

The Behavior of Phenolic Compounds from Apples during Simulated Gastrointestinal Digestion with Focus on Chlorogenic Acid

Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards, Chemicals, and Solutions
2.2. Apple Samples and the Extraction of Phenolic Compounds
2.3. Simulated Digestion
2.4. Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC)
2.5. Degradation of Chlorogenic Acids in Simulated Digestion Electrolyte Solutions (SSF, SGF, SIF)
2.6. Statistical Analysis
3. Results
3.1. Phenolic Compounds in Apples
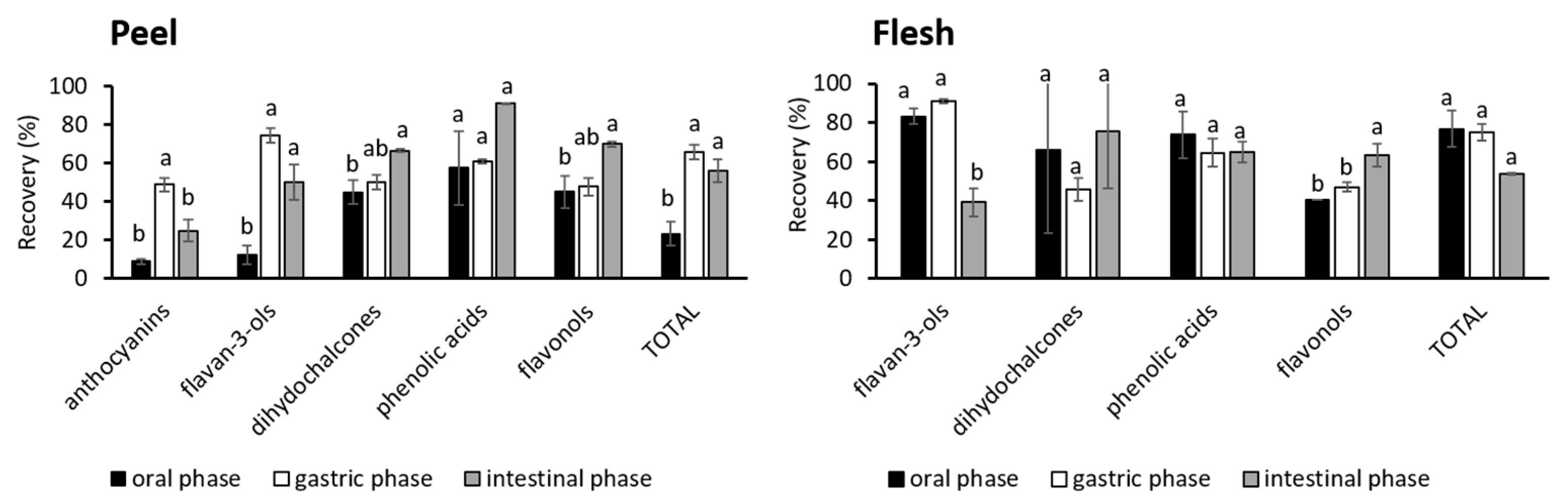
3.2. Phenolic Compounds in Simulated Digestion
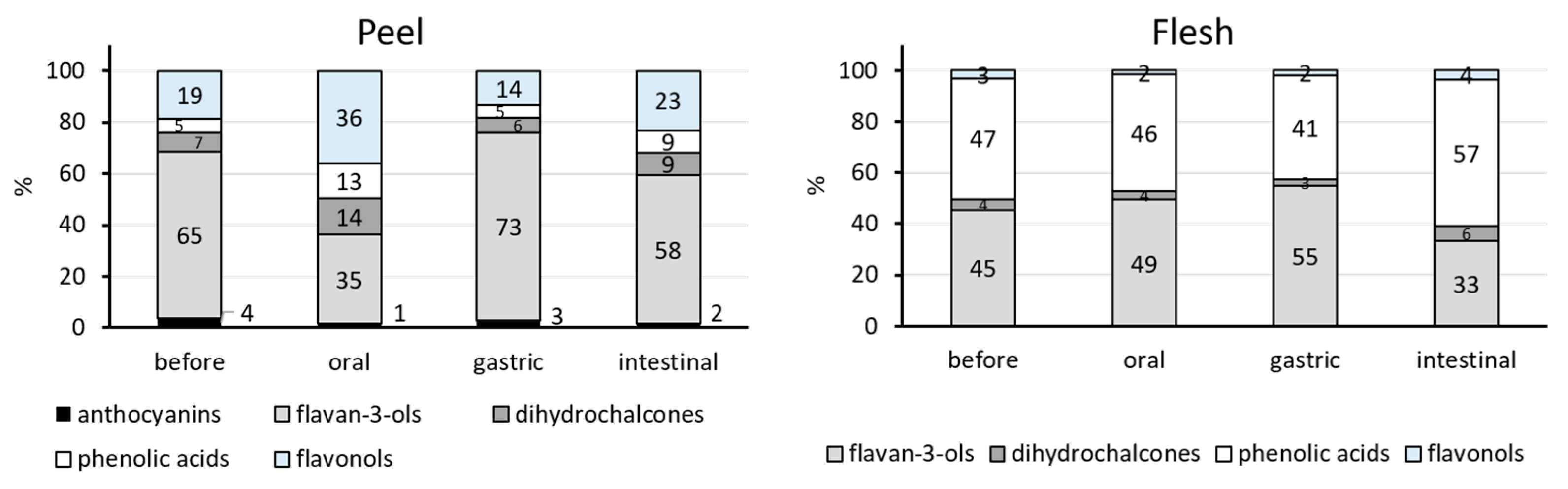
Phenolic Classes in the Simulated Digestion
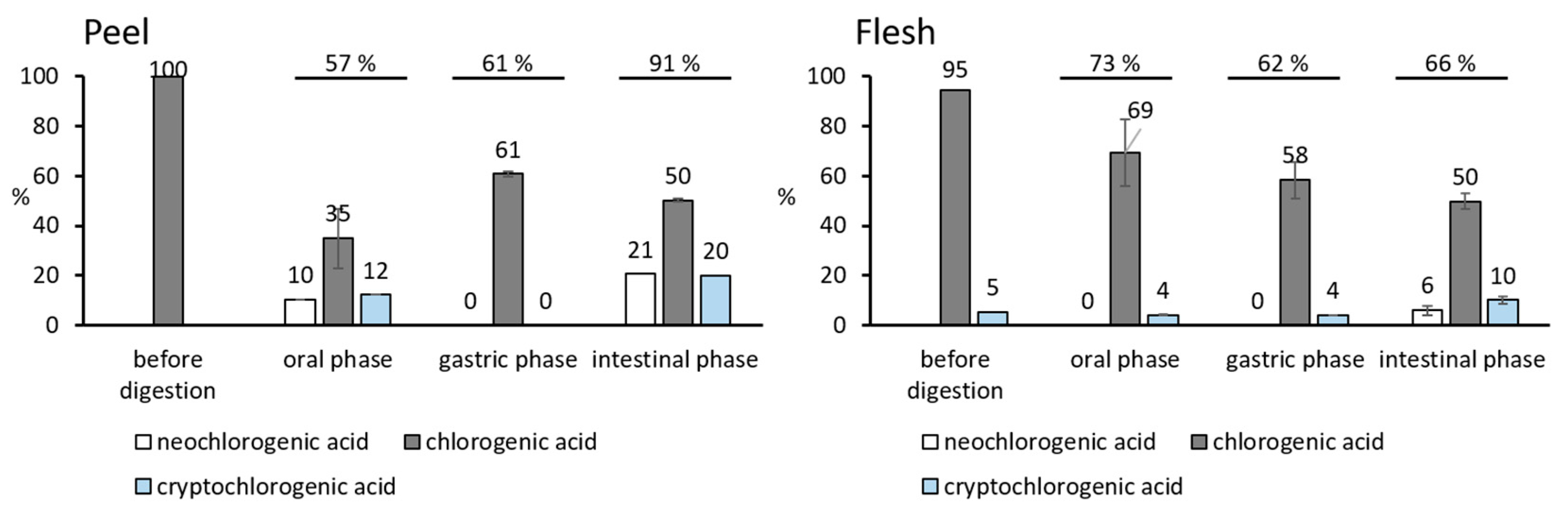
3.3. The Degradation of Chlorogenic Acids from Apples during the Digestion
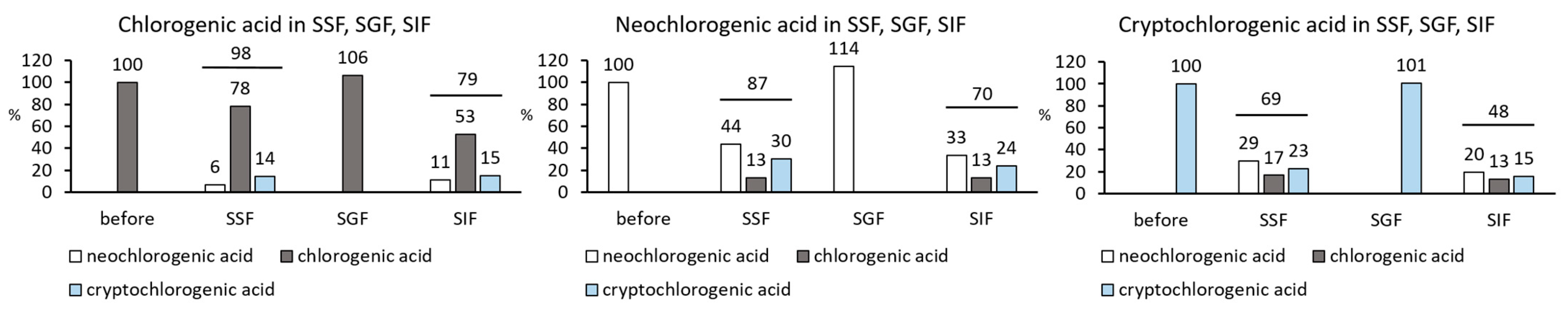
3.4. The Degradation of Chlorogenic Acids in Simulated Salivary, Gastric, and Intestinal Electrolyte Solutions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, M.A.E.; Mohanad, M.; Ahmed, A.A.E.; Aboulhoda, B.E.; El-Awdan, S.A. Mechanistic insights into the protective effects of chlorogenic acid against indomethacin-induced gastric ulcer in rats: Modulation of the cross talk between autophagy and apoptosis signaling. Life Sci. 2021, 275, 119370. [Google Scholar] [CrossRef] [PubMed]
- Hamauzu, Y.; Irie, M.; Kondo, M.; Fujita, T. Antiulcerative properties of crude polyphenols and juice of apple, and Chinese quince extracts. Food Chem. 2008, 108, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Pastene, E.; Speisky, H.; Troncoso, M.; Alarcon, J.; Figueroa, G. In vitro inhibitory effect of apple peel extract on the growth of Helicobacter pylori and respiratory burst induced on human neutrophils. J. Agric. Food Chem. 2009, 57, 7743–7749. [Google Scholar] [CrossRef] [PubMed]
