
Role and Mechanism of Cold Plasma in Inactivating Alicyclobacillus acidoterrestris in Apple Juice


Abstract
:1. Introduction
2. Materials and Methods
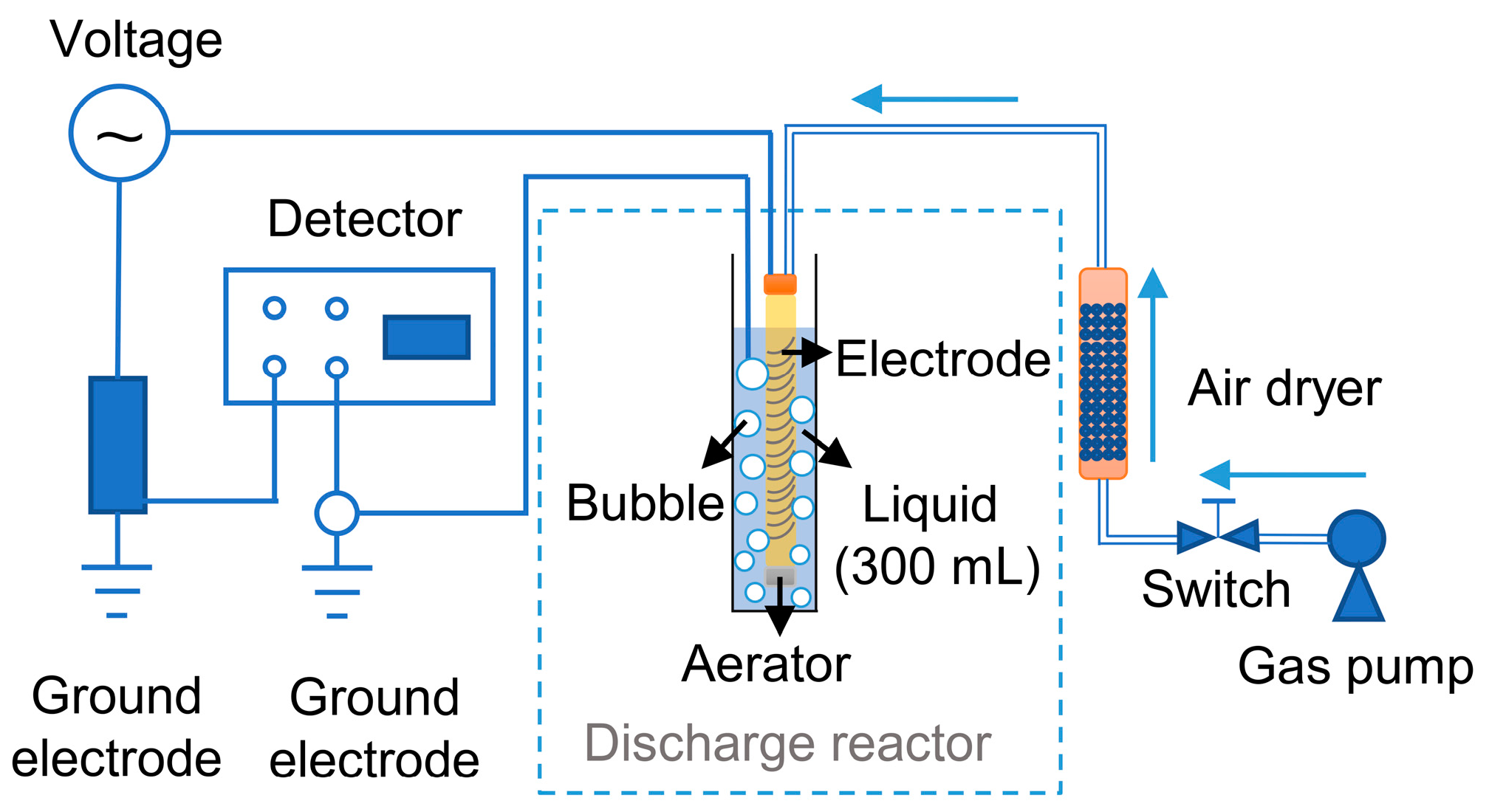
2.1. Cold Atmospheric Plasma System
2.2. Strains and Growth Condition
2.3. Cold Plasma Treatment
2.4. Bacterial Recovery and Enumeration
2.5. Modeling of Inactivation Data
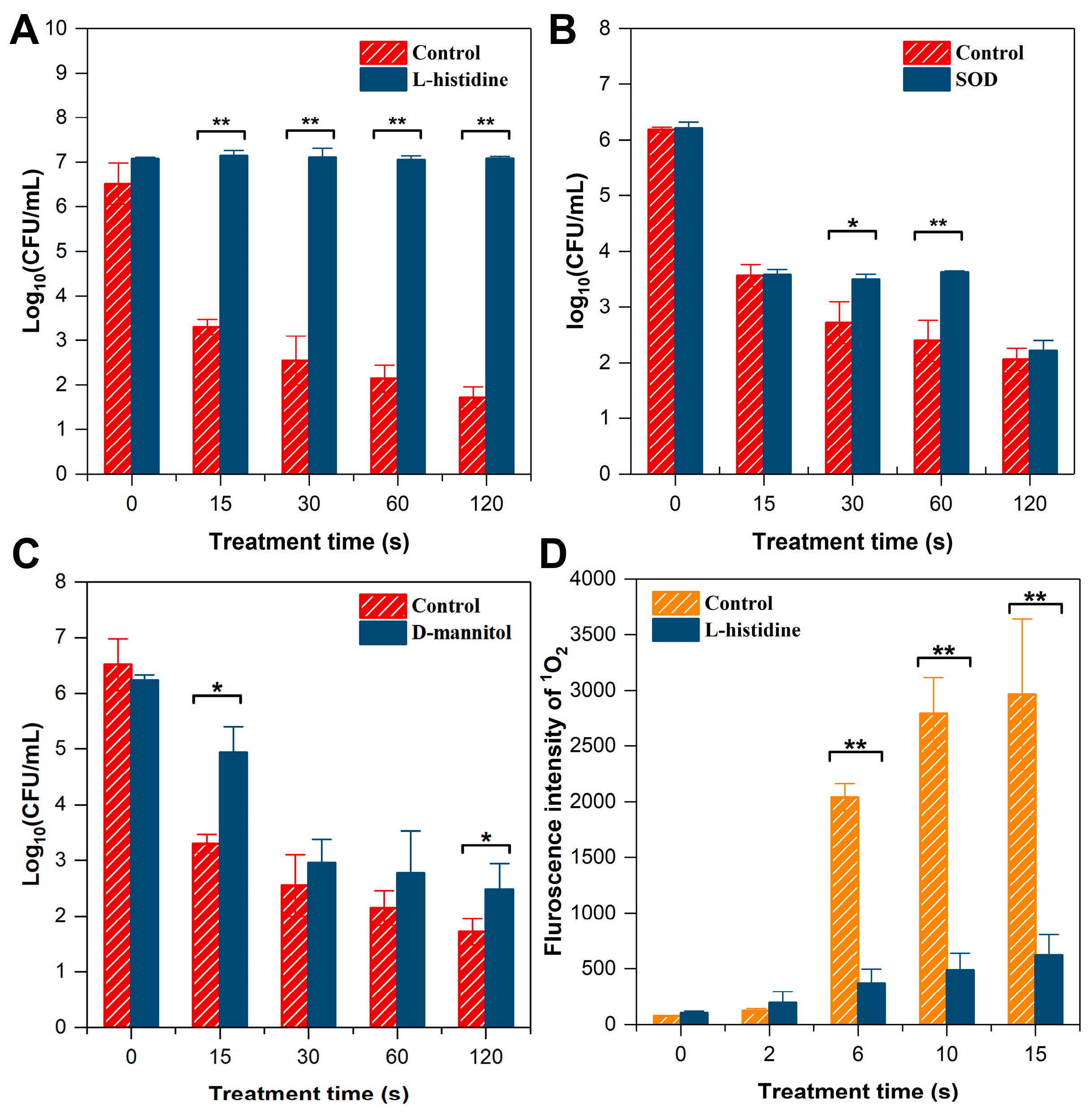
2.6. Scavenging Experiments
2.7. Singlet Oxygen Determination
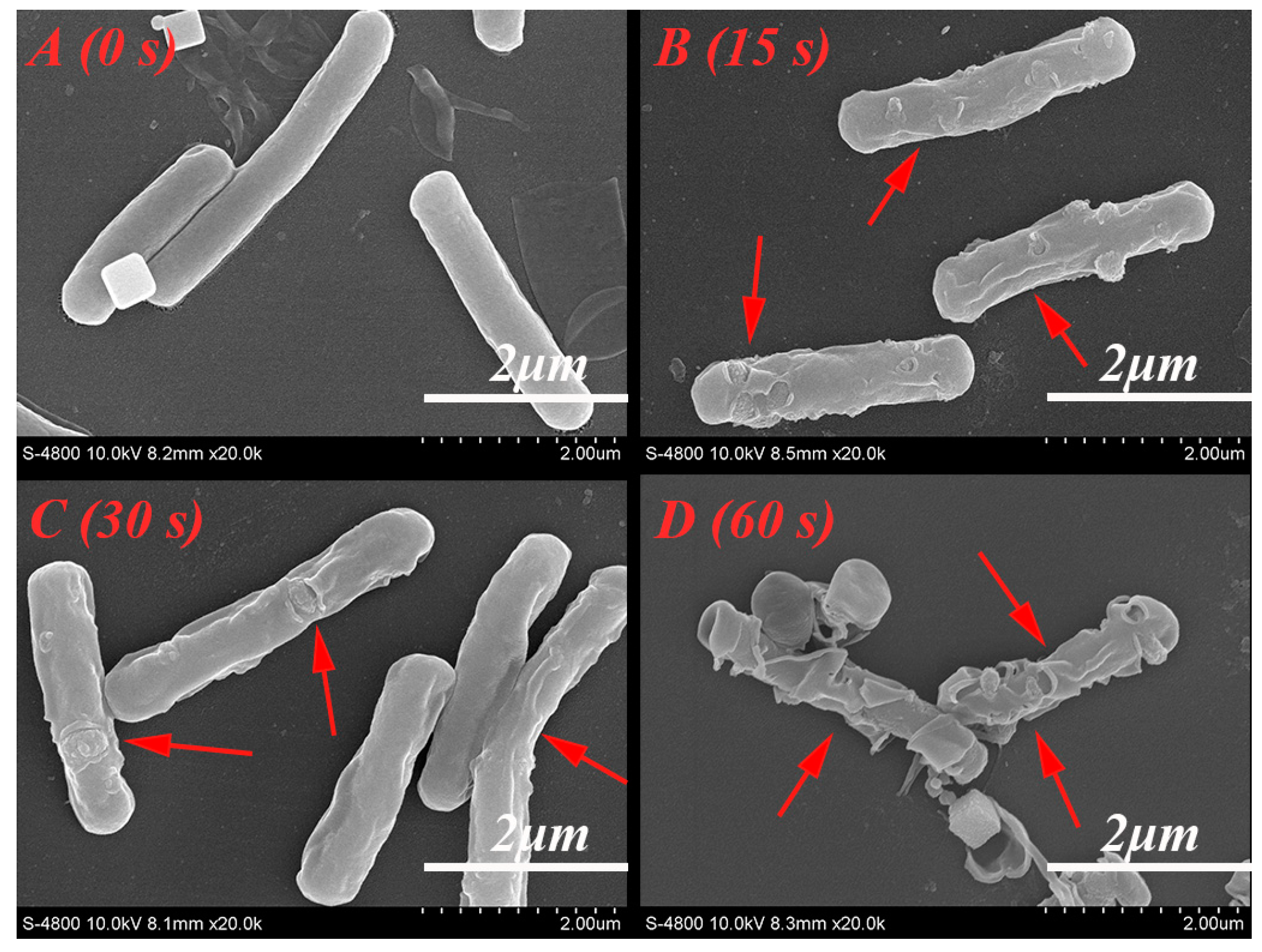
2.8. SEM Analysis
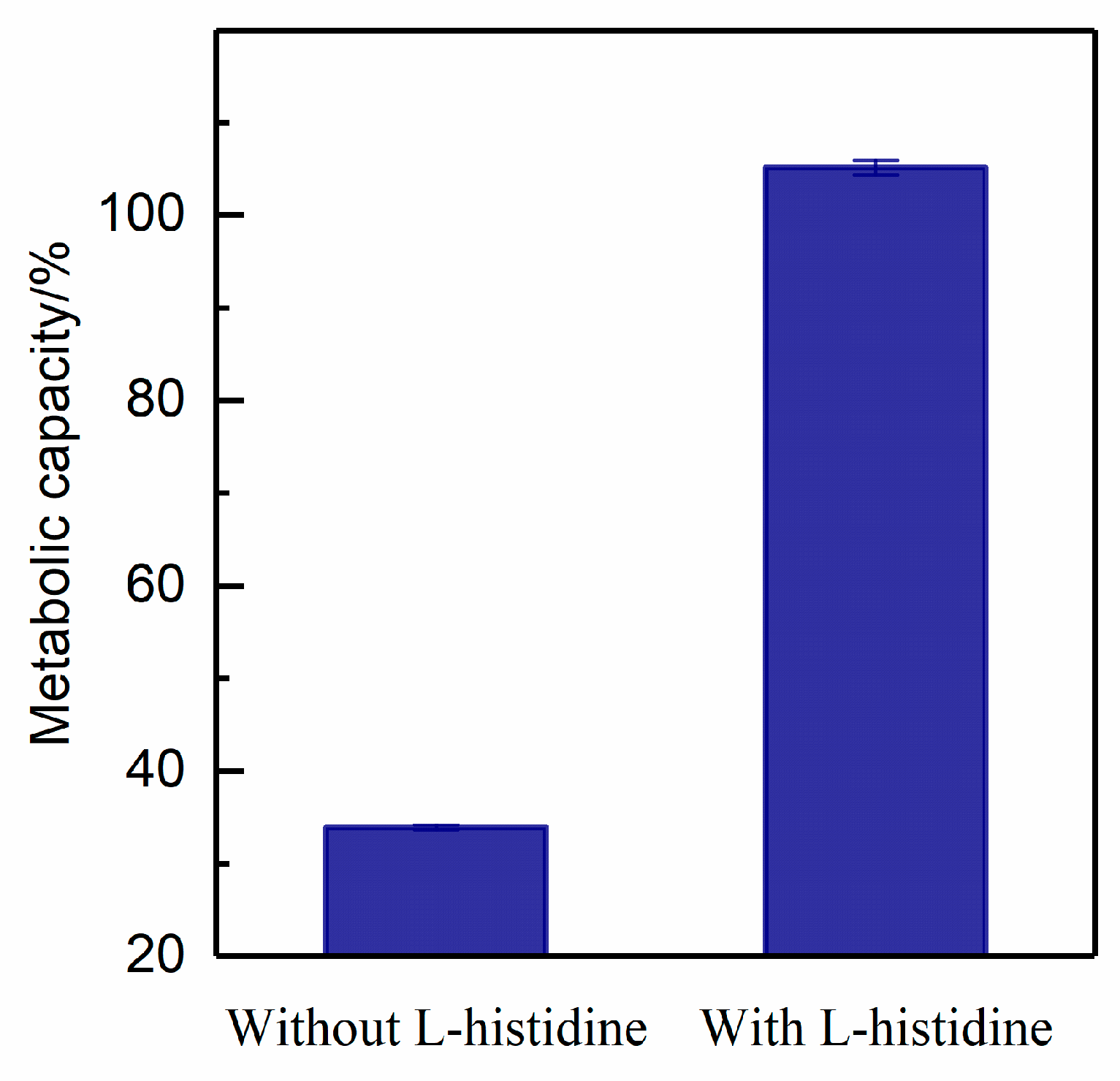
2.9. Metabolic Capacity
2.10. Raman Spectrum Analysis
2.11. Evaluation of Apple Juice Quality
2.12. Statistical Analysis
3. Results
3.1. Inactivation Efficacy of A. acidoterrestris by Cold Plasma
3.2. Kinetic Models
3.3. Inactivation Mechanism of Cold Plasma Used on A. acidoterrestris
3.3.1. Scavenging Experiments
3.3.2. Morphological Changes
3.3.3. Raman Spectra
3.3.4. Metabolic Capacity
3.3.5. The Effects of Cold Plasma Treatment on Apple Juice Quality
3.3.6. Changes in the Quality Parameters of Apple Juice
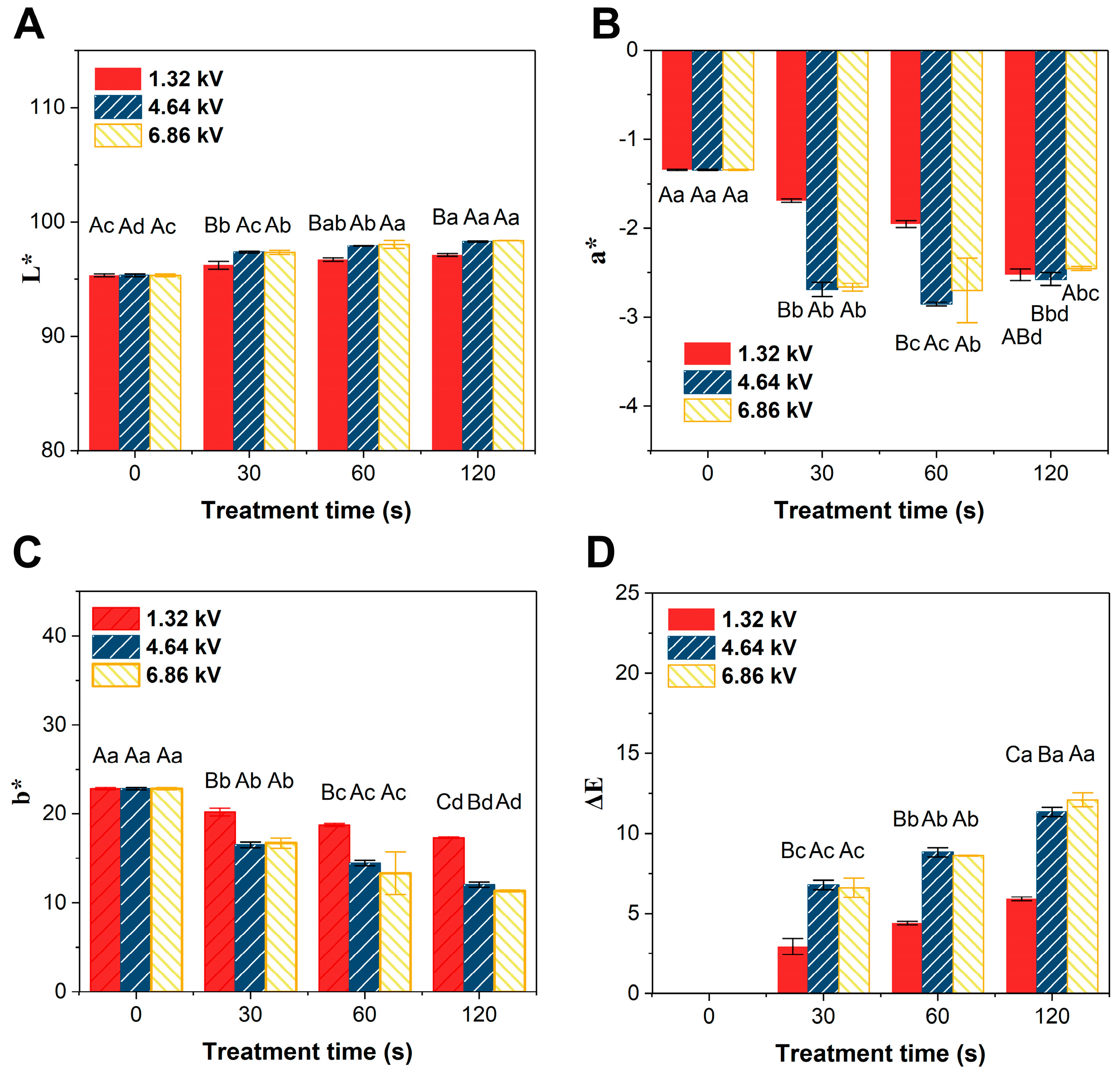
3.3.7. Changes in the Color Value of Apple Juice
4. Discussion
4.1. Inactivation Efficacy of Cold Plasma
4.2. Singlet Oxygen Plays the Most Important Role in Inactivating A. acidoterrestris by Cold Plasma
4.3. The Influence of Cold Plasma on A. acidoterrestris
4.4. Changes in Apple Juice Quality
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yue, T.L.; Zhang, J.B.; Yuan, Y.H. Spoilage by alicyclobacillus bacteria in juice and beverage products: Chemical, physical, and combined control methods. Compr. Rev. Food Sci. Food Saf. 2014, 13, 771–797. [Google Scholar] [CrossRef]
- Sokołowska, B.; Połaska, M.; Dekowska, A. Alicyclobacillus—Still Current Issues in the Beverage Industry. Safety Issues in Beverage Production; Andre Gerhard Wolff: Duxford, UK, 2020; Volume 18, pp. 105–146. [Google Scholar] [CrossRef]
