extracts of freshwater green algae, Cladophora glomerata Kützing andMicrospora floccosa (Vaucher) Thuret. J. BioScience Biotechnol. 2014, 3, 211–218. [Google Scholar]
- Konkol, D.; Górniak, W.; Świniarska, M.; Korczyński, M. Algae Biomass in Animal Production. In Algae Biomass: Characteristics and Applications; Springer International Publishing: Cham, Switzerland, 2018; pp. 123–130. [Google Scholar]
- Kovač, D.J.; Simeunović, J.B.; Babić, O.B.; Mišan, A.Č.; Milovanović, I.L. Algae in food and feed. Food Feed. Res. 2013, 40, 21–32. [Google Scholar]
- Bruneel, C.; Lemahieu, C.; Fraeye, I.; Ryckebosch, E.; Muylaert, K.; Buyse, J.; Foubert, I. Impact of microalgal feed supplementation on omega-3 fatty acid enrichment of hen eggs. J. Funct. Foods 2013, 5, 897–904. [Google Scholar] [CrossRef]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Innovative Natural Functional Ingredients from Microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of high added-value compounds—A brief review of recent work. Biotechnol. Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef]
- Wan, A.H.L.; Davies, S.J.; Soler-Vila, A.; Fitzgerald, R.; Johnson, M.P. Macroalgae as a sustainable aquafeed ingredient. Rev. Aquac. 2019, 11, 458–492. [Google Scholar] [CrossRef]
- Mihranyan, A. Cellulose from cladophorales green algae: From environmental problem to high-tech composite materials. J. Appl. Polym. Sci. 2011, 119, 2449–2460. [Google Scholar] [CrossRef]
- Zulkifly, S.B.; Graham, J.M.; Young, E.B.; Mayer, R.J.; Piotrowski, M.J.; Smith, I.; Graham, L.E. The Genus Cladophora Kützing (Ulvophyceae) as a Globally Distributed Ecological Engineer. J. Phycol. 2013, 49, 1–17. [Google Scholar] [CrossRef]
- Pikosz, M.; Messyasz, B.; Gąbka, M. Functional structure of algal mat (Cladophora glomerata) in a freshwater in western Poland. Ecol. Indic. 2017, 74, 1–9. [Google Scholar] [CrossRef]
- Messyasz, B.; Leska, B.; Fabrowska, J.; Pikosz, M.; Roj, E.; Cieslak, A.; Schroeder, G. Biomass of freshwater Cladophora as a raw material for agriculture and the cosmetic industry. Open Chem. 2015, 13, 1108–1118. [Google Scholar] [CrossRef]
- Akköz, C.; Arslan, D.; Ünver, A.; Özcan, M.M.; Yilmaz, B. Chemical composition, total phenolic and mineral contents of Enteromorpha intestinalis (L.) Kütz. and Cladophora glomerata (L.) Kütz. seaweeds. J. Food Biochem. 2011, 35, 513–523. [Google Scholar] [CrossRef]
- Nutautaitė, M.; Racevičiūtė-Stupelienė, A.; Bliznikas, S.; Jonuškienė, I.; Karosienė, J.; Koreivienė, J.; Vilienė, V. Evaluation of Phenolic Compounds and Pigments in Freshwater Cladophora glomerata Biomass from Various Lithuanian Rivers as a Potential Future Raw Material for Biotechnology. Water 2022, 14, 1138. [Google Scholar] [CrossRef]
- Nutautaitė, M.; Vilienė, V.; Racevičiūtė-Stupelienė, A.; Bliznikas, S.; Karosienė, J.; Koreivienė, J. Freshwater Cladophora glomerata Biomass as Promising Protein and Other Essential Nutrients Source for High Quality and More Sustainable Feed Production. Agriculture 2021, 11, 582. [Google Scholar] [CrossRef]
- Anh, N.; Hai, T.; Hien, T. Effects of partial replacement of fishmeal protein with green seaweed (Cladophora spp.) protein in practical diets for the black tiger shrimp (Penaeus monodon) postlarvae. J. Appl. Phycol. 2018, 30, 2649–2658. [Google Scholar] [CrossRef]