- Pastene, E.; Troncoso, M.; Fiġueroa, G.; Alarcón, J.; Speisky, H. Association between polymerization degree of apple peel polyphenols and inhibition of Helicobacter pylori urease. J. Agric. Food Chem. 2009, 57, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Pastene, E.; Spiesky, H.; García, A.; Moreno, J.; Troncoso, M.; Figueroa, G. In vitro and in vivo effects of apple polyphenols against Helicobacter pylori. J. Agric. Food Chem. 2010, 58, 7172–7179. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Pozo, C.; Speisky, H.; Brunser, O.; Pastene, E.; Gotteland, M. Apple peel polyphenols protect against gastrointestinal mucosa alterations induced by indomethacin in rats. J. Agric. Food Chem. 2011, 59, 6459–6466. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ji, Y.; Guo, Y.; Wang, H.; Wu, Z.; Li, H.; Wang, H. Dietary supplementation of apple phlorizin attenuates the redox state related to gut microbiota homeostasis in C57BL/6J mice fed with a high-fat diet. J. Agric. Food Chem. 2021, 69, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Qiu, D.; Wu, Z.; Yang, H.; Xie, Y.; Li, S.; Yin, Y.; Li, X. Apple polyphenol extract alleviates DSS-induced ulcerative colitis and linked behavioral disorders via regulating the gut-brain axis. Food Biosci. 2023, 53, 102720. [Google Scholar] [CrossRef]
- Li, D.; Sun, L.; Yang, Y.; Wang, Z.; Yang, X.; Zhao, T.; Gong, T.; Zou, L.; Guo, Y. Young apple polyphenols postpone starch digestion in vitro and in vivo. J. Funct. Food 2019, 56, 127–135. [Google Scholar] [CrossRef]
- Zhao, T.; Sun, L.; Wang, Z.; Nisar, T.; Gong, T.; Li, D.; Niu, P.; Guo, Y. The antioxidant property and α-amylase inhibition activity of young apple polyphenols are related with apple varieties. LWT-Food Sci. Technol. 2019, 111, 252–259. [Google Scholar] [CrossRef]
- Gong, T.; Yang, X.; Bai, F.; Li, D.; Zhao, T.; Zhang, J.; Sun, L.; Guo, Y. Young apple polyphenols as natural α-glucosidase inhibitors: In vitro and in silico studies. Bioorg. Chem. 2020, 96, 103625. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F.; Cui, Y.; Yin, Y.; Li, S.; Li, X. Apple polyphenols extracts ameliorate high carbohydrate diet- induced body weight gain by regulating the gut microbiota and appetite. J. Agric. Food Chem. 2022, 70, 196–210. [Google Scholar] [CrossRef]