- Chang, S.-S.; Kang, D.-H. Alicyclobacillus spp. in the fruit juice industry: History, characteristics, and current isolation/detection procedures. Crit. Rev. Microbiol. 2004, 30, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Artés-Hernández, F.; Castillejo, N.; Martínez-Zamora, L.; Martínez-Hernández, G.B. Phytochemical fortification in fruit and vegetable beverages with green technologies. Foods 2021, 10, 2534. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.Y.; Li, J.; Muhammad, A.I.; Suo, Y.J.; Chen, S.G.; Ye, X.Q.; Liu, D.H.; Ding, T. Application of a dielectric barrier discharge atmospheric cold plasma (DBD-ACP) for Eshcerichia coli inactivation in apple juice. J. Food Sci. 2018, 83, 401–408. [Google Scholar] [CrossRef]
- Ozen, E.; Singh, R.K. Atmospheric cold plasma treatment of fruit juices: A review. Trends Food Sci. Technol. 2020, 103, 144–151. [Google Scholar] [CrossRef]
- Waghmare, R. Cold plasma technology for fruit based beverages: A review. Trends Food Sci. Technol. 2021, 114, 60–69. [Google Scholar] [CrossRef]
- Xiang, Q.; Liu, X.; Li, J.; Liu, S.; Zhang, H.; Bai, Y. Effects of dielectric barrier discharge plasma on the inactivation of Zygosaccharomyces rouxii and quality of apple juice. Food Chem. 2018, 254, 201–207. [Google Scholar] [CrossRef]
- Surowsky, B.; Frohling, A.; Gottschalk, N.; Schluter, O.; Knorr, D. Impact of cold plasma on Citrobacter freundii in apple juice: Inactivation kinetics and mechanisms. Int. J. Food Microbiol. 2014, 174, 63–71. [Google Scholar] [CrossRef]
- Scholtz, V.; Pazlarova, J.; Souskova, H.; Khun, J.; Julak, J. Nonthermal plasma—A tool for decontamination and disinfection. Biotechnol. Adv. 2015, 33, 1108–1119. [Google Scholar] [CrossRef]
- Liao, X.Y.; Liu, D.H.; Xiang, Q.S.; Ahn, J.; Chen, S.G.; Ye, X.Q.; Ding, T. Inactivation mechanisms of non-thermal plasma on microbes: A review. Food Control. 2017, 75, 83–91. [Google Scholar] [CrossRef]
- Xu, H.; Ma, R.; Zhu, Y.; Du, M.; Zhang, H.; Jiao, Z. A systematic study of the antimicrobial mechanisms of cold atmospheric-pressure plasma for water disinfection. Sci. Total Environ. 2020, 703, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Sun, P.; Feng, H.; Zhou, H.; Wang, R.; Liang, Y.; Lu, J.; Zhu, W.; Zhang, J.; Fang, J. Reactive oxygen species in a non-thermal plasma microjet and water system: Generation, conversion, and contributions to bacteria inactivation—An analysis by electron spin resonance spectroscopy. Plasma Process. Polym. 2012, 9, 417–424. [Google Scholar] [CrossRef]
