
Intracellular Antioxidant and Anti-Inflammatory Effects and Bioactive Profiles of Coffee Cascara and Black Tea Kombucha Beverages

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Raw Materials
2.3. Infusions Preparation, Kombucha Consortium, and Fermentation
- Kombucha Consortium and fermentation: The Kombucha Consortium was part of the collection of the Microbiology Institute of the Federal University of Rio de Janeiro in Brazil. Previously cultivated in green tea, the consortium was separately fermented 3 times in black tea and in coffee cascara tea infusion prior to experimental use in order to stabilize the microbial consortium in these matrixes [46]. All kombucha beverages were prepared according to the protocol described by Nummer [47];
- Black tea kombucha (BT K): Black tea kombucha was prepared by mixing 10% (volume/volume—v/v) of black tea starter, 80% black tea infusion (weight/volume—w/v), 10% (w/v) sugar, and 2.5% (v/v) of a Symbiotic Culture of Bacteria and Yeast (SCOBY) and letting the mixture ferment for 14 days at 23 °C (Sanyo™ MIR-154PE, Sanyo Electric Co., Ltd., Osaka, Japan). Samples were collected before fermentation (day 0) and after 3, 6, and 9 days of fermentation;
- Coffee cascara kombucha (CCB K and CCN K): Coffee cascara kombuchas were prepared using 80% (v/v) of the coffee cascara infusion, 10% (v/v) of the black tea kombucha, 10% (w/v) sugar, and 2.5% (w/v) of SCOBY. The mixture was allowed to ferment at 23 °C (Sanyo™ MIR-154PE, Sanyo Electric Co., Ltd., Japan). Samples were collected before fermentation (day 0) and after 3, 6, and 9 days of fermentation.
2.4. DNA Extraction, Amplicon Sequencing Data Analysis and Library Preparation
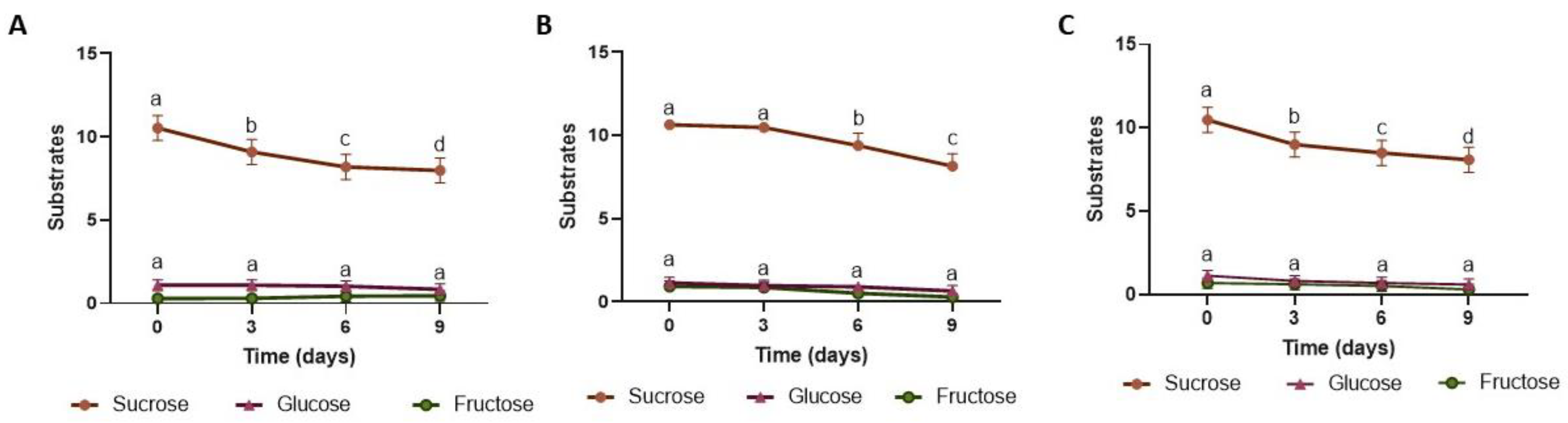
2.5. Analysis of Titratable Acidity, pH, Soluble Solids, and Sugars
2.6. Analysis of Bioactive Compounds
2.6.1. Analysis of Catechins, Chlorogenic Acids and Free Phenolic Acids