- Li, D.; Cui, Y.; Wang, X.; Liu, F.; Li, X. Apple polyphenol extract improves high-fat diet-induced hepatic steatosis by regulating bile acid synthesis and gut microbiota in C57BL/6 Male Mice. J. Agric. Food Chem. 2021, 69, 6829–6841. [Google Scholar] [CrossRef]
- Elkahoui, S.; Levin, C.E.; Bartley, G.E.; Yokoyama, W.; Friedman, M. Levels of fecal procyanidins and changes in microbiota and metabolism in mice fed a high-fat diet supplemented with apple peel. J. Agric. Food Chem. 2019, 67, 10352–10360. [Google Scholar] [CrossRef]
- Hung, C.H.; Huang, C.C.; Hsu, L.S.; Kao, S.H.; Wang, C.J. Apple polyphenol inhibits colon carcinoma metastasis via disrupting Snail binding to focal adhesion kinase. J. Funct. Food 2015, 12, 80–91. [Google Scholar] [CrossRef]
- Bars-Cortina, D.; Martínez-Bardají, A.; Macià, A.; Motilva, M.J.; Piñol-Felis, C. Consumption evaluation of one apple flesh a day in the initial phases prior to adenoma/adenocarcinoma in an azoxymethane rat colon carcinogenesis model. J. Nutr. Biochem. 2020, 83, 108418. [Google Scholar] [CrossRef]
- Khanizadeh, S.; Tsao, R.; Rekika, D.; Yang, R.; Charles, M.T.; Rupasinghe, H.P.V. Polyphenol composition and total antioxidant capacity of selected apple genotypes for processing. J. Food Compos. Anal. 2008, 21, 396–401. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Laskowski, P. Polyphenolic compounds and antioxidant activity of new and old apple varieties. J. Agric. Food Chem. 2008, 56, 6520–6530. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, R.P.; Antunes, C.; Ramos, A.; Serra, A.T.; Figueira, M.E.; Duarte, C.M.M.; de Carvalho, A.; Bronze, M.R. Characterization of traditional and exotic apple varieties from Portugal. Part 1—Nutritional, phytochemical and sensory evaluation. J. Funct. Food 2010, 2, 35–45. [Google Scholar] [CrossRef]
- Giomaro, G.; Karioti, A.; Bilia, A.R.; Bucchini, A.; Giamperi, L.; Ricci, D.; Fraternale, D. Polyphenols profile and antioxidant activity of skin and pulp of a rare apple from Marche region (Italy). Chem. Cent. J. 2014, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Lo Piccolo, E.; Landi, M.; Massai, R.; Remorini, D.; Conte, G.; Guidi, L. Ancient apple cultivars from Garfagnana (Tuscany, Italy): A potential sourcefor ‘nutrafruit’ production. Food Chem. 2019, 294, 518–525. [Google Scholar] [CrossRef]