- Aboubakr, H.A.; Gangal, U.; Youssef, M.M.; Goyal, S.M.; Bruggeman, P.J. Inactivation of virus in solution by cold atmospheric pressure plasma: Identification of chemical inactivation pathways. J. Phys. D Appl. Phys. 2016, 49, 204001. [Google Scholar] [CrossRef]
- Takamatsu, T.; Kawano, H.; Sasaki, Y.; Uehara, K.; Miyahara, H.; Matsumura, Y.; Iwasawa, A.; Azuma, T.; Okino, A. Imaging of the Staphylococcus aureus inactivation process induced by a multigas plasma jet. Curr. Microbiol. 2016, 73, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xu, R.; Gou, L.; Liu, Z.; Zhao, Y.; Liu, D.; Zhang, L.; Chen, H.; Kong, M.G. Mechanism of virus inactivation by cold atmospheric-pressure plasma and plasma-activated water. Appl. Environ. Microbiol. 2018, 84, e00726-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Wang, T.; Qu, G.; Zhang, Y.; Guo, X.; Jia, H.; Zhu, L. Insights into the underlying mechanisms for integrated inactivation of A. spiroides and depression of disinfection byproducts by plasma oxidation. Water Res. 2021, 196, 117027. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Kang, Z.; Jiang, E.; Song, R.; Zhang, Y.; Qu, G.; Wang, T.; Jia, H.; Zhu, L. Plasma induced efficient removal of antibiotic-resistant Escherichia coli and antibiotic resistance genes, and inhibition of gene transfer by conjugation. J. Hazard. Mater. 2021, 419, 126465. [Google Scholar] [CrossRef]
- Cao, Y.; Qian, X.; Zhang, Y.; Qu, G.; Xia, T.; Guo, X.; Jia, H.; Wang, T. Decomplexation of edta-chelated copper and removal of copper ions by non-thermal plasma oxidation/alkaline precipitation. Chem. Soc. Rev. 2019, 362, 487–496. [Google Scholar] [CrossRef]
- Cai, R.; Zhang, M.; Cui, L.; Yuan, Y.; Yang, Y.; Wang, Z.; Yue, T. Antibacterial activity and mechanism of thymol against Alicyclobacillus acidoterrestris vegetative cells and spores. LWT-Food Sci. Technol. 2019, 105, 377–384. [Google Scholar] [CrossRef]