2.6.2. Analysis of Caffeine
2.7. Cell Viability Assay
2.8. Indoxyl Sulfate-Induced Oxidative Stress in HK-2 Diabetic Nephropathy Cell Model
2.9. Lipopolysaccharide-Induced Inflammation in RAW 264.7 Macrophages
2.10. Statistics
3. Results and Discussion
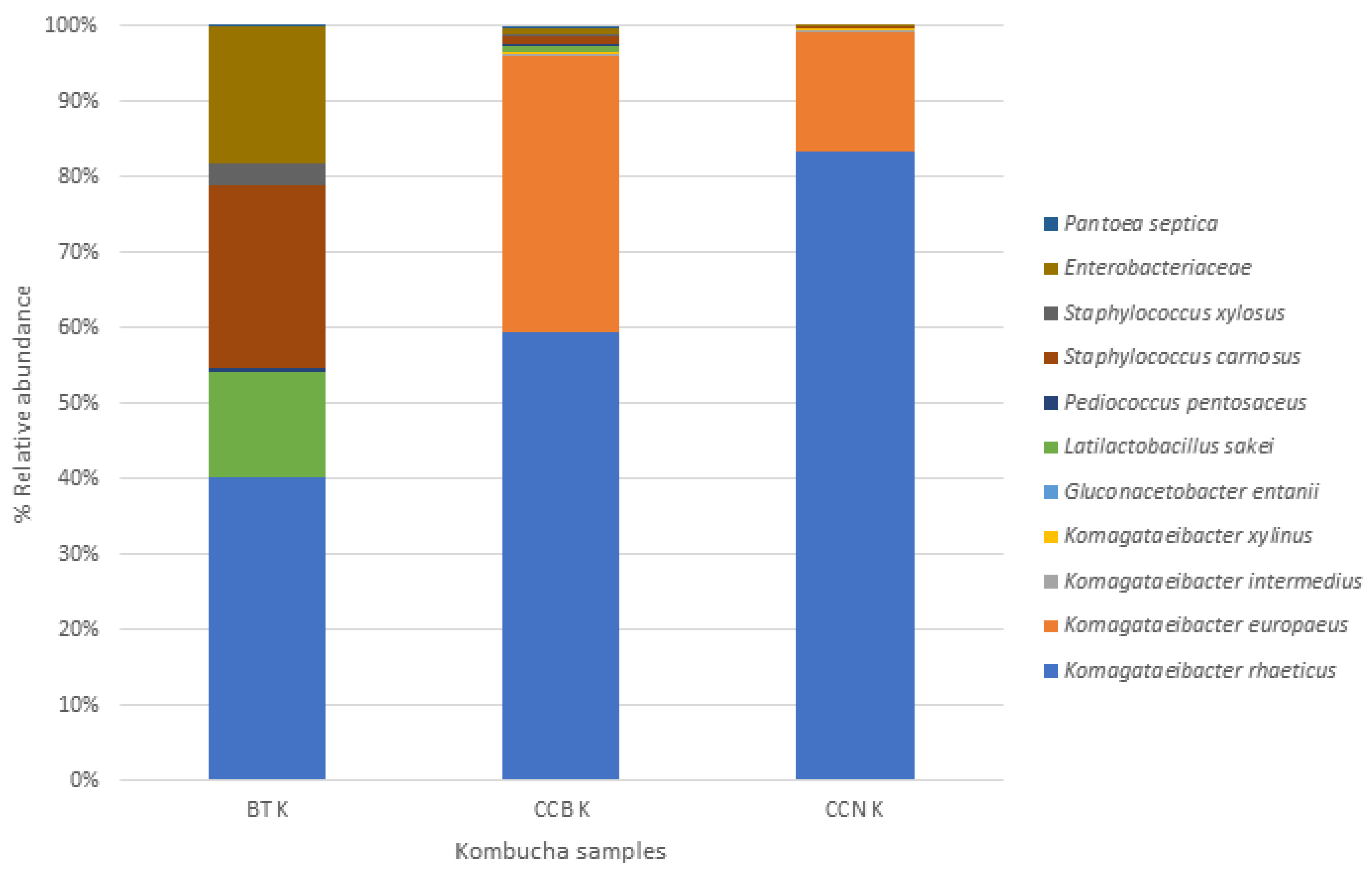
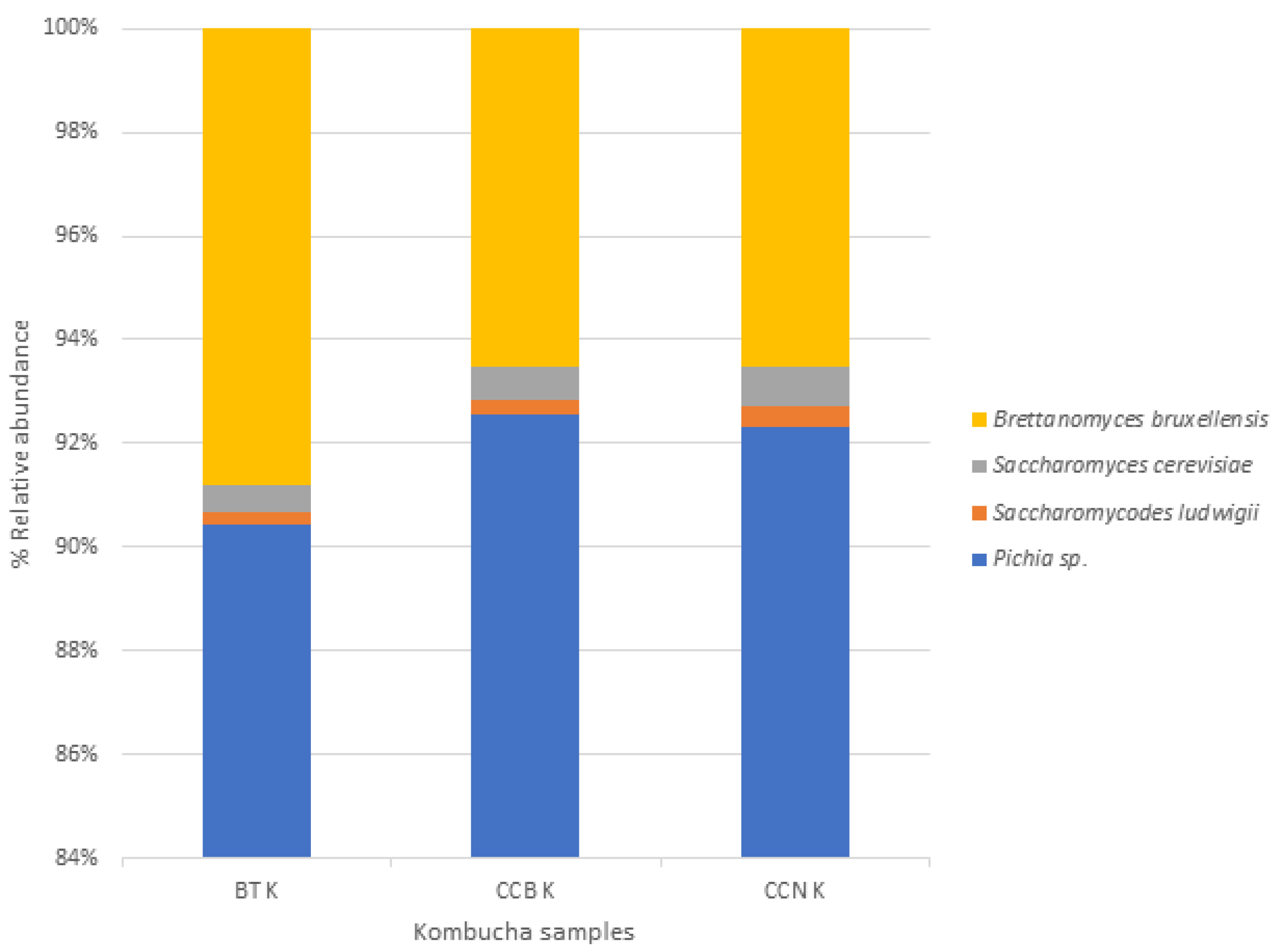
3.1. Microbial Taxonomy
3.2. Beverages Characterization
3.2.1. pH, Total Acidity, Soluble Solids, and Sugars
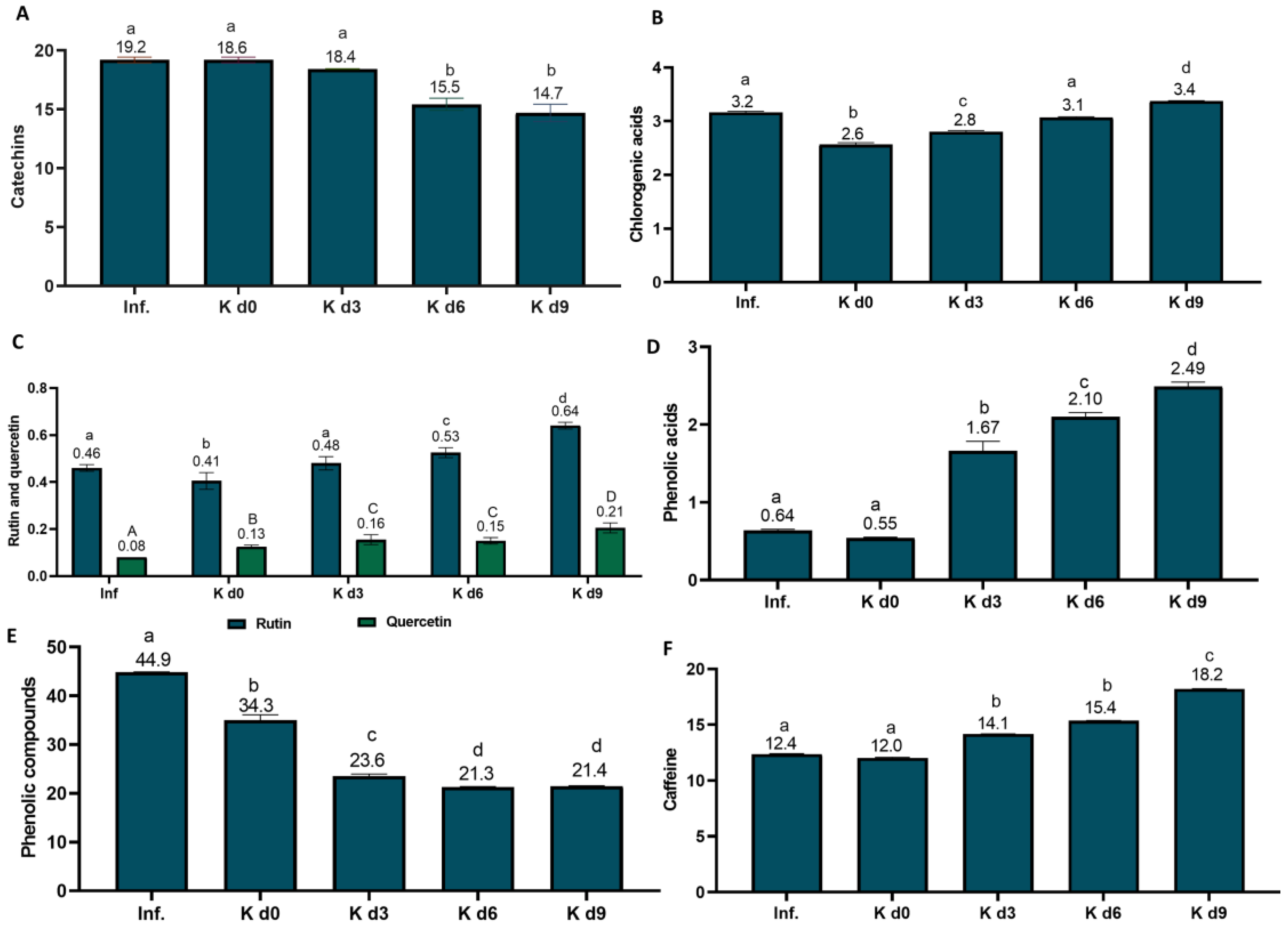
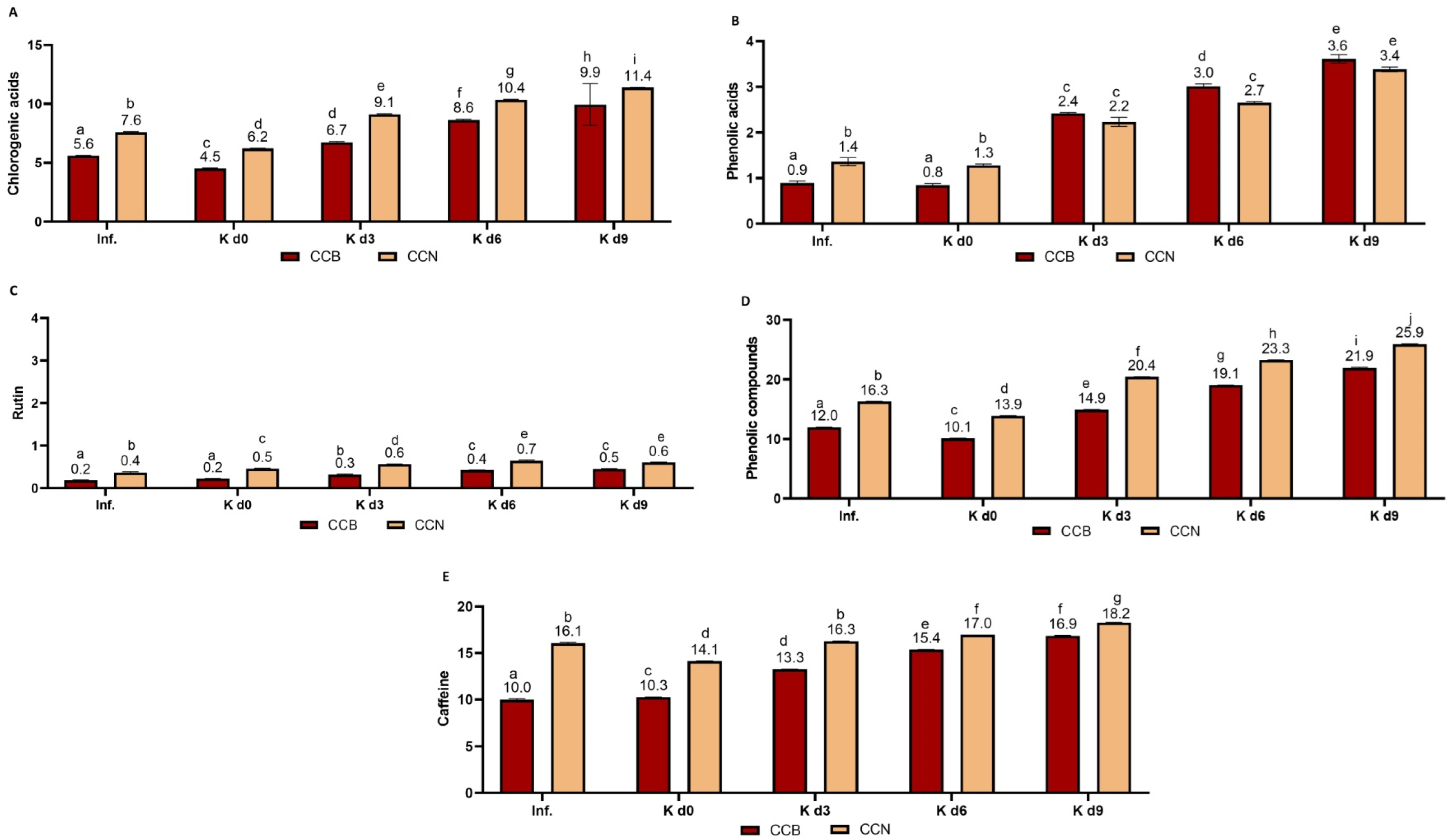
3.2.2. Bioactive Compounds
Black Tea
Coffee Cascara
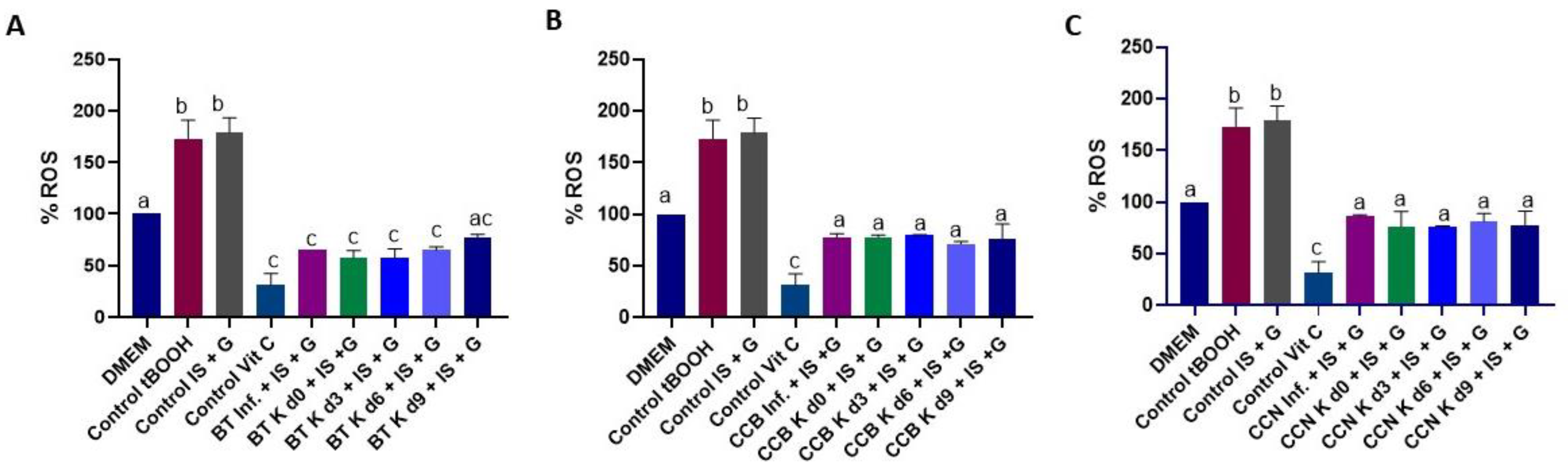
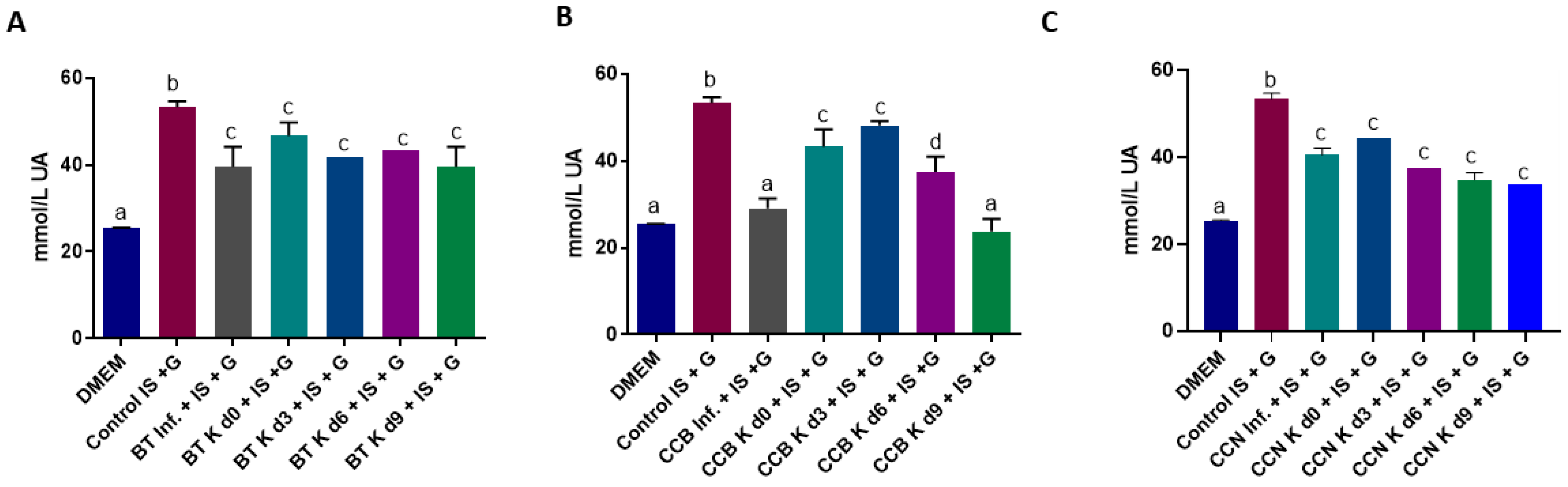
3.3. Potential Antioxidant Effect of Black Tea and Coffee Cascara Beverages in Indoxyl Sulfate-Induced HK-2 Cells
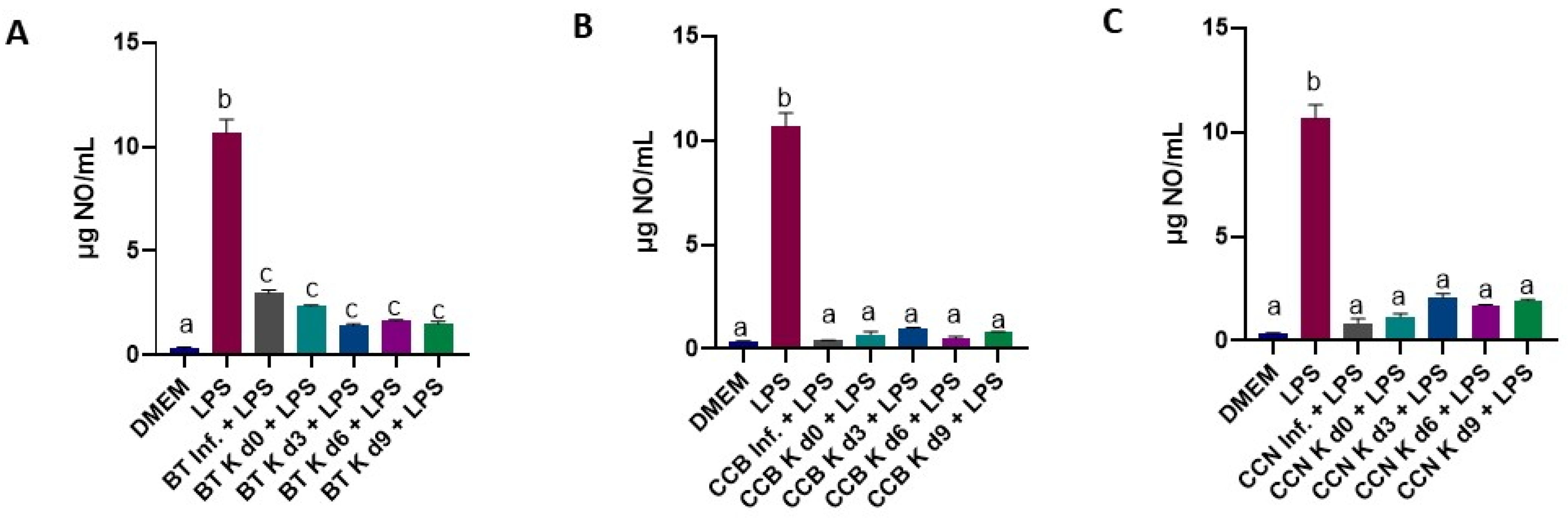
3.4. Effect of Black Tea and Coffee Cascara Beverages on Anti-Inflammatory Activity in Lipopolysaccharide-Induced RAW 264.7 Cell Line
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, M.-Z.; Lu, D.-M.; Ouyang, J.; Zhou, F.; Huang, P.-F.; Liu, Z.-H. Tea consumption and colorectal cancer risk: A meta-analysis of prospective cohort studies. Eur. J. Nutr. 2020, 59, 3603–3615. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Zhao, N.; Wang, D.; Shams-White, M.; Karlsen, M.; Cassidy, A.; Wallace, T.C. Dose-response relation between tea consumption and risk of cardiovascular disease and all-cause mortality: A systematic review and meta-analysis of population-based studies. Adv. Nutr. 2020, 11, 790–814. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-S.; Wang, W.-Y.; Fan, W.-Y.; Deng, Q.; Wang, X. Tea consumption and risk of type 2 diabetes: A dose–response meta-analysis of cohort studies. Br. J. Nutr. 2014, 111, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Arab, L.; Khan, F.; Lam, H. Tea consumption and cardiovascular disease risk. Am. J. Clin. Nutr. 2013, 98, 1651S–1659S. [Google Scholar] [CrossRef]