- Appler, H.N.; Jauncey, K. The utilization of a filamentous green alga (Cladophora glomerata (L) Kutzin) as a protein source in pelleted feeds for Sarotherodon (Tilapia) niloticus fingerlings. Aquaculture 1983, 30, 21–30. [Google Scholar] [CrossRef]
- Promya, J.; Chitmanat, C. The effects of Spirulina platensis and Cladophora algae on the growth performance, meat quality and immunity stimulating capacity of the African sharptooth catfish (Clarias gariepinus). Int. J. Agric. Biol. 2011, 13, 77–82. [Google Scholar]
- Al-Soufi, S.; García, J.; Muíños, A.; López-Alonso, M. Marine Macroalgae in Rabbit Nutrition—A Valuable Feed in Sustainable Farming. Animals 2022, 12, 2346. [Google Scholar] [CrossRef]
- Rossi, R.; Vizzarri, F.; Chiapparini, S.; Ratti, S.; Casamassima, D.; Palazzo, M.; Corino, C. Effects of dietary levels of brown seaweeds and plant polyphenols on growth and meat quality parameters in growing rabbit. Meat Sci. 2020, 161, 107987. [Google Scholar] [CrossRef]
- Moroney, N.C.; O’Grady, M.N.; Robertson, R.C.; Stanton, C.; O’Doherty, J.V.; Kerry, J.P. Influence of level and duration of feeding polysaccharide (laminarin and fucoidan) extracts from brown seaweed (Laminaria digitata) on quality indices of fresh pork. Meat Sci. 2015, 99, 132–141. [Google Scholar] [CrossRef]
- Wang, J.; Su, Y.; Elzo, M.A.; Jia, X.; Chen, S.; Lai, S. Comparison of Carcass and Meat Quality Traits among Three Rabbit Breeds. Korean J. Food Sci. Anim. Resour. 2016, 36, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Dalle Zotte, A.; Szendrő, Z. The role of rabbit meat as functional food. Meat Sci. 2011, 88, 319–331. [Google Scholar] [CrossRef] [PubMed]
- de Blas, C.; Wiseman, J.; Carabano, R.; Abad-Guamán, R.; Allain, D.; Badiola, I.; Blas, E.; Cervera, C.; Zotte, A.D.; Carmona, J.F. Nutrition of the Rabbit, 3rd ed.; CAB International: Oxford, UK, 2020; pp. 243–254. [Google Scholar]
- Arrington, L.R.; Cheeke, P.R.; Lebas, F.; Lebas, S. Nutrient Requirements of Rabbits, 2nd ed.; National Reasearch Council (NRC): Washington, DC, USA, 1977. [Google Scholar]
- Nutautaitė, M.; Vilienė, V.; Racevičiūtė-Stupelienė, A.; Bliznikas, S.; Karosienė, J.; Koreivienė, J. Cladophora glomerata as a potential nutrient source in animal nutrition. In Proceedings of the 1st International Ph.D. Student’s Conference at the University of Life Sciences, Lublin, Poland, 26 April 2022; Environment–Plant–Animal–Product. University of Life Sciences in Lublin: Lublin, Poland, 2022; p. A023. [Google Scholar]
- Blasco, A.; Ouhayoun, J. Harmonization of criteria and terminology in rabbit meat research. Revised proposal. World Rabbit. Sci. 2010, 4, 93–99. [Google Scholar] [CrossRef] [Green Version]
- King-Brink, M.; Sebranek, J.G. Combustion method for determination of crude protein in meat and meat products: Collaborative study. J. AOAC Int. 1993, 76, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, W. Official Methods of Analysis of AOAC International, 18th ed.; Current Through Rev. 2, 2007 ed.; AOAC International: Gaithersburg, MD, USA, 2005; pp. 24–36. [Google Scholar]
- Stegemann, H.; Stalder, K. Determination of hydroxyproline. Clin. Chim. Acta 1967, 18, 267–273. [Google Scholar] [CrossRef]