- Cai, R.; Miao, M.; Yue, T.; Zhang, Y.; Cui, L.; Wang, Z.; Yuan, Y. Antibacterial activity and mechanism of cinnamic acid and chlorogenic acid against Alicyclobacillus acidoterrestris vegetative cells in apple juice. Int. J. Food Sci. Technol. 2019, 54, 1697–1705. [Google Scholar] [CrossRef]
- Geeraerd, A.H.; Valdramidis, V.P.; Van Impe, J.F. Ginafit, a freeware tool to assess non-log-linear microbial survivor curves. Int. J. Food Microbiol. 2005, 102, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Fröhling, A.; Baier, M.; Ehlbeck, J.; Knorr, D.; Schlüter, O. Atmospheric pressure plasma treatment of Listeria innocua and Escherichia coli at polysaccharide surfaces: Inactivation kinetics and flow cytometric characterization. Innov. Food Sci. Emerg. Technol. 2012, 13, 142–150. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, J.; Yuan, Y.; Chen, H.; Dai, L.; Wang, X.; Yue, T. Bactericidal effect of cold plasma on microbiota of commercial fish balls. Innov. Food Sci. Emerg. Technol. 2019, 52, 394–405. [Google Scholar] [CrossRef]
- Xu, Z.M.; Cheng, C.; Shen, J.; Lan, Y.; Hu, S.H.; Han, W.; Chu, P.K. In vitro antimicrobial effects and mechanisms of direct current air-liquid discharge plasma on planktonic Staphylococcus aureus and Escherichia coli in liquids. Bioelectrochemistry 2018, 121, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, L.; Mustapha, A.; Li, H.; Hu, Z.Q.; Lin, M. Antibacterial activities of zinc oxide nanoparticles against Escherichia coli o157:H7. J. Appl. Microbiol. 2009, 107, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, Y.; Yuan, Y.; Dai, L.; Yue, T. Characteristic fruit wine production via reciprocal selection of juice and non-Saccharomyces species. Food Microbiol. 2019, 79, 66–74. [Google Scholar] [CrossRef]
- Liao, X.; Liu, D.; Ding, T. Nonthermal plasma (NTP) induces viable but nonculturable (VBNC) state in Staphylococcus aureus via metabolic suppression and oxidative stress response. Appl. Environ. Microbiol. 2019, 86, e02216-19. [Google Scholar] [CrossRef]
- Deng, S.; Ruan, R.; Mok, C.K.; Huang, G.; Lin, X.; Chen, P. Inactivation of Escherichia coli on almonds using nonthermal plasma. J. Food Sci. 2007, 72, M62–M66. [Google Scholar] [CrossRef]