- Stracke, B.A.; Rűfer, C.E.; Weibel, F.P.; Bub, A.; Watzl, B. Three-year comparison of the polyphenol contents and antioxidant capacities in organically and conventionally produced apples (Malus domestica Bork. Cultivar Golden Delicious). J. Agric. Food Chem. 2009, 57, 4598–4605. [Google Scholar] [CrossRef]
- Zhao, X.L.; Yu, L.; Zhang, S.D.; Ping, K.; Ni, H.Y.; Qin, X.Y.; Zhao, C.J.; Wang, W.; Efferth, T.; Fu, Y.J. Cryptochlorogenic acid attenuates LPS-induced inflammatory response and oxidative stress via upregulation of the Nrf2/HO-1 signaling pathway in RAW 264.7 macrophages. Int. Immunopharmacol. 2020, 83, 106436. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Okyere, S.K.; Hu, L.; Wen, J.; Ren, Z.; Deng, J.; Hu, Y. Anti-inflammatory activity and mechanism of cryptochlorogenic acid from Ageratina adenophora. Nutrients 2022, 14, 439. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jin, M.L.; Yi, E.H.; Kim, Y.; Park, G. Neochlorogenic acid inhibits against LPS-activated inflammatory responses through up-regulation of Nrf2/HO-1 and involving AMPK pathway. Environ. Toxicol. Pharmacol. 2018, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Deußer, H.; Hoffmann, L.; Bohn, T. Bioaccessible and dialysable polyphenols in selected apple varieties following in vitro digestion vs. their native patterns. Food Chem. 2012, 131, 1466–1472. [Google Scholar] [CrossRef]
- Graziani, G.; Gaspari, A.; Di Vaio, C.; Cirillo, A.; Ronca, C.L.; Grosso, M.; Ritieni, A. Assessment of in vitro bioaccessibility of polyphenols from Annurca, Limoncella, Red Delicious, and Golden Delicious apples using a sequential enzymatic digestion model. Antioxidants 2021, 10, 541. [Google Scholar] [CrossRef] [PubMed]