- Miller, E.L. Determination of the tryptophan content of feedingstuffs with particular reference to cereals. J. Sci. Food Agric. 1967, 18, 381–386. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christopherson, S.W.; Glass, R.L. Preparation of milk fat methylesters by alcoholysis in an essentially nonalcoholic solution. J. Dairy Sci. 1969, 52, 1289–1290. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Fernández, M.; Ordóñez, J.A.; Cambero, I.; Santos, C.; Pin, C.; Hoz, L.D.L. Fatty acid compositions of selected varieties of Spanish dry ham related to their nutritional implications. Food Chem. 2007, 101, 107–112. [Google Scholar] [CrossRef]
- Polak, T.; Rajar, A.; Gašperlin, L.; Žlender, B. Cholesterol concentration and fatty acid profile of red deer (Cervus elaphus) meat. Meat Sci. 2008, 80, 864–869. [Google Scholar] [CrossRef]
- Mendes, R.; Cardoso, C.; Pestana, C. Measurement of malondialdehyde in fish: A comparison study between HPLC methods and the traditional spectrophotometric test. Food Chem. 2009, 112, 1038–1045. [Google Scholar] [CrossRef]
- Khaled, A.Y.; Parrish, C.A.; Adedeji, A. Emerging nondestructive approaches for meat quality and safety evaluation—A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3438–3463. [Google Scholar] [CrossRef]
- Huang, L.; Zhao, J.; Chen, Q.; Zhang, Y. Rapid detection of total viable count (TVC) in pork meat by hyperspectral imaging. Food Res. Int. 2013, 54, 821–828. [Google Scholar] [CrossRef]
- Abu Hafsa, S.H.; Khalel, M.S.; El-Gindy, Y.M.; Hassan, A.A. Nutritional potential of marine and freshwater algae as dietary supplements for growing rabbits. Ital. J. Anim. Sci. 2021, 20, 784–793. [Google Scholar] [CrossRef]
- Fogliano, V.; Vitaglione, P. Functional foods: Planning and development. Mol. Nutr. Food Res. 2005, 49, 256–262. [Google Scholar] [CrossRef]
- Morshdy, A.E.M.A.; Darwish, W.S.; El Sebay, E.S.S.; Mesallam Ali, E.S. Amino acid profile of rabbit meat: Dietary intake and the effect of freezing on the amino acid composition. Thai J. Vet. Med. 2022, 52, 251–258. [Google Scholar]
- Wu, G.; Bazer, F.W.; Cross, H.R. Land-based production of animal protein: Impacts, efficiency, and sustainability. Ann. N. Y. Acad. Sci. 2014, 1328, 18–28. [Google Scholar] [CrossRef]
- Elango, R.; Ball, R.; Pencharz, P. Amino acid requirements in humans: With a special emphasis on the metabolic availability of amino acids. Amino Acids 2009, 37, 19–27. [Google Scholar] [CrossRef]
- Hernández, P.; Zotte, A.D. Influence of diet on rabbit meat quality. In Nutrition of the Rabbit; CABI: Wallingford, UK, 2010; pp. 163–178. [Google Scholar]
- Nutautaitė, M.; Alijošius, S.; Bliznikas, S.; Šašytė, V.; Vilienė, V.; Pockevičius, A.; Racevičiūtė-Stupelienė, A. Effect of betaine, a methyl group donor, on broiler chicken growth performance, breast muscle quality characteristics, oxidative status and amino acid content. Ital. J. Anim. Sci. 2020, 19, 621–629. [Google Scholar] [CrossRef]
- Sun, H.; Yang, W.R.; Yang, Z.B.; Wang, Y.; Jiang, S.Z.; Zhang, G.G. Effects of betaine supplementation to methionine deficient diet on growth performance and carcass characteris-tics of broilers. Am. J. Anim. Vet. Sci. 2008, 3, 78–84. [Google Scholar]