- Han, L.; Patil, S.; Boehm, D.; Milosavljevic, V.; Cullen, P.J.; Bourke, P. Mechanisms of inactivation by high-voltage atmospheric cold plasma differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 450–458. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Wang, Z.; Zhang, W.; Yang, H.; Zhu, X.; Wang, Y. Modeling the effect of initial cell concentration and soluble solids on the plasma inactivation of yeast in apple juices. LWT-Food Sci. Technol. 2021, 151, 112227. [Google Scholar] [CrossRef]
- Cao, Y.; Qu, G.; Li, T.; Jiang, N.; Wang, T. Review on reactive species in water treatment using electrical discharge plasma: Formation, measurement, mechanisms and mass transfer. Plasma Sci. Technol. 2018, 20, 103001. [Google Scholar] [CrossRef] [Green Version]
- Burlica, R.; Kirkpatrick, M.J.; Locke, B.R. Formation of reactive species in gliding arc discharges with liquid water. J. Electrostat. 2006, 64, 35–43. [Google Scholar] [CrossRef]
- Los, A.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Bourke, P.; Dudley, E.G. Inactivation efficacies and mechanisms of gas plasma and plasma-activated water against Aspergillus flavus spores and biofilms: A comparative study. Appl. Environ. Microbiol. 2020, 86, e02619-19. [Google Scholar] [CrossRef] [PubMed]
- Planas, O.; Macia, N.; Agut, M.; Nonell, S.; Heyne, B. Distance-dependent plasmon-enhanced singlet oxygen production and emission for bacterial inactivation. J. Am. Chem. Soc. 2016, 138, 2762–2768. [Google Scholar] [CrossRef] [PubMed]
- Giulio Jori, M.T. Singlet oxygen applications in biosciences and nanosciences. R. Soc. Chem. 2016, 1, 25–67. [Google Scholar]
- Hasegawa, K.; Yamada, K.; Sasase, R.; Miyazaki, R.; Kikuchi, A.; Yagi, M.Y. Direct measurements of absolute concentration and lifetime of singlet oxygen in the gas phase by electron paramagnetic resonance. Chem. Phys. Lett. 2008, 457, 312–314. [Google Scholar] [CrossRef]
- Takamatsu, T.; Uehara, K.; Sasaki, Y.; Hidekazu, M.; Matsumura, Y.; Iwasawa, A.; Ito, N.; Kohno, M.; Azuma, T.; Okino, A. Microbial inactivation in the liquid phase induced by multigas plasma jet. PLoS ONE 2015, 10, e0132381. [Google Scholar] [CrossRef] [Green Version]