- Quatrin, A.; Rampelotto, C.; Pauletto, R.; Maurer, L.H.; Nichelle, S.M.; Klein, B.; Fritzsche Rodrigues, R.; Maróstica Junior, M.R.; de Souza Fonseca, B.; Ragagnin de Menezes, C.; et al. Bioaccessibility and catabolism of phenolic compounds from jaboticaba (Myrciaria trunciflora) fruit peel during in vitro gastrointestinal digestion and colonic fermentation. J. Funct. Food 2020, 65, 103714. [Google Scholar] [CrossRef]
- Fernández-Jalao, I.; Balderas, C.; Sánchez-Moreno, C.; De Ancos, B. Impact of an in vitro dynamic gastrointestinal digestion on phenolic compounds and antioxidant capacity of apple treated by high-pressure processing. Innov. Food Sci. Emerg. 2020, 66, 102486. [Google Scholar] [CrossRef]
- Jakobek, L.; Ištuk, J.; Matić, P.; Skendrović Babojelić, M. Interactions of polyphenols from traditional apple varieties ‘Bobovac’, ‘Ljepocvjetka’ and ‘Crvenka’ with β-Glucan during in vitro simulated digestion. Food Chem. 2021, 363, 130283. [Google Scholar] [CrossRef]
- Jakobek, L.; Ištuk, J.; Barron, A.; Matić, P. Bioactive phenolic compounds from apples during simulated in vitro gastrointestinal digestion: Kinetics of their release. Appl. Sci. 2023, 13, 8434. [Google Scholar] [CrossRef]
- Zahid, H.F.; Ali, A.; Ranadheera, C.S.; Fang, Z.; Ajlouni, S. Identification of phenolics profile in freeze-dried apple peel and their bioactivities during in vitro digestion and colonic fermentation. Int. J. Mol. Sci. 2023, 24, 1514. [Google Scholar] [CrossRef]
- Tenore, G.C.; Campiglia, P.; Ritieni, A.; Novellino, E. In vitro bioaccessibility, bioavailability and plasma protein interaction of polyphenols from Annurca apple (M. pumila Miller cv Annurca). Food Chem. 2013, 141, 3519–3524. [Google Scholar] [CrossRef]
- Kahle, K.; Huemmer, W.; Kempf, M.; Scheppach, W.; Erk, T.; Richling, E. Polyphenols are intensively metabolized in the human gastrointestinal tract after apple juice consumption. J. Agric. Food Chem. 2007, 55, 10605–10614. [Google Scholar] [CrossRef]
- Xi, Q.; Hoth-Hannig, W.; Deng, S.; Jin, X.; Fu, B.; Hannig, M. The effect of polyphenol-containing solutions on in situ biofilm formation on enamel and dentin. J. Dent. 2020, 102, 103482. [Google Scholar] [CrossRef] [PubMed]