- Aboulwafa, M.M.; Youssef, F.S.; Gad, H.A.; Altyar, A.E.; Al-Azizi, M.M.; Ashour, M.L. A comprehensive insight on the health benefits and phytoconstituents of Camellia sinensis and recent approaches for its quality control. Antioxidants 2019, 8, 455. [Google Scholar] [CrossRef]
- Tang, G.-Y.; Meng, X.; Gan, R.-Y.; Zhao, C.-N.; Liu, Q.; Feng, Y.-B.; Li, H.-B. Health functions and related molecular mechanims of tea components: An update review. Int. J. Mol. Sci. 2019, 20, 6196. [Google Scholar] [CrossRef]
- Martínez-Leal, J.; Suárez, L.V.; Jayabalan, R.; Oros, J.H.; Escalante-Aburto, A. A review on health benefits of kombucha nutritional compounds and metabolites. CyTA-J. Food 2018, 16, 390–399. [Google Scholar] [CrossRef]
- Coelho, R.M.D.; de Almeida, A.L.; do Amaral, R.Q.G.; da Mota, R.N.; de Sousa, P.H.M. Kombucha: Review. Int. J.Gastron. Food Sci. 2020, 22, 100272. [Google Scholar] [CrossRef]
- Diez-Ozaeta, I.; Astiazaram, O.J. Recent advances in Kombucha tea: Microbial consortium, chemical parameters, health implications and biocellulose production. Int. J. Food Microbiol. 2022, 377, 109783. [Google Scholar] [CrossRef]
- Greenwalt, C.J.; Ledford, R.A.; Steinkraus, K.H. Determination and characterization of the antimicrobial activity of the fermented tea kombucha. LWT—Food Sci. Technol. 1998, 31, 291–296. [Google Scholar] [CrossRef]
- Jayabalan, R.; Subathradevi, P.; Marimuthu, S.; Sathishkumar, M.; Swaminathan, K. Changes in free-radical scavenging ability of kombucha tea during fermentation. Food Chem. 2008, 109, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Gachhui, R.; Sil, P.C. Hepatoprotective properties of kombucha tea against TBHP-induced oxidative stress via suppression of mitochondria dependent apoptosis. Pathophysiology 2011, 18, 221. [Google Scholar] [CrossRef] [PubMed]