- Bauer, G.; Graves, D.B. Mechanisms of selective antitumor action of cold atmospheric plasma-derived reactive oxygen and nitrogen species. Plasma Process. Polym. 2016, 13, 1157–1178. [Google Scholar] [CrossRef]
- Savin, Y.V.; Goryachev, L.V.; Adamenkov, Y.A.; Rakhimova, T.V.; Mankelevich, Y.A.; Popov, N.A.; Adamenkov, A.A.; Egorov, V.V.; Ilyin, S.P.; Kolobyanin, Y.V.; et al. Singlet oxygen production and quenching mechanisms in travelling microwave discharges. J. Phys. D Appl. Phys. 2004, 37, 3121–3128. [Google Scholar] [CrossRef]
- Braginskiy, O.V.; Vasilieva, A.N.; Klopovskiy, K.S.; Kovalev, A.S.; Lopaev, D.V.; Proshina, O.V.; Rakhimova, T.V.; Rakhimov, A.T. Singlet oxygen generation in O2 flow excited by RF discharge: I. Homogeneous discharge mode: A-mode. J. Phys. D Appl. Phys. 2005, 38, 3609–3625. [Google Scholar] [CrossRef]
- Wu, Y.; Liang, Y.; Wei, K.; Li, W.; Yao, M.; Zhang, J.; Grinshpun, S.A. Ms2 virus inactivation by atmospheric-pressure cold plasma using different gas carriers and power levels. Appl. Environ. Microbiol. 2015, 81, 996–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, T.; Yuan, Y.; Fan, Y.; Guo, K.; Yue, T. Inactivation of yeast in apple juice using gas-phase surface discharge plasma treatment with a spray reactor. LWT-Food Sci. Technol. 2018, 97, 530–536. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.; Zhou, H.; Hu, Z.; Shang, W.; Rao, Y.; Peng, H.; Zheng, Y.; Hu, Q.; Zhang, R.; et al. Protective effect of the golden staphyloxanthin biosynthetic pathway on Staphylococcus aureus under cold atmospheric plasma treatment. Appl. Environ. Microbiol. 2019, 86, e01998-19. [Google Scholar] [CrossRef] [PubMed]
- Ozen, E.; Kumar, G.D.; Mishra, A.; Singh, R.K. Inactivation of Escherichia coli in apple cider using atmospheric cold plasma. Int. J. Food Microbiol. 2022, 382, 109913. [Google Scholar] [CrossRef]
- Qian, J.; Ma, L.; Yan, W.; Zhuang, H.; Huang, M.; Zhang, J.; Wang, J. Inactivation kinetics and cell envelope damages of foodborne pathogens Listeria monocytogenes and Salmonella Enteritidis treated with cold plasma. Food Microbiol. 2022, 101, 103891. [Google Scholar] [CrossRef]