- Hannig, C.; Sorg, J.; Spitzmüller, B.; Hannig, M.; Al-Ahmad, A. Polyphenolic beverages reduce initial bacterial adherence to enamel in situ. J. Dent. 2009, 37, 560–566. [Google Scholar] [CrossRef]
- Gorelik, S.; Ligumsky, M.; Kohen, R.; Kanner, J. The stomach as a “bioreactor”: When red meat meets red wine. J. Agric. Food Chem. 2008, 56, 5002–5007. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, S.; Ligumsky, M.; Kohen, R.; Kanner, J. A novel function of red wine polyphenols in humans: Prevention of absorption of cytotoxic lipid peroxidation products. Fed. Am. Soc. Exp. Biol. 2008, 22, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Kuffa, M.; Priesbe, T.J.; Krueger, C.G.; Reed, J.D.; Richards, M.P. Ability of dietary antioxidants to affect lipid oxidation of cooked turkey meat in a simulated stomach and blood lipids after a meal. J. Funct. Food 2009, 1, 208–216. [Google Scholar] [CrossRef]
- da Silva, L.M.; Pezzini, B.C.; Bordignon Somensi, L.; Bolda Mariano, L.N.; Mariott, M.; Boeing, T.; dos Santos, A.C.; Longo, B.; Cechinel-Filho, V.; de Souza, P.; et al. Hesperidin, a citrus flavanone glycoside, accelerates the gastric healing process of acetic acid-induced ulcer in rats. Chem-Biol. Interact. 2019, 308, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Valcheva-Kuzmanova, S.; Denev, P.; Eftimov, M.; Georgieva, A.; Kuzmanova, V.; Kuzmanov, A.; Kuzmanov, K.; Tzaneva, M. Protective effects of Aronia melanocarpa juices either alone or combined with extracts from Rosa canina or Alchemilla vulgaris in a rat model of indomethacin-induced gastric ulcers. Food Chem. Toxicol. 2019, 132, 110739. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Before Digestion | Oral Phase | Gastric Phase | Intestinal Phase | |
|---|---|---|---|---|
| Anthocyanins | ||||
| cyanidin-3-galactoside | 83.5 ± 2.6 a | 7.4 ± 1.0 d | 40.7 ± 3.0 b | 20.8 ± 4.6 c |
| total | 83.5 ± 2.6 a | 7.4 ± 1.0 d | 40.7 ± 3.0 b | 20.8 ± 4.6 c |
| Flavan-3-ols | ||||
| procyanidin B1 | 134.2 ± 5.5 a | 18.9 ± 10.1 c | 142.9 ± 4.4 a | 70.8 ± 12.2 b |
| (+)-catechin | 163.6 ± 14.0 a | 20.2 ± 7.0 b | 56.8 ± 8.5 b | 46.1 ± 19.4 b |