- Srihari, T.; Kathikesan, K.; Ashokkumar, N.; Satyanarayana, U. Antihyperglycaemic efficacy of kombucha in streptozotocin-induced rats. J. Funct. Foods 2013, 5, 1794–1802. [Google Scholar] [CrossRef]
- Rahmani, R.; Beaufort, S.; Villarreal-Soto, S.A.; Taillandier, P.; Bouajila, J.; Debouba, M. Kombucha fermentation of African mustard (Brassica tournefortii) leaves: Chemical composition and bioactivity. Food Biosci. 2019, 30, 100414. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Renard, T.; Rollan, S.; Taillandier, P. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process. Biochem. 2019, 83, 44–54. [Google Scholar] [CrossRef]
- Kaewkod, T.; Bovonsombut, S.; Tragoolpua, Y. Efficacy of kombucha obtained from green, oolong, and black teas on tnhibition of pathogenic bacteria, antioxidation and toxicity on colorectal cancer cell line. Microorganisms 2019, 7, 700. [Google Scholar] [CrossRef]
- Cardoso, R.R.; Neto, R.O.; D’Almeida, C.T.S.; do Nascimento, T.P.; Pressete, C.G.; Azevedo, L.; Martino, H.S.D.; Cameron, L.C.; Ferreira, M.S.L.; de Barros, F.A.R. Kombuchas from green and black teas have different phenolic profile, which impacts their antioxidant capacities, antibacterial and antiproliferative activities. Food Res. Int. 2020, 128, 108782. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef]
- Mukherjee, A.; Gómez-Sala, B.; O’Connor, E.; Kenny, J.G.; Cotter, P.D. Global regulatory frameworks for fermented foods: A review. Front. Nutr. 2022, 9, 902642. [Google Scholar] [CrossRef]
- Grand View Research. Kombucha Market Size, Share & Trends Analysis Report by Product (Conventional, Hard), By Distribution Channel (On-Trade, Off-Trade), By Region, and Segment Forecasts, 2022–2030. Available online: https://www.grandviewresearch.com/industry-analysis/kombucha-market (accessed on 13 September 2022).