- Elango, R.; Ball, R.O.; Pencharz, P.B. Recent advances in determining protein and amino acid requirements in humans. Br. J. Nutr. 2012, 108, S22–S30. [Google Scholar] [CrossRef] [PubMed]
- Elango, R. Methionine Nutrition and Metabolism: Insights from Animal Studies to Inform Human Nutrition. J. Nutr. 2020, 150, 2518S–2523S. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino Acids, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Bohrer, B.M. Review: Nutrient density and nutritional value of meat products and non-meat foods high in protein. Trends Food Sci. Technol. 2017, 65, 103–112. [Google Scholar] [CrossRef]
- Li, S.; He, Z.; Hu, Y.; Li, H. Shotgun proteomic analysis of protein profile changes in female rabbit meat: The effect of breed and age. Ital. J. Anim. Sci. 2019, 18, 1335–1344. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J. Taste-active peptides and amino acids of pork meat as components of dry-cured meat products: An in-silico study. J. Sens. Stud. 2017, 32, e12301. [Google Scholar] [CrossRef]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations—A review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]
- Vinauskiene, R.; Leskauskaite, D.; Akromaite, E. Nutritional composition of farm chinchilla (Chinchilla lanigera) meat. J. Food Compos. Anal. 2019, 84, 103303. [Google Scholar] [CrossRef]
- Saad, M.S.; Hassan, M.A.; Amin, R.A.; El-Shater, M.A.; Shanab, M.S. Detection of starch and hydroxyproline content in some meat products. Benha Vet. Med. J. 2018, 35, 195–201. [Google Scholar] [CrossRef]
- Messia, M.C.; Marconi, E. Innovative and Rapid Procedure for 4-Hydroxyproline Determination in Meat-Based Foods. In Amino Acid Analysis; Humana Press: Totowa, NJ, USA, 2011; Volume 828, pp. 281–289. [Google Scholar]
- Zaitsev, S.; Bogolyubova, N. Estimation of the Amino Acid Composition of Pig Meat and Its Correlation with Pork Quality. In Fundamental and Applied Scientific Research in the Development of Agriculture in the Far East (AFE-2021); Springer International Publishing: Cham, Switzerland, 2021; pp. 459–466. [Google Scholar]
- Nistor, E.; Bampidis, V.A.; Păcală, N.; Pentea, M.; Tozer, J.; Prundeanu, H. Nutrient Content of Rabbit Meat as Compared to Chicken, Beef and Pork Meat. J. Anim. Prod. Adv. 2013, 3, 172. [Google Scholar] [CrossRef]
- Hernández, P.; Gondret, F. Rabbit Meat Quality. In Recent Advances in Rabbit Sciences; Maertens, L., Coudert, P., Eds.; ILVO: Merelbeke, Belgium, 2006; pp. 269–290. [Google Scholar]
- Poławska, E.; Cooper, R.G.; Jóźwik, A.; Pomianowski, J. Meat from alternative species-nutritive and dietetic value, and its benefit for human health—A review. CYTA J. Food 2013, 11, 37–42. [Google Scholar] [CrossRef]
- Dalle Zotte, A. Perception of rabbit meat quality and major factors influencing the rabbit carcass and meat quality. Livest. Prod. Sci. 2002, 75, 11–32. [Google Scholar] [CrossRef]
- Para, A.P.; Ganguly, S.; Wakchaure, R.; Sharma, R.; Mahajam, T.; Praveen, P.K. Rabbit Meat has the Potential of Being a Possible Alternative to Other Meats as a Protein Source: A Brief Review. Int. J. Pharm. Biomed. Res. 2015, 2, 17–19. [Google Scholar]