| procyanidin B2 | 456.6 ± 12.0 a | 46.0 ± 27.1 c | 484.3 ± 23.1 a | 323.8 ± 68.3 b |
| (−)-epicatechin | 646.2 ± 15.3 a | 88.9 ± 22.6 d | 353.7 ± 16.7 b | 257.5 ± 26.8 c |
| total | 1400.6 ± 35.1 a | 174.0 ± 66.7 d | 1037.7 ± 52.8 b | 698.2 ± 126.8 c |
| Dihydrochalcones | ||||
| phloretin-2′-glucoside | 160.1 ± 8.0 a | 71.5 ± 10.0 c | 80.1 ± 5.9 c | 106.1 ± 1.4 b |
| total | 160.1 ± 8.0 a | 71.5 ± 10.0 c | 80.1 ± 5.9 c | 106.1 ± 1.4 b |
| Phenolic acids | ||||
| neochlorogenic acid | nd | 11.8 ± 4.1 b | nd | 23.9 ± 0.3 a |
| chlorogenic acid | 115.7 ± 4.1 a | 40.4 ± 13.8 c | 70.4 ± 1.3 b | 58.2 ± 0.5 b,c |
| cryptochlorogenic acid | nd | 14.2 ± 4.0 b | nd | 23.0 ± 0.3 a |
| total | 115.7 ± 4.1 a | 66.4 ± 21.9 b | 70.4 ± 1.3 b | 105.1 ± 0.1 a |
| Flavonols | ||||
| quercetin-3-galactoside | 70.4 ± 3.9 a | 32.9 ± 9.7 b | 36.4 ± 3.8 b | 46.8 ± 1.0 b |
| quercetin-3-glucoside | 42.6 ± 1.3 a | 19.1 ± 2.8 c | 19.3 ± 1.7 c | 33.7 ± 1.7 b |
| quercetin derivative * | 55.5 ± 1.1 a | 20.3 ± 3.1 c | 21.7 ± 1.9 c | 34.8 ± 0.4 b |
| quercetin-3-xyloside * | 95.5 ± 1.7 a | 39.5 ± 6.6 c | 41.2 ± 3.6 c | 63.2 ± 0.9 b |
| quercetin-3-rhamnoside | 138.9 ± 1.8 a | 69.8 ± 11.2 c | 73.9 ± 7.5 c | 102.5 ± 5.9 b |
| total | 402.9 ± 9.8 a | 181.6 ± 33.5 c | 192.5 ± 18.6 c | 281.0 ± 5.7 b |
| TOTAL | 2162.8 ± 50.3 a | 500.9 ± 133.1 c | 1421.4 ± 81.5 b | 1211.2 ± 124.4 b |
| Before Digestion | Oral Phase | Gastric Phase | Intestinal Phase | |
|---|---|---|---|---|
| Flavan-3-ols | ||||
| procyanidin B1 | 38.9 ± 6.4 b | 57.3 ± 0.4 a | 65.4 ± 1.6 a | Nd |
| (+)-catechin | 43.6 ± 5.2 a | 19.6 ± 2.1 b | 18.3 ± 0.0 b | 17.2 ± 0.0 b |
| procyanidin B2 | 121.7 ± 19.5 a | 112.1 ± 6.1 a | 126.1 ± 1.6 a | 53.1 ± 18.0 b |
| (−)-epicatechin | 88.9 ± 14.1 a | 55.1 ± 3.9 b | 56.7 ± 0.3 b | 44.6 ± 3.4 b |
| total | 293.1 ± 45.2 a | 244.1 ± 11.7 a | 266.5 ± 3.5 a | 114.9 ± 21.4 b |
| Dihydrochalcones | ||||
| phloretin-2′-glucoside | 26.8 ± 5.3 a | 17.7 ± 11.4 a | 12.3 ± 1.6 a | 20.3 ± 7.8 a |
| total | 26.8 ± 5.3 a | 17.7 ± 11.4 a | 12.3 ± 1.6 a | 20.3 ± 7.8 a |
| Phenolic acids | ||||