- Kim, J.; Adhikari, K. Current trends in kombucha: Marketing perspectives and the need for improved sensory research. Beverages 2020, 6, 15. [Google Scholar] [CrossRef]
- Leonarski, E.; Guimarães, A.C.; Cesca, K.; Poletto, P. Production process and characteristics of kombucha fermented from alternative raw materials. Food Biosci. 2022, 49, 101841. [Google Scholar] [CrossRef]
- Esquivel, P.; Jiménez, V.M. Functional properties of coffee and coffee by-products. Food Res. Int. 2012, 46, 488–495. [Google Scholar] [CrossRef]
- Coffee Production Worldwide from 2003/04 to 2020/21 (In Million 60 Kilogram Bags). Available online: https://www.statista.com/statistics/263311/worldwide-production-of-coffee/ (accessed on 16 December 2022).
- Coffee Production by Exporting Countries. Available online: https://www.ico.org/prices/po-production.pdf (accessed on 16 December 2022).
- DePaula, J.; Cunha, S.; Cruz, A.; Sales, A.L.; Revi, I.; Fernandes, J.; Ferreira, I.M.P.L.V.O.; Miguel, M.A.L.; Farah, A. Volatile fingerprinting and sensory profiles of coffee cascara teas produced in Latin American countries. Foods 2022, 11, 3144. [Google Scholar] [CrossRef] [PubMed]
- Iriondo-DeHond, A.; García, N.A.; Fernandez-Gomez, B.; Guisantes-Batan, E.; Escobar, F.V.; Blanch, G.P.; San Andres, M.I.; Sanchez-Fortun, S.; del Castillo, M.D. Validation of coffee by-products as novel food ingredients. Innov. Food Sci. Emerg. Technol. 2019, 51, 194–204. [Google Scholar] [CrossRef]
- Murthy, P.S.; Naidu, M.M. Recovery of phenolic antioxidants and functional compounds from coffee industry by-products. Food Bioprocess. Technol. 2012, 5, 897–903. [Google Scholar] [CrossRef]
- Janissen, B.; Huynh, T. Chemical composition and value-adding applications of coffee industry byproducts: A review. Resour., Conserv. Recycl. 2018, 128, 110–117. [Google Scholar] [CrossRef]
- Prata, E.R.B.A.; Oliveira, L.S. Fresh coffee husks as potential sources of anthocyanins. LWT 2007, 40, 1555–1560. [Google Scholar] [CrossRef]
- Rios, M.B.; Iriondo-DeHond, A.; Iriondo-DeHond, M.; Herrera, T.; Velasco, D.; Gómez-Alonso, S.; Callejo, M.J.; del Castillo, M.D. Effect of coffee cascara dietary fiber on the physicochemical, nutritional and sensory properties of a gluten-free bread formulation. Molecules 2020, 25, 1358. [Google Scholar] [CrossRef]
- Bondam, A.F.; da Silveira, D.D.; dos Santos, J.P.; Hoffmann, J.F. Phenolic compounds from coffee by-products: Extraction and application in the food and pharmaceutical industries. Trends Food Sci. Technol. 2022, 123, 172–186. [Google Scholar] [CrossRef]
- Bobková, A.; Poláková, K.; Demianová, A.; Belej, L.; Bobko, M.; Jurcaga, L.; Gálik, B.; Novotná, I.; Iriondo-DeHond, A.; del Castillo, M.D. Comparative analysis of selected chemical parameters of Coffea arabica, from Cascara to Silverskin. Nutrients 2022, 11, 1082. [Google Scholar] [CrossRef]
- Iriondo-DeHond, A.; Iriondo-DeHond, M.; del Castillo, M.D. Applications of compounds from coffee processing by-products. Biomolecules 2020, 10, 1219. [Google Scholar] [CrossRef] [PubMed]
- Duangjai, A.; Suphrom, N.; Wungrath, J.; Ontawong, A.; Nuengchamnong, N.; Yosboonruang, A. Comparison of antioxidant, antimicrobial activities and chemical profiles of three coffee (Coffea arabica L.) pulp aqueous extracts. Integr. Med. Res. 2016, 5, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Heeger, A.; Kosinska-Cagnazzo, A.; Cantergiani, E.; Andlauer, W. Bioactives of coffee cherry pulp and its utilisation for production of Cascara beverage. Food Chem. 2017, 221, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Magoni, C.; Bruni, I.; Guzzetti, L.; Dell’Agli, M.; Sangiovanni, E.; Pizza, S.; Regonesi, M.E.; Maldini, M.; Spezzano, R.; Caruso, D.; et al. Valorizing coffee pulp by-products as anti-inflammatory ingredient of food supplements acting on IL-8 release. Food Res. Int. 2018, 112, 129–135. [Google Scholar] [CrossRef]
- Khochapong, W.; Ketnawa, S.; Ogawa, Y.; Punbusayakul, N. Effect of in vitro digestion on bioactive compounds, antioxidant and antimicrobial activities of coffee (Coffea arabica L.) pulp aqueous extract. Food Chem. 2021, 348, 129094. [Google Scholar] [CrossRef]
- Del Castillo, M.D.; Fernandez-Gomez, B.; Martinez-Saez, N.; Iriondo-DeHond, A.; Mesa, M.D. Coffee by-products. In Coffee: Production, Quality and Chemistry, 1st ed.; Farah, A., Ed.; Royal Society of Chemistry: London, UK, 2019; Volume 1, pp. 309–336. [Google Scholar]
- EFSA NDA Panel (EFSA Panel on Nutrition, Novel Foods and Food Allergens). Safety of Dried Coffee Husk (Cascara) from Coffea arabica L. as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2022, 20, 7085. [Google Scholar]
- Iriondo-DeHond, M.; Iriondo-DeHond, A.; Herrera, T.; Fernández-Fernández, A.M.; Sorzano, C.O.S.; Miguel, E.; del Castillo, M.D. Sensory acceptance, appetite control and gastrointestinal tolerance of yogurts containing coffee-cascara extract and inulin. Nutrients 2020, 12, 627. [Google Scholar] [CrossRef]
- Einfalt, D.; Meissner, K.; Kurz, L.; Intami, K.; Müller, J. Fruit spirit production from coffee cherries—Process analysis and sensory evaluation. Beverages 2020, 6, 57. [Google Scholar] [CrossRef]
- Blumenthal, P.; Steger, M.C.; Bellucci, A.Q.; Segatz, V.; Rieke-Zapp, J.; Sommerfeld, K.; Schwarz, S.; Einfalt, D.; Lachenmeier, D.W. Prodution of coffee cherry spirits from Coffea arabica varieties. Foods 2022, 11, 1672. [Google Scholar] [CrossRef]
- Muzaifa, M.; Andini, R.; Sulaiman, M.I.; Abubakar, Y.; Rahmi, F.; Nurzainura. Novel utilization of coffee processing by-products: Kombucha cascara originated from ‘Gayo-Arabica’. IOP Conf. Ser. Earth Environ. Sci. 2021, 644, 012048. [Google Scholar] [CrossRef]
- Huynh-Ba, T. Preparation of Quinic Acid Derivatives. U.S. Patent 5401858, 28 March 1995. [Google Scholar]
- Villarreal-Soto, S.A.; Bouajila, J.; Pace, M.; Leech, J.; Cotter, P.D.; Souchard, J.-P.; Taillandier, P.; Beaufort, S. Metabolome-microbiome signatures in the fermented beverage, Kombucha. Int. J. Food Microbiol. 2020, 333, 108778. [Google Scholar] [CrossRef] [PubMed]
- Nummer, B.A. Kombucha brewing under the Food and Drug Administration model food code: Risk analysis and processing guidance. J. Environ. Health 2013, 76, 8–11. [Google Scholar] [PubMed]
- Yamanaka, L.E.; Abdala, P.; Christoff, A.P. Extração de DNA: Avaliação da Metodologia Utilizada pela Neoprospecta. Neoprospecta Microbiome Technologies. Nota Técnica: Extração de DNA Neo. 2018. Available online: http://neoprospecta.com (accessed on 26 August 2022).
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 20 March 2023).
- Smyth, R.P.; Schlub, T.E.; Grimm, A.; Venturi, V.; Chopra, A.; Mallal, S.; Davenport, M.P.; Mak, J. Reducing chimera formation during PCR amplification to ensure accurate genotyping. Gene 2010, 469, 45–51. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Instituto Adolfo Lutz. Métodos Físico-Químicos para Análise de Alimentos, 4th ed.; Instituto Adolfo Lutz: São Paulo, Brazil, 2008. Available online: http://www.ial.sp.gov.br/ial/publicacoes/livros/metodos-fisico-quimicos-para-analise-de-alimentos (accessed on 5 May 2023).