- Khan, R.S.; Grigor, J.; Winger, R.; Win, A. Functional food product development—Opportunities and challenges for food manufacturers. Trends Food Sci. Technol. 2013, 30, 27–37. [Google Scholar] [CrossRef]
- Prachayasittikul, V.; Prachayasittikul, S.; Ruchirawat, S.; Prachayasittikul, V. Coriander (Coriandrum sativum): A promising functional food toward the well-being. Food Res. Int. 2018, 105, 305–323. [Google Scholar] [CrossRef]
- Ali, M.; Imran, M.; Nadeem, M.; Khan, M.K.; Sohaib, M.; Suleria, H.A.R.; Bashir, R. Oxidative stability and Sensoric acceptability of functional fish meat product supplemented with plant−based polyphenolic optimal extracts. Lipids Health Dis. 2019, 18, 35. [Google Scholar] [CrossRef] [Green Version]
- Khajavi, M.Z.; Abhari, K.; Barzegar, F.; Hosseini, H. Functional Meat Products: The New Consumer’s Demand. Curr. Nutr. Food Sci. 2020, 16, 260–267. [Google Scholar] [CrossRef]
- Fike, J.H.; Saker, K.E.; O’Keefe, S.F.; Marriott, N.G.; Ward, D.L.; Fontenot, J.P.; Veit, H.P. Effects of Tasco (a seaweed extract) and heat stress on N metabolism and meat fatty acids in wether lambs fed hays containing endophyte-infected fescue. Small Rumin. Res. 2005, 60, 237–245. [Google Scholar] [CrossRef]
- Huang, C.B.; Ebersole, J.L. A novel bioactivity of omega-3 polyunsaturated fatty acids and their ester derivatives. Mol. Oral Microbiol. 2010, 25, 75–80. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n−3 and n−6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Mozaffarian, D.; WU, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on Risk Factors, Molecular Pathways, and Clinical Events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Zuniga, E.; Ojima, I. Novel Taxoid-Based Tumor-Targeting Drug Conjugates. Chim. Oggi 2009, 27, 54–56. [Google Scholar] [PubMed]
- Hernandez, P. Enhancement of Nutritional Quality and Safety in Rabbit Meat. In Proceedings of the 9th World Rabbit Congress, Verona, Italy, 10–13 June 2008; pp. 1287–1299. [Google Scholar]
- Chen, J.; Liu, H. Nutritional Indices for Assessing Fatty Acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef] [PubMed]
- Omri, B.; Chalghoumi, R.; Izzo, L.; Ritieni, A.; Lucarini, M.; Durazzo, A.; Abdouli, H.; Santini, A. Effect of Dietary Incorporation of Linseed Alone or Together with Tomato-Red Pepper Mix on Laying Hens’ Egg Yolk Fatty Acids Profile and Health Lipid Indexes. Nutrients 2019, 11, 813. [Google Scholar] [CrossRef]
- González-Félix, M.L.; Maldonado-Othón, C.A.; Perez-Velazquez, M. Effect of dietary lipid level and replacement of fish oil by soybean oil in compound feeds for the shortfin corvina (Cynoscion parvipinnis). Aquaculture 2016, 454, 217–228. [Google Scholar] [CrossRef]
- Yurchenko, S.; Sats, A.; Tatar, V.; Kaart, T.; Mootse, H.; Jõudu, I. Fatty acid profile of milk from Saanen and Swedish Landrace goats. Food Chem. 2018, 254, 326–332. [Google Scholar] [CrossRef]
- Šašytė, V.; Racevičiūtė Stupelienė, A.; Vilienė, V.; Daukšienė, A.; Gružauskas, R.; Alijošius, S. The Effect of extruded full-fat rapeseed on productivity and eggs quality of Isa brown laying hens. In Proceedings of the 19th International Conference on Animal Nutrition (ICAN 2017), Dubai, United Arab Emirates, 30–31 January 2017; World Academy of Science, Engineering and Technology: Dubai, United Arab Emirates, 2017; Volume 19, pp. 2954–2961. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Diet 3 | |||