- Van Impe, J.; Smet, C.; Tiwari, B.; Greiner, R.; Ojha, S.; Stulic, V.; Vukusic, T.; Rezek Jambrak, A. State of the art of nonthermal and thermal processing for inactivation of micro-organisms. J. Appl. Microbiol. 2018, 125, 16–35. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.-H.; Lv, X.; Pan, Y.; Sun, D.-W. Foodborne bacterial stress responses to exogenous reactive oxygen species (ros) induced by cold plasma treatments. Trends Food Sci. Technol. 2020, 103, 239–247. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Zhu, X.; Yuan, Y.; Gao, Z.; Yue, T. Application of electrical discharge plasma on the inactivation of Zygosaccharomyces rouxii in apple juice. LWT-Food Sci. Technol. 2020, 121, 108974. [Google Scholar] [CrossRef]
- Umair, M.; Jabbar, S.; Nasiru, M.M.; Sultana, T.; Senan, A.M.; Awad, F.N.; Hong, Z.; Zhang, J. Exploring the potential of high-voltage electric field cold plasma (HVCP) using a dielectric barrier discharge (DBD) as a plasma source on the quality parameters of carrot juice. Antibiotics 2019, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Han, Y.; Niu, H.; Zhang, L.; Xiang, Q.; Zong, W. Alternaria mycotoxin degradation and quality evaluation of jujube juice by cold plasma treatment. Food Control. 2022, 137, 108926. [Google Scholar] [CrossRef]
- Pankaj, S.K.; Wan, Z.F.; Keener, K.M. Effects of cold plasma on food quality: A review. Foods 2018, 79, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Input Power (kV) | Kinetic Parameters | Modeling Efficiency | ||||
|---|---|---|---|---|---|---|
| f | kmax1 | kmax2 | lgN0 | RMSE | Adj-R2 | |
| 1.32 | 0.99 ± 0.00 | 0.07 ± 0.01 | 0.00 ± 0.00 | 6.73 ± 0.07 | 0.11 | 0.99 |
| 4.64 | 0.99 ± 0.00 | 0.20 ± 0.50 | 0.00 ± 0.00 | 6.85 ± 0.20 | 0.25 | 0.96 |
| 6.86 | 1.00 ± 0.00 | 0.30 ± 0.05 | 0.00 ± 0.00 | 6.99 ± 0.33 | 0.42 | 0.95 |
| Input Power/kV | Time/s | TSS/% | pH | TS (g/100 mL) | RS (g/100 mL) | TA (g/100 mL) | TP (g/100 mL) |
|---|---|---|---|---|---|---|---|
| 1.32 | |||||||
| 0 | 12.13 ± 0.12 a | 3.74 ± 0.08 a | 10.89 ± 0.15 a | 9.36 ± 0.06 a | 0.24 ± 0.00 a | 0.17 ± 0.01 a | |
| 30 | 12.07 ± 0.12 a | 3.66 ± 0.01 b | 10.85 ± 0.51 a | 9.32 ± 0.19 a | 0.24 ± 0.01 a | 0.17 ± 0.00 b | |