- Wischral, D.; Arias, J.M.; Modesto, L.F.; Passos, D.F.; Pereira, N., Jr. Lactic acid production from sugarcane bagasse hydrolysates by Lactobacillus pentosus: Integrating xylose and glucose fermentation. Biotechnol. Prog. 2019, 35, e2718. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Liang, Y.; Dong, J.; Lu, J. Tea extraction methods in relation to control of epimerization of tea catechins. J. Sci. Food Agric. 2007, 87, 1748–1752. [Google Scholar] [CrossRef]
- Farah, A.; de Paulis, T.; Moreira, D.P.; Trugo, L.C.; Martin, P.R. Chlorogenic acids and lactones in regular and water-decaffeinated arabica coffees. J. Agric. Food Chem. 2006, 53, 374–381. [Google Scholar] [CrossRef]
- Duarte, G.S.; Farah, A. Effect of simultaneous consumption of milk and coffee on chlorogenic acids’ bioavailability in humans. J. Agric. Food Chem. 2011, 59, 7925–7931. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.C.; Calado, V.; Franca, A.S.; Trugo, L.C. Correlation between cup quality and chemical attributes of Brazilian coffees. Food Chem. 2006, 98, 373–380. [Google Scholar] [CrossRef]
- Bakondi, E.; Gönczi, M.; Szabó, É.; Bai, P.; Pacher, P.; Gergely, P.; Virág, L. Role of intracellular calcium mobilization and cell-density-dependent signaling in oxidative-stress-induced cytotoxicity in HaCaT keratinocytes. J. Investig. Dermatol. 2003, 121, 88–95. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C. Fluorescence probes used for detection of reactive oxygen species. J. Biochem. Biophys. Methods 2005, 65, 45–80. [Google Scholar] [CrossRef] [PubMed]
- IriondoDeHond, A.; Abdolmaleky, H.M.; Zhou, J.R.; Del Castillo, M.D. Assesment of antioxidante and anti-inflammatory effects of coffee by-product extracts on a cellular model of diabetic nephropathy. Nutr. Clín. Diet. Hosp. 2018, 38 (Suppl. 1), 116–117. [Google Scholar]
- Benayad, Z.; Martinez-Villaluenga, C.; Frias, J.; Gomez-Cordoves, C.; Es-Safi, N.E. Phenolic Composition, Antioxidant and Anti-Inflammatory Activities of Extracts from Moroccan Opuntia Ficus-Indica Flowers Obtained by Different Extraction Methods. Ind. Crops Prod. 2014, 62, 412–420. [Google Scholar] [CrossRef]
- Bishop, P.; Pitts, E.R.; Budner, D.; Thompson-Witrick, K.A. Kombucha: Biochemical and microbiological impacts on the chemical and flavor profile. Food Chem. 2022, 1, 100025. [Google Scholar] [CrossRef]
- Arikan, M.; Mitchell, A.L.; Finn, R.D.; Gürel, F. Microbial composition of Kombucha determined using amplicon sequencing and shotgun metagenomics. J. Food Sci. 2020, 82, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Marsh, A.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef]
- Landis, E.A.; Fogarty, E.; Edwards, J.C.; Popa, O.; Eren, A.M.; Wolfe, B.E. Microbial diversity and interaction specificity in kombucha tea fermentations. mSystems 2022, 7, e0015722. [Google Scholar] [CrossRef]
- Lavasani, P.S.; Motevaseli, E.; Sanikhani, N.S.; Modarressi, M.H. Komagataeibacter xylinus as a novel probiotic candidate with high glucose conversion rate properties. Heliyon 2019, 5, e01571. [Google Scholar] [CrossRef]
- Liu, M.; Chen, Q.; Sun, Y.; Zeng, L.; Wu, H.; Gu, Q.; Li, P. Probiotic potential of a folate-producing strains LatilactobacillussakeiLZ217 and its modulation effects on human gut Microbiota. Foods 2022, 11, 234. [Google Scholar] [CrossRef]
- Qi, Y.; Huang, L.; Zeng, Y.; Li, W.; Zhou, D.; Xie, J.; Xie, J.; Tu, Q.; Deng, D.; Yin, J. Pediococcus pentosaceus: Screening and application as probiotic in food processing. Front. Mirobiol. 2021, 12, 762467. [Google Scholar] [CrossRef]
- Singh, N.; Anand, S. Enterobacteriaceae. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: London, UK, 2011; Update in 2020; pp. 67–71. [Google Scholar]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in alcoholic fermentation processes: Role of physiological fitness and microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- Staniszewski, A.; Kordowska-Wiater, M. Probiotic and potentially probiotic yeasts—Characteristics and food application. Foods 2021, 10, 1306. [Google Scholar] [CrossRef] [PubMed]
- Vejarano, R. Saccharomycodes ludwigii, control and potential uses in winemaking process. Fermentation 2018, 4, 71. [Google Scholar] [CrossRef]
- Teoh, A.L.; Heard, G.; Cox, J. Yeast ecology of Kombucha fermentation. Int. J. Food Microbiol. 2004, 95, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Júnior, J.C.; Magnani, M.; da Costa, W.K.A.; Madruga, M.S.; Olegário, L.S.; Borges, G.S.C.; Santas, A.M.; Lima, M.S.; de Lima, L.C.; Brito, I.L.; et al. Traditional and flavored kombuchas with pitanga and umbu-cajá pulps: Chemical properties, antioxidants, and bioactive compounds. Food Biosci. 2021, 44, 101380. [Google Scholar] [CrossRef]
- May, A.; Narayanan, S.; Alcock, J.; Varsani, A.; Maley, C.; Aktips, A. Kombucha: A novel model system for cooperation and conflict in a complex multi-species microbial ecosystem. PeerJ 2019, 7, e7565. [Google Scholar] [CrossRef]
- Jafari, R.; Naghavi, N.S.; Khosravi-Darani, K.; Doudi, M.; Shahanipour, K. Kombucha microbial starter with enhanced production of antioxidant compounds and invertase. Biocatal. Agric. Biotechnol. 2020, 29, 101789. [Google Scholar] [CrossRef]
- Jayabalan, R.; Marimuthu, S.; Swaminathan, K. Changes in content of organic acids and tea polyphenols during kombucha tea fermentation. Food Chem. 2007, 102, 392–398. [Google Scholar] [CrossRef]
- De Noronha, M.C.; Cardoso, R.R.; D’Almeida, C.T.S.; do Carmo, M.A.V.; Azevedo, L.; Maltarollo, V.G.; Ribeiro Júnior, J.I.; Eller, M.R.; Cameron, L.C.; Ferreira, M.S.L.; et al. Black tea kombucha: Physicochemical, microbiological and comprehensive phenolic profile changes during fermentation, and antimalarial activity. Food Chem. 2022, 384, 132515. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Get the Facts: Sugar-Sweetened Beverages and Consumption. Available online: https://www.cdc.gov/nutrition/data-statistics/sugar-sweetened-beverages-intake.html (accessed on 17 April 2023).
- Brazilian Institute of Geography and Statistics (IBGE). Pesquisa de Orçamentos Familiares 2017–2018: Análise do Consumo Alimentar Pessoal No Brasil, Coordenação de Trabalho e Rendimento; Brazilian Institute of Geography and Statistics: Rio de Janeiro, Brazil, 2020.
- Eurostat. How Often Do You Drink Sugar-Sweetened Soft Drinks? Available online: https://ec.europa.eu/eurostat/web/products-eurostat-news/-/ddn-20210727-1 (accessed on 17 April 2023).