|---|---|---|---|
| Ingredient (%) 1,2 | SCD | CG4 | CG8 |
| Corn | 3.00 | 3.13 | 3.44 |
| Barley | 18.00 | 18.00 | 18.00 |
| Oats | 25.00 | 25.00 | 25.00 |
| Sunflower meal | 13.22 | 11.97 | 9.97 |
| Linseed meal | 1.00 | 1.00 | 1.00 |
| Soy meal | 3.72 | 3.00 | 3.00 |
| Vegetable oil | 1.00 | 1.00 | 1.00 |
| Beer east | 2.00 | 2.00 | 2.00 |
| Hay | 29.26 | 27.10 | 24.79 |
| C. glomerata | – | 4.00 | 8.00 |
| Antimycotoxin | 0.30 | 0.30 | 0.30 |
| Vitamin-mineral premix | 3.50 | 3.50 | 3.50 |
| Total | 100 | 100 | 100 |
| Chemical composition (%) | |||
| DE (MJ/kg) | 10.49 | 11.13 | 12.16 |
| Crude protein | 17.54 | 17.53 | 17.54 |
| Crude fibre | 13.56 | 14.39 | 15.05 |
| Ash | 10.03 | 10.37 | 10.31 |
| Ether extract | 3.12 | 3.20 | 3.10 |
| NDF | 32.49 | 34.19 | 35.89 |
| ADF | 19.71 | 20.12 | 20.73 |
| ADL | 4.89 | 4.94 | 5.15 |
| Diet 3,4,5 | ||||||
|---|---|---|---|---|---|---|
| Item (%) 1 | Muscle 2 | SCD | CG4 | CG8 | SEM 6 | p-Value 7 |
| Protein | LD | 21.19 ab | 22.17 a | 20.36 b | 0.51 | 0.003 |
| HL | 20.65 | 19.43 | 19.69 | 0.69 | n.s. | |
| Fat | LD | 1.62 a | 1.15 b | 0.54 c | 0.11 | 0.000 |
| HL | 2.73 a | 1.44 b | 0.83 c | 0.28 | 0.000 | |
| Ash | LD | 1.13 a | 1.11 ab | 1.06 b | 0.03 | 0.026 |
| HL | 1.12 | 1.10 | 1.06 | 0.03 | n.s. | |
| DM | LD | 25.29 a | 24.60 b | 22.42 c | 0.26 | 0.000 |
| HL | 25.63 a | 23.55 b | 22.47 c | 0.31 | 0.000 | |
| Diet 2,3,4 | ||||||
|---|---|---|---|---|---|---|
| Amino Acid (g/kg) | Muscle 1 | SCD | CG4 | CG8 | SEM 5 | p-Value 6 |
| Aspartic | LD | 18.39 a | 18.43 a | 17.13 b | 0.22 | 0.000 |
| HL | 15.59 a | 16.73 b | 16.45 ab | 0.42 | 0.016 | |
| Threonine | LD | 8.58 a | 8.65 a | 8.24b | 0.12 | 0.003 |
| HL | 7.29 | 7.59 | 7.61 | 0.19 | n.s. | |
| Serine | LD | 7.35 | 7.36 | 7.11 | 0.17 | n.s. |
| HL | 6.32 | 6.83 | 6.89 | 0.31 | n.s. | |
| Glutamic | LD | 31.65 ab | 32.63 a | 31.08 b | 0.51 | 0.08 |
| HL | 27.79 a | 29.61 b | 29.54 b | 0.69 | 0.019 | |
| Proline | LD | 7.26 | 7.40 | 7.14 | 0.15 | n.s. |
| HL | 7.57 | 7.82 | 8.02 | 0.31 | n.s. | |
| Glycine | LD | 10.10 | 9.94 | 10.01 | 0.20 | n.s. |
| HL | 11.05 a | 11.65 ab | 12.42 b | 0.51 | 0.017 | |
| Alanine | LD | 10.27 a | 10.31 a | 9.29 b | 0.25 | 0.001 |
| HL | 8.87 a | 9.89 b | 10.14 b | 0.29 | 0.001 | |
| Valine | LD | 10.40 a | 10.42 a | 9.56 b | 0.20 | 0.001 |
| HL | 8.80 | 9.07 | 9.16 | 0.24 | n.s. | |
| Methionine | LD | 8.66 a | 10.68 b | 8.73 a | 0.68 | 0.009 |
| HL | 7.74 | 8.09 | 8.13 | 0.29 | n.s. | |
| Isoleucine | LD | 9.25 a | 9.28 a | 8.44 b | 0.14 | 0.000 |
| HL | 7.50 | 7.88 | 7.91 | 0.23 | n.s. | |
| Leucine | LD | 14.75 a | 14.57 a | 13.55 b | 0.18 | 0.000 |
| HL | 12.10 a | 12.83 b | 12.90 b | 0.33 | 0.030 | |
| Tyrosine | LD | 6.63 ab | 6.69 a | 6.13 b | 0.25 | 0.039 |
| HL | 5.25 a | 6.65 b | 5.70 b | 0.19 | 0.030 | |
| Phenylalanine | LD | 7.47 a | 7.36 a | 6.89 b | 0.10 | 0.000 |
| HL | 6.28 a | 6.60 ab | 6.68 b | 0.18 | 0.043 | |
| Histidine | LD | 8.78 a | 8.50 ab | 8.11 b | 0.19 | 0.003 |
| HL | 7.46 | 7.63 | 7.98 | 0.26 | n.s. | |
| Lysine | LD | 15.86 a | 16.71 b | 15.60 a | 0.28 | 0.001 |
| HL | 13.70 | 14.15 | 14.41 | 0.37 | n.s. | |
| Arginine | LD | 13.90 a | 13.23 ab | 12.67 b | 0.45 | 0.016 |