| neochlorogenic acid | nd | Nd | nd | 16.0 ± 4.9 a |
| chlorogenic acid | 250.4 ± 37.8 a | 183.7 ± 35.1 a,b | 154.5 ± 19.5 b | 132.1 ± 7.8 b |
| cryprochlorogenic acid | 14.5 ± 1.8 b | 10.9 ± 0.5 b | 10.5 ± 0.2 b | 26.9 ± 4.0 a |
| unknown phenolic acid * | 29.4 ± 0.9 a | 23.1 ± 0.6 b | 24.8 ± 1.9 b | 18.5 ± 0.7 c |
| p-coumaroylquinic acid * | 11.0 ± 1.9 a | 7.4 ± 0.6 a,b | 7.1 ± 0.4 a,b | 4.7 ± 0.1 b |
| total | 305.3 ± 40.7 a | 225.1 ± 36.8 a,b | 196.9 ± 22.1 b | 198.2 ± 16.0 b |
| Flavonols | ||||
| quercetin-3-galactoside | 0.1 ± 0.0 b | 0.2 ± 0.0 a | 0.2 ± 0.0 a,b | 0.1 ± 0.0 b |
| quercetin-3-glucoside | 7.6 ± 0.3 a | 3.6 ± 0.2 c | 4.5 ± 0.3 c | 6.2 ± 0.5 b |
| quercetin derivative | 4.6 ± 0.1 a | Nd | nd | Nd |
| quercetin-3-xyloside | 4.8 ± 0.2 a | 2.4 ± 0.1 c | 3.0 ± 0.2 c | 4.2 ± 0.3 b |
| quercetin-3-rhamnoside | 2.1 ± 0.5 a | 1.5 ± 0.4 a | 1.4 ± 0.0 a | 1.7 ± 0.3 a |
| total | 19.2 ± 1.2 a | 7.7± 0.0 c | 9.1 ± 0.5 b,c | 12.2 ± 1.1 b |
| TOTAL | 644.4 ± 92.3 a | 494.6 ± 59.9 a,b | 484.8 ± 27.7 a,b | 345.6 ± 3.6 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakobek, L.; Pöc, K.; Valenteković, M.; Matić, P. The Behavior of Phenolic Compounds from Apples during Simulated Gastrointestinal Digestion with Focus on Chlorogenic Acid. Foods 2024, 13, 693. https://doi.org/10.3390/foods13050693
Jakobek L, Pöc K, Valenteković M, Matić P. The Behavior of Phenolic Compounds from Apples during Simulated Gastrointestinal Digestion with Focus on Chlorogenic Acid. Foods. 2024; 13(5):693. https://doi.org/10.3390/foods13050693
Chicago/Turabian StyleJakobek, Lidija, Kristina Pöc, Matea Valenteković, and Petra Matić. 2024. "The Behavior of Phenolic Compounds from Apples during Simulated Gastrointestinal Digestion with Focus on Chlorogenic Acid" Foods 13, no. 5: 693. https://doi.org/10.3390/foods13050693
APA StyleJakobek, L., Pöc, K., Valenteković, M., & Matić, P. (2024). The Behavior of Phenolic Compounds from Apples during Simulated Gastrointestinal Digestion with Focus on Chlorogenic Acid. Foods, 13(5), 693. https://doi.org/10.3390/foods13050693