- Koch, W.; Kukula-Koch, W.; Glowniak, K. Catechin composition and antioxidant activity of black teas in relation to brewing time. J. AOAC Int. 2017, 100, 1694–1699. [Google Scholar] [CrossRef]
- Gaggia, F.; Baffoni, L.; Galiano, M.; Nielsen, D.S.; Jakobsen, R.R.; Castro-Mejía, J.L.; Bosi, S.; Truzzi, F.; Musumeci, F.; Dinelli, G.; et al. Kombucha beverage from green, black and rooibos teas: A comparative study looking at microbiology, chemistry and antioxidant activity. Nutrients 2019, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of dietary flavonoids and phenolic compounds. Mol. Aspects Med. 2010, 31, 446–467. [Google Scholar] [CrossRef] [PubMed]
- Alberto, M.R.; Gómez-Cordovés, C.; de Nadra, M.C.M. Metabolism of gallic acid and catechin by Lactobacillus hilgardii from wine. J. Agric. Food Chem. 2004, 52, 6465–6469. [Google Scholar] [PubMed]
- Farah, A.; Lima, J.P. Consumption of chlorogenic acids through coffee and health implications. Beverages 2019, 5, 11. [Google Scholar] [CrossRef]
- Li, C.; Lee, M.-J.; Sheng, S.; Meng, X.; Prabhu, S.; Winnik, B.; Huang, B.; Chung, J.Y.; Yan, S.; Ho, C.-T.; et al. Structural identification of two metabolites of catechins and their kinetics in human urine and blood after tea ingestion. Chem. Res. Toxicol. 2000, 13, 177–184. [Google Scholar] [CrossRef]
- Stoikidou, T.; Koidis, A. Chapter 2-Coffee and tea bioactive compounds. In Functional Foods and Their Implications for Health Promotion, 1st ed.; Zabetakis, I., Tsoupras, A., Lordan, R., Ramji, D., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2023; pp. 29–53. [Google Scholar]
- Farah, A.; Donangelo, C.M. Phenolic compounds in coffee. Braz. J. Plant. Physiol. 2006, 18, 23–36. [Google Scholar] [CrossRef]
- Pereira, G.V.M.; Carvalho Neto, D.P.; Magalhães Júnior, A.I.; do Prado, F.G.; Pagnoncelli, M.G.B.; Karp, S.G.; Soccol, C.R. Chemical composition and health properties of coffee and coffee by-products. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 91, pp. 65–96. [Google Scholar]
- Plumb, G.W.; Garcia-Conesa, M.T.; Kroon, P.A.; Rhodes, M.; Ridley, S.; Williamson, G. Metabolism of chlorogenic acid by human plasma, liver, intestine and gut microflora. J. Sci. Food Agric. 1999, 79, 390–392. [Google Scholar] [CrossRef]
- da Silveira, J.S.; Durand, N.; Lacour, S.; Belleville, M.-P.; Perez, A.; Loiseau, G.; Dornier, M. Solid-state fermentation as a sustainable method for coffee pulp treatment and production of an extract rich in chlorogenic acids. Food Bioprod. Process. 2019, 115, 175–184. [Google Scholar] [CrossRef]
- Riva, A.; Kolimár, C.; Spittler, A.; Wisgrill, L.; Herbold, C.W.; Abrankó, L.; Berry, D. Conversion of rutin, a prevalent dietary flavonol, by the human gut microbiota. Front. Microbiol. 2020, 11, 585428. [Google Scholar] [CrossRef]
- Ramirez-Martinez, J.R. Phenolic compounds in coffee pulp: Quantitative determination by HPLC. J. Sci. Food Agric. 1988, 43, 135–144. [Google Scholar] [CrossRef]
- Mullen, W.; Nemzer, B.; Stalmach, A.; Ali, S.; Combet, E. Polyphenolic and hydroxycinnamate contents of whole coffee fruits from China, India and Mexico. J. Agric. Food Chem. 2013, 61, 5298–5309. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T. Indoxyl sulfate is a nephro-vascular toxin. J. Renal Nutr. 2010, 20, S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.-H.; Ma, M.-C.; Liao, M.-T.; Zheng, C.M.; Lu, K.C.; Liao, C.H.; Hou, Y.-C.; Liu, W.-C.; Lu, C.-L. Indoxyl sulfate, a tubular toxin, contributes to the development of chronic kidney disease. Toxins 2020, 12, 684. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Small, D.M.; Vesey, D.A.; Johnson, D.W.; Francis, R.; Vitetta, L.; Gobe, G.C.; Morais, C. Indoxyl sulfate and kidney disease: Causes, consequences and interventions. Nephrology 2016, 21, 170–177. [Google Scholar] [CrossRef]
- Ellis, R.J.; Small, D.M.; Ng, K.L.; Vesey, D.A.; Vitetta, L.; Francis, R.S.; Gobe, G.C.; Morais, C. Indoxyl sulfate induces apoptosis and hypertrophy in human kidney proximal tubular cells. Toxicol. Pathol. 2018, 4, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Stopper, H.; Zegura, B.; Dusinska, M.; Moller, P. Do cytotoxicity and cell death cause false positive results in the in vitro comet assay? Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2022, 881, 503520. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Villegas, A.; Sánchez-Tapia, M.; Avila-Nava, A.; Ramírez, V.; Tovar, A.R.; Torres, N. Differential effect of sucrose and fructose in combination with a high fat diet on intestinal microbiota and kidney oxidative stress. Nutrients 2017, 9, 393. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Gachhui, R.; Sil, P.C. Effect of Kombucha, a fermented black tea in attenuating oxidative stress mediated tissue damage in alloxan induced diabetic rats. Food Chem. Toxicol. 2013, 60, 328–340. [Google Scholar] [CrossRef]
- Sun, X.-Y.; Wang, J.M.; Ouyang, J.-M.; Kuang, L. Antioxidant activities and repair effects on oxidatively damaged HK-2 cells of tea polysaccharides with different molecular weights. Oxid. Med. Cell. Longev. 2018, 21, 5297539. [Google Scholar] [CrossRef]
- Visioli, F.; De La Lastra, C.A.; Andres-Lacueva, C.; Aviram, M.; Calhau, C.; Cassano, A.; D’Archivo, M.; Faria, A.; Favé, G.; Fogliano, V.; et al. Polyphenols and human health: A prospectus. Crit. Rev. Food Sci. Nutr. 2011, 51, 524–546. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxid. Med. Cell. Longev. 2016, 2016, 7432979. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The role of catechins in cellular responses to oxidative stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; de Mejia, E.G. Relationship of the phytochemicals from coffee and cocoa by-products with their potential to modulate biomarkers of metabolic syndrome in vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; de Mejia, E.G. Phenolic compounds from coffee by-products modulate adipogenesis-related inflammation, mitochondrial dysfunction, and insulin resistance in adipocytes, via insulin/PI3K/AKT signaling pathways. Food Chem. Toxicol. 2019, 132, 110672. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Clifford, M.N.; Lean, M.E.J.; Ashihara, H.; Crozier, A. Coffee: Biochemistry and potential impact on health. Food Funct. 2014, 5, 1695–1717. [Google Scholar] [CrossRef]
- Lima, J.P.; Farah, A. Caffeine metabolism and health effects. In Coffee: Consumption and Health Implications, 1st ed.; Farah, A., Ed.; Royal Society of Chemistry: London, UK, 2019; Volume 2, pp. 340–363. [Google Scholar]
- Chaudhary, K.; Malhota, K.; Sowers, J.; Arror, A. Uric Acid—Key ingredient in the recipe for cardiorenal metabolic syndrome. Cardiorenal Med. 2013, 3, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Kocak, M.Z.; Aktas, G.; Duman, T.T.; Atak, B.M.; Savli, H. Is Uric Acid elevation a random finding or a causative agent of diabetic nephropathy? Rev. Assoc. Med. Bras. 2019, 65, 1156–1160. [Google Scholar] [CrossRef]
- Khan, N.U.; Lin, J.; Liu, X.; Li, H.; Lu, W.; Zong, Z.; Zhang, H.; Waqas, M.; Shen, L. Insights into predicting diabetic nephropathy using urinary biomarkers. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140475. [Google Scholar] [CrossRef]
- Mahmoodpoor, F.; Saadat, Y.R.; Barzegari, A.; Ardalan, M.; Vahed, S.Z. The impact of gut microbiota on kidney function and pathogenesis. Biomed. Pharmacother. 2017, 93, 412–419. [Google Scholar] [CrossRef]