| HL | 11.99 | 12.13 | 12.19 | 0.40 | n.s. | |
| Total | LD | 189.30 a | 192.16 a | 179.74 b | 2.58 | 0.000 |
| HL | 165.32 a | 174.33 b | 176.13 b | 4.18 | 0.021 | |
| Diet 2,3,4 | ||||||
|---|---|---|---|---|---|---|
| Item (g/kg) | Muscle 1 | SCD | CG4 | CG8 | SEM 5 | p-Value 6 |
| Tryptophan (T) | LD | 29.40 a | 30.84 b | 25.28 c | 0.35 | 0.000 |
| HL | 26.27 a | 22.79 b | 24.02 c | 0.37 | 0.000 | |
| Hydroxyproline (H) | LD | 7.71 a | 6.30 b | 8.85 c | 0.23 | 0.000 |
| HL | 18.71 a | 15.73 b | 16.70 ab | 1.39 | 0.049 | |
| T/H | LD | 3.82 a | 4.91 b | 2.86 c | 0.13 | 0.000 |
| HL | 1.43 | 1.50 | 1.44 | 0.13 | n.s. | |
| Diet 3,4,5 | ||||||
|---|---|---|---|---|---|---|
| Fatty Acid (%) 1 | Muscle 2 | SCD | CG4 | CG8 | SEM 6 | p-Value 7 |
| C10:0 | LD | 0.09 a | 0.06 ab | 0.01 b | 0.02 | 0.011 |
| HL | 0.12 | 0.05 | 0.02 | 0.04 | n.s. | |
| C12:0 | LD | 0.11 a | 0.09 ab | 0.06 b | 0.02 | 0.018 |
| HL | 0.15 a | 0.08 ab | 0.05 b | 0.03 | 0.023 | |
| C14:0 | LD | 2.20 | 2.06 | 1.76 | 0.25 | n.s. |
| HL | 2.18 a | 2.08 ab | 1.58 b | 0.22 | 0.032 | |
| C15:0 | LD | 0.44 a | 0.36 ab | 0.31 b | 0.03 | 0.008 |
| HL | 0.42 a | 0.35 ab | 0.30 b | 0.03 | 0.008 | |
| C16:0 | LD | 26.81 a | 26.03 a | 23.54 b | 0.55 | 0.001 |
| HL | 25.21 a | 25.00 ab | 22.34 b | 0.53 | 0.002 | |
| C17:0 | LD | 0.50 a | 0.44 b | 0.44 b | 0.02 | 0.038 |
| HL | 0.47 a | 0.44 b | 0.42 b | 0.01 | 0.005 | |
| C18:0 | LD | 5.39 a | 6.03 a | 7.57 b | 0.28 | 0.000 |
| HL | 5.73 a | 7.00 b | 8.25 c | 0.41 | 0.001 | |
| C20:0 | LD | 0.08 a | 0.09 ab | 0.10 a | 0.01 | 0.017 |
| HL | 0.06 a | 0.09 b | 0.07 c | 0.00 | 0.001 | |
| C22:00 | LD | n.d. | 0.04 | 0.05 | 0.03 | n.s. |
| HL | 0.04 a | 0.05 a | 0.13 b | 0.03 | 0.016 | |
| Σ SFA | LD | 35.61 a | 35.21 ab | 33.85 b | 0.68 | 0.042 |
| HL | 34.38 a | 35.12 a | 33.15 b | 0.45 | 0.004 | |
| C14:1 n7 | LD | 0.14 a | 0.08 ab | 0.03 b | 0.04 | 0.039 |
| HL | 0.18 a | 0.12 ab | 0.07 b | 0.04 | 0.041 | |
| C16:1 n9 | LD | 0.34 a | 0.33 ab | 0.32 b | 0.01 | 0.032 |
| HL | 0.36 | 0.34 | 0.33 | 0.02 | n.s. | |
| C16:1 n7 | LD | 2.81 a | 2.09 ab | 1.17 b | 0.42 | 0.008 |
| HL | 3.42 a | 2.08 b | 1.24 b | 0.50 | 0.005 | |
| C17:1 n9 | LD | 0.25 a | 0.18 b | 0.19 b | 0.02 | 0.018 |
| HL | 0.26 a | 0.17 b | 0.14 b | 0.03 | 0.010 | |
| C18:1 n9t | LD | 0.30 | 0.31 | 0.41 | 0.06 | n.s. |
| HL | 0.25 a | 0.32 b | 0.34 b | 0.03 | 0.017 | |
| C18:1 n9 | LD | 25.96 a | 22.82 b | 20.82 b | 0.82 | 0.001 |
| HL | 25.34 a | 21.65 b | 19.61 c | 0.68 | 0.000 | |
| C18:1 n7 | LD | 1.08 a | 1.07 a | 1.16 b | 0.03 | 0.022 |
| HL | 1.10 ab | 1.03 a | 1.16b | 0.05 | 0.041 | |
| C20:1 n9 | LD | 0.30 | 0.18 | 0.20 | 0.07 | n.s. |
| HL | 0.20 a | 0.18 ab | 0.16 b | 0.01 | 0.028 | |
| Σ MUFA | LD | 31.18 a | 27.04 b | 24.30 c | 1.01 | 0.000 |
| HL | 31.12 a | 25.90 b | 23.05 c | 0.94 | 0.000 | |
| C18:2 n6t | LD | 0.09 | 0.07 | 0.06 | 0.01 | n.s. |
| HL | 0.07 ab | 0.08 a | 0.06 b | 0.01 | 0.008 | |
| C18:2 n6ct | LD | 0.04 | 0.06 | 0.10 | 0.05 | n.s. |
| HL | 0.08 | 0.02 | 0.14 | 0.08 | n.s. | |
| C18:2 n6 | LD | 24.40 | 26.51 | 24.21 | 1.47 | n.s. |
| HL | 25.46 | 28.26 | 27.59 | 1.37 | n.s. | |
| C18:3 n6 | LD | 0.06 | 0.07 | 0.08 | 0.01 | n.s. |
| HL | 0.06 | 0.06 | 0.06 | 0.02 | n.s. | |