| 60 | 12.02 ± 0.04 a | 3.69 ± 0.05 a | 10.92 ± 0.19 a | 9.23 ± 0.23 a | 0.23 ± 0.00 a | 0.16 ± 0.00 c | |
| 120 | 12.02 ± 0.04 a | 3.78 ± 0.06 a | 10.75 ± 0.24 a | 9.18 ± 0.20 a | 0.23 ± 0.00 a | 0.16 ± 0.00 c | |
| 4.64 | |||||||
| 0 | 12.13 ± 0.12 a | 3.74 ± 0.08 a | 10.89 ± 0.15 ac | 9.36 ± 0.06 b | 0.24 ± 0.00 a | 0.17 ± 0.01 a | |
| 30 | 11.98 ± 0.04 a | 3.70 ± 0.06 a | 10.92 ± 0.49 ac | 9.23 ± 0.20 b | 0.23 ± 0.00 a | 0.16 ± 0.01 b | |
| 60 | 11.80 ± 0.12 a | 3.66 ± 0.02 a | 10.67 ± 0.22 bc | 9.71 ± 0.16 a | 0.24 ± 0.01 a | 0.14 ± 0.01 c | |
| 120 | 11.97 ± 0.05 a | 3.68 ± 0.06 a | 11.28 ± 0.07 a | 9.29 ± 0.15 b | 0.24 ± 0.00 a | 0.13 ± 0.00 c | |
| 6.86 | |||||||
| 0 | 12.13 ± 0.12 a | 3.74 ± 0.08 a | 10.89 ± 0.15 a | 9.36 ± 0.06 a | 0.24 ± 0.00 a | 0.17 ± 0.01 a | |
| 30 | 12.02 ± 0.08 a | 3.68 ± 0.04 a | 10.97 ± 0.39 a | 9.32 ± 0.07 a | 0.24 ± 0.00 a | 0.15 ± 0.01 a | |
| 60 | 11.92 ± 0.04 a | 3.76 ± 0.03 a | 10.30 ± 0.41 b | 9.33 ± 0.15 a | 0.24 ± 0.00 a | 0.14 ± 0.00 b | |
| 120 | 11.92 ± 0.04 a | 3.74 ± 0.03 a | 10.73 ± 0.22 ab | 9.29 ± 0.20 a | 0.24 ± 0.00 a | 0.13 ± 0.00 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, H.; Wang, T.; Sun, Y.; Zhang, Y.; Wei, J.; Cai, R.; Guo, C.; Yuan, Y.; Yue, T. Role and Mechanism of Cold Plasma in Inactivating Alicyclobacillus acidoterrestris in Apple Juice. Foods 2023, 12, 1531. https://doi.org/10.3390/foods12071531
Ding H, Wang T, Sun Y, Zhang Y, Wei J, Cai R, Guo C, Yuan Y, Yue T. Role and Mechanism of Cold Plasma in Inactivating Alicyclobacillus acidoterrestris in Apple Juice. Foods. 2023; 12(7):1531. https://doi.org/10.3390/foods12071531
Chicago/Turabian StyleDing, Hao, Tiecheng Wang, Yuhan Sun, Yuxiang Zhang, Jianping Wei, Rui Cai, Chunfeng Guo, Yahong Yuan, and Tianli Yue. 2023. "Role and Mechanism of Cold Plasma in Inactivating Alicyclobacillus acidoterrestris in Apple Juice" Foods 12, no. 7: 1531. https://doi.org/10.3390/foods12071531
APA StyleDing, H., Wang, T., Sun, Y., Zhang, Y., Wei, J., Cai, R., Guo, C., Yuan, Y., & Yue, T. (2023). Role and Mechanism of Cold Plasma in Inactivating Alicyclobacillus acidoterrestris in Apple Juice. Foods, 12(7), 1531. https://doi.org/10.3390/foods12071531