- Hou, C.; Liu, D.; Wang, M.; Gong, C.; Li, Y.; Yang, L.; Yao, M.; Yuan, E.; Ren, J. Novel xanthine oxidase-based cell model using HK-2 cell for screening antihyperuricemic functional compounds. Free. Radic. Biol. Med. 2019, 136, 135–145. [Google Scholar] [CrossRef]
- Zhu, H.; Song, D.; Zhao, X. Potential applications and preliminary mechanism of action of dietary polyphenols against hyperuricemia: A review. Food Biosci. 2021, 43, 101297. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, B.; Zhao, X.; Lin, Y.; Wang, J.; Wang, X.; Hu, N.; Wang, S. Chlorogenic acid supplementation ameliorates hyperuricemia, relieves renal inflammation, and modulates intestinal homeostasis. Food Funct. 2021, 12, 5637–5649. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Jiang, C.; Liu, Q.; Yn, H.; Zhou, H.; Ma, H.; Geng, Q. The inverted U-Shaped association of caffeine intake with serum uric acid in U.S. adults. J. Nutr. Health Aging 2022, 26, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-Q.; Fan, X.H.; Li, J.-F.; Chen, J.-H.; Liang, Y.; Hu, X.L.; Ma, S.M.; Hao, X.-Y.; Shi, T.; Wang, Z. Discovery of a novel inhibitor of nitric oxide production with potential therapeutic effect on acute inflammation. Bioorg. Med. Chem. Lett. 2021, 44, 128106. [Google Scholar] [CrossRef]
- Moghaddam, A.S.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef]
- Lim, A.K.H.; Tesch, G.H. Inflammation in diabetic nephropathy. World J. Diabetes 2014, 5, 393–398. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, Y.; Hu, W.; Ren, L.; Jiang, P.; Margolskee, R.F.; Wang, H.; Feng, S. Lipopolysaccharide-induced inflammation increases nitric oxide production in taste buds. Brain Behav. Immun. 2022, 103, 145–153. [Google Scholar] [CrossRef]
- Page, M.J.; Kell, D.B.; Preterius, E. The role of lipopolysaccharide-induces cell signalling in chronic inflammation. Chronic Stress. 2022, 6, 24705470221076390. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-S.; Wu, S.-S.; Lin, J.-K. Determination of tea polyphenols and caffeine in tea flowers (Camellia sinensis) and their hydroxyl radical scavenging and nitric oxide suppressing effects. J. Agric. Food Chem. 2003, 51, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Novilla, A.; Djamhuri, D.S.; Nurhayati, B.; Rihibiha, D.D.; Afifah, E.; Widowati, W. Anti-inflammatory properties of oolong tea (Camellia sinensis) ethanol extract and epigallocatechin gallate in LPS-induced RAW 264.7 cells. Asian Pac. J. Trop. Biomed. 2017, 7, 1005–1009. [Google Scholar] [CrossRef]
- Wang, P.; Feng, Z.; Sang, X.; Chen, W.; Zhang, X.; Xiao, J.; Chen, Y.; Chen, Q.; Yang, M.; Du, J. Kombucha ameliorates LPS-induced sepsis in a mouse model. Food Funct. 2021, 12, 10263–10280. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Cabral, B.D.; Larrosa-Pérez, M.; Gallegos-Infante, J.A.; Moreno-Jiménez, M.R.; González-Laredo, R.F.; Rutiaga-Quiñones, J.G.; Gamboa-Gómez, C.I.; Rocha-Guzmán, N.E. Oak kombucha protects against oxidative stress and inflammatory processes. Chem.-Biol. Interact. 2017, 272, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Diao, P.; Shu, X.; Li, L.; Xiong, L. Quercetin and quercitrin attenuates the inflammatory response and oxidative stress in LPS-induced RAW 264.7 cells: In vitro assessment and a theoretical model. Biomed. Res. Int. 2019, 28, 7039802. [Google Scholar]
- Lin, H.-Y.; Juan, S.-H.; Shen, S.-C.; Hsu, F.L.; Chen, Y.-C. Inhibition of lipopolysaccharide-induced nitric oxide production by flavonoids in RAW 264.7 macrophages involves heme oxygenase-1. Biochem. Pharmacol. 2003, 9, 1821–1832. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.; Aisa, H.A. Anti-inflammatory effect of pomegranate flower in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages. Pharm. Biol. 2017, 55, 2095–2101. [Google Scholar] [CrossRef]
- Gu, I.; Brownmiller, C.; Stebbind, N.B.; Mauromoustakos, A.; Howard, L.; Lee, S.-O. Berry phenolic and volatile extracts inhibit pro-inflammatory cytokine secretion in LPS—Stimulated RAW 264.7 cells throught suppression of NK-kB signaling pathway. Antioxidants 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, A.M.; Dellacassa, E.; Nardin, T.; Larcher, R.; Gámbaro, A.; Medrano-Fernandez, A.; del Castillo, M.D. In vitro bioaccessibility of bioactive compounds from citrus pomaces and orange pomace biscuits. Molecules 2021, 26, 3480. [Google Scholar] [CrossRef]
- Gutierrez-Barrutia, M.B.; Cozzano, S.; Arcia, P.; del Castillo, M.D. In vitro digestibility and bioaccessibility of nutrients and non-nutrients composing extruded brewers’ spent grain. Nutrients 2022, 14, 3480. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Kim, K.J.; Ryu, S.J.; Lee, B.-Y. Caffeine prevents LPS-induced inflammatory responses in RAW 264.7 cells and zebrafish. Chem. -Biol. Interact. 2016, 248, 1–7. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Days of Fermentation | Titratable Acidity (mEq/L) | pH | Soluble Solids (°Brix) |
|---|---|---|---|---|
| Black tea | Infusion | 0.02 ± 0.01 a | 5.5 ± 0.01 a | 1.1 ± 0.12 a |
| 0 | 0.1 ± 0.00 b | 3.8 ± 0.07 b | 10.4 ± 0.07 b | |
| 3 | 0.2 ± 0.05 b | 3.5 ± 0.00 b | 10.8 ± 0.14 b | |
| 6 | 0.3 ± 0.06 b | 3.5 ± 0.00 b | 9.7 ± 0.00 c | |
| 9 | 0.3 ± 0.06 b | 3.4 ± 0.00 b | 9.3 ± 0.28 c | |
| CCB | Infusion | 0.04 ± 0.00 a | 4.4 ± 0.05 a | 1.3 ± 0.12 a |
| 0 | 0.1 ± 0.00 a | 3.7 ± 0.07 b | 11.5 ± 0.63 b | |
| 3 | 0.2 ± 0.00 b | 3.6 ± 0.07 b | 11.3 ± 0.00 b | |
| 6 | 0.2 ± 0.00 b | 3.6 ± 0.07 b | 10.4 ± 0.14 c | |
| 9 | 0.2 ± 0.00 b | 3.5 ± 0.00 b | 10.0 ± 0.42 c | |
| CCN | Infusion | 0.04 ± 0.00 a | 4.3 ± 0.15 a | 1.2 ± 0.15 a |
| 0 | 0.04 ± 0.00 a | 3.8 ± 0.21 b | 11.6 ± 0.14 b | |
| 3 | 0.2 ± 0.00 b | 3.6 ± 0.07 b | 10.9 ± 0.14 b | |
| 6 | 0.2 ± 0.05 b | 3.5 ± 0.00 b | 10.6 ± 0.49 b | |
| 9 | 0.4 ± 0.08 b | 3.5 ± 0.00 b | 9.9 ± 0.56 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sales, A.L.; Iriondo-DeHond, A.; DePaula, J.; Ribeiro, M.; Ferreira, I.M.P.L.V.O.; Miguel, M.A.L.; del Castillo, M.D.; Farah, A. Intracellular Antioxidant and Anti-Inflammatory Effects and Bioactive Profiles of Coffee Cascara and Black Tea Kombucha Beverages. Foods 2023, 12, 1905. https://doi.org/10.3390/foods12091905
Sales AL, Iriondo-DeHond A, DePaula J, Ribeiro M, Ferreira IMPLVO, Miguel MAL, del Castillo MD, Farah A. Intracellular Antioxidant and Anti-Inflammatory Effects and Bioactive Profiles of Coffee Cascara and Black Tea Kombucha Beverages. Foods. 2023; 12(9):1905. https://doi.org/10.3390/foods12091905
Chicago/Turabian StyleSales, Amanda L., Amaia Iriondo-DeHond, Juliana DePaula, Mafalda Ribeiro, Isabel M. P. L. V. O. Ferreira, Marco Antonio L. Miguel, María Dolores del Castillo, and Adriana Farah. 2023. "Intracellular Antioxidant and Anti-Inflammatory Effects and Bioactive Profiles of Coffee Cascara and Black Tea Kombucha Beverages" Foods 12, no. 9: 1905. https://doi.org/10.3390/foods12091905
APA StyleSales, A. L., Iriondo-DeHond, A., DePaula, J., Ribeiro, M., Ferreira, I. M. P. L. V. O., Miguel, M. A. L., del Castillo, M. D., & Farah, A. (2023). Intracellular Antioxidant and Anti-Inflammatory Effects and Bioactive Profiles of Coffee Cascara and Black Tea Kombucha Beverages. Foods, 12(9), 1905. https://doi.org/10.3390/foods12091905