| C18:3 n3 | LD | 2.90 a | 2.50 ab | 1.80 b | 0.37 | 0.025 |
| HL | 3.01 a | 2.44 b | 1.69 c | 0.24 | 0.001 | |
| C20:2 n6 | LD | 0.13 a | 0.12 a | 0.17 b | 0.01 | 0.004 |
| HL | 0.17 a | 0.14 b | 0.19 a | 0.01 | 0.001 | |
| C20:3 n6 | LD | 0.15 a | 0.21 a | 0.39 b | 0.04 | 0.002 |
| HL | 0.21 a | 0.27 a | 0.48 b | 0.04 | 0.001 | |
| C20:3 n3 | LD | 0.03 | 0.01 | n.d. | 0.02 | n.s. |
| HL | 0.05 a | 0.01 b | 0.02 ab | 0.01 | 0.050 | |
| C20:4 n6 | LD | 1.96 a | 3.05 b | 6.40 c | 0.84 | 0.002 |
| HL | 2.23 a | 3.17 a | 6.01 b | 0.67 | 0.001 | |
| C20:5 n3 | LD | 0.06 a | 0.11 b | 0.14 c | 0.02 | 0.019 |
| HL | 0.05 a | 0.08 a | 0.12 b | 0.01 | 0.002 | |
| C22:4 n6 | LD | 0.41 a | 0.39 a | 0.64 b | 0.09 | 0.029 |
| HL | 0.47 a | 0.42 a | 0.72 b | 0.08 | 0.009 | |
| C22:5 n3 | LD | 0.48 a | 0.87 b | 1.62 c | 0.21 | 0.002 |
| HL | 0.58 a | 0.91 b | 1.48 c | 0.15 | 0.001 | |
| Σ PUFA | LD | 30.81 a | 34.18 b | 35.99 b | 0.92 | 0.001 |
| HL | 32.54 a | 36.08 b | 38.92 c | 0.93 | 0.000 | |
| Σ unidentified | LD | 2.48 a | 3.70 a | 5.96 b | 0.89 | 0.008 |
| HL | 2.11 a | 3.02 a | 4.96 b | 0.45 | 0.001 | |
| Σ PUFA/Σ SFA | LD | 0.87 a | 0.97 b | 1.06 b | 0.04 | 0.003 |
| HL | 0.95 a | 1.03 a | 1.17 b | 0.04 | 0.001 | |
| omega-3 (ω-3) | LD | 3.57 | 3.68 | 3.93 | 0.44 | n.s. |
| HL | 3.78 | 3.65 | 3.66 | 0.30 | n.s. | |
| omega-6 (ω-6) | LD | 27.11 a | 30.36 b | 31.90 b | 1.08 | 0.004 |
| HL | 28.61 a | 32.33 b | 35.06 b | 1.16 | 0.001 | |
| (ω-6)/(ω-3) | LD | 7.64 | 8.60 | 8.15 | 1.16 | n.s. |
| HL | 7.65 | 9.03 | 9.57 | 0.99 | n.s. | |
| AI | LD | 0.58 | 0.56 | 0.51 | 0.03 | n.s. |
| HL | 0.54 a | 0.54 a | 0.46 b | 0.02 | 0.022 | |
| TI | LD | 0.86 | 0.86 | 0.82 | 0.02 | n.s. |
| HL | 0.80 a | 0.85 b | 0.80 a | 0.01 | 0.002 | |
| h/H | LD | 1.93 a | 2.00 ab | 2.19 b | 0.08 | 0.017 |
| HL | 2.08 a | 2.10 a | 2.38 b | 0.08 | 0.011 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nutautaitė, M.; Racevičiūtė-Stupelienė, A.; Bliznikas, S.; Vilienė, V. Enhancement of Rabbit Meat Functionality by Replacing Traditional Feed Raw Materials with Alternative and More Sustainable Freshwater Cladophora glomerata Macroalgal Biomass in Their Diets. Foods 2023, 12, 744. https://doi.org/10.3390/foods12040744
Nutautaitė M, Racevičiūtė-Stupelienė A, Bliznikas S, Vilienė V. Enhancement of Rabbit Meat Functionality by Replacing Traditional Feed Raw Materials with Alternative and More Sustainable Freshwater Cladophora glomerata Macroalgal Biomass in Their Diets. Foods. 2023; 12(4):744. https://doi.org/10.3390/foods12040744
Chicago/Turabian StyleNutautaitė, Monika, Asta Racevičiūtė-Stupelienė, Saulius Bliznikas, and Vilma Vilienė. 2023. "Enhancement of Rabbit Meat Functionality by Replacing Traditional Feed Raw Materials with Alternative and More Sustainable Freshwater Cladophora glomerata Macroalgal Biomass in Their Diets" Foods 12, no. 4: 744. https://doi.org/10.3390/foods12040744
APA StyleNutautaitė, M., Racevičiūtė-Stupelienė, A., Bliznikas, S., & Vilienė, V. (2023). Enhancement of Rabbit Meat Functionality by Replacing Traditional Feed Raw Materials with Alternative and More Sustainable Freshwater Cladophora glomerata Macroalgal Biomass in Their Diets. Foods, 12(4), 744. https://doi.org/10.3390/foods12040744