3.2. The Effect of Milk Protein Concentration on the Elastic Modulus and Processing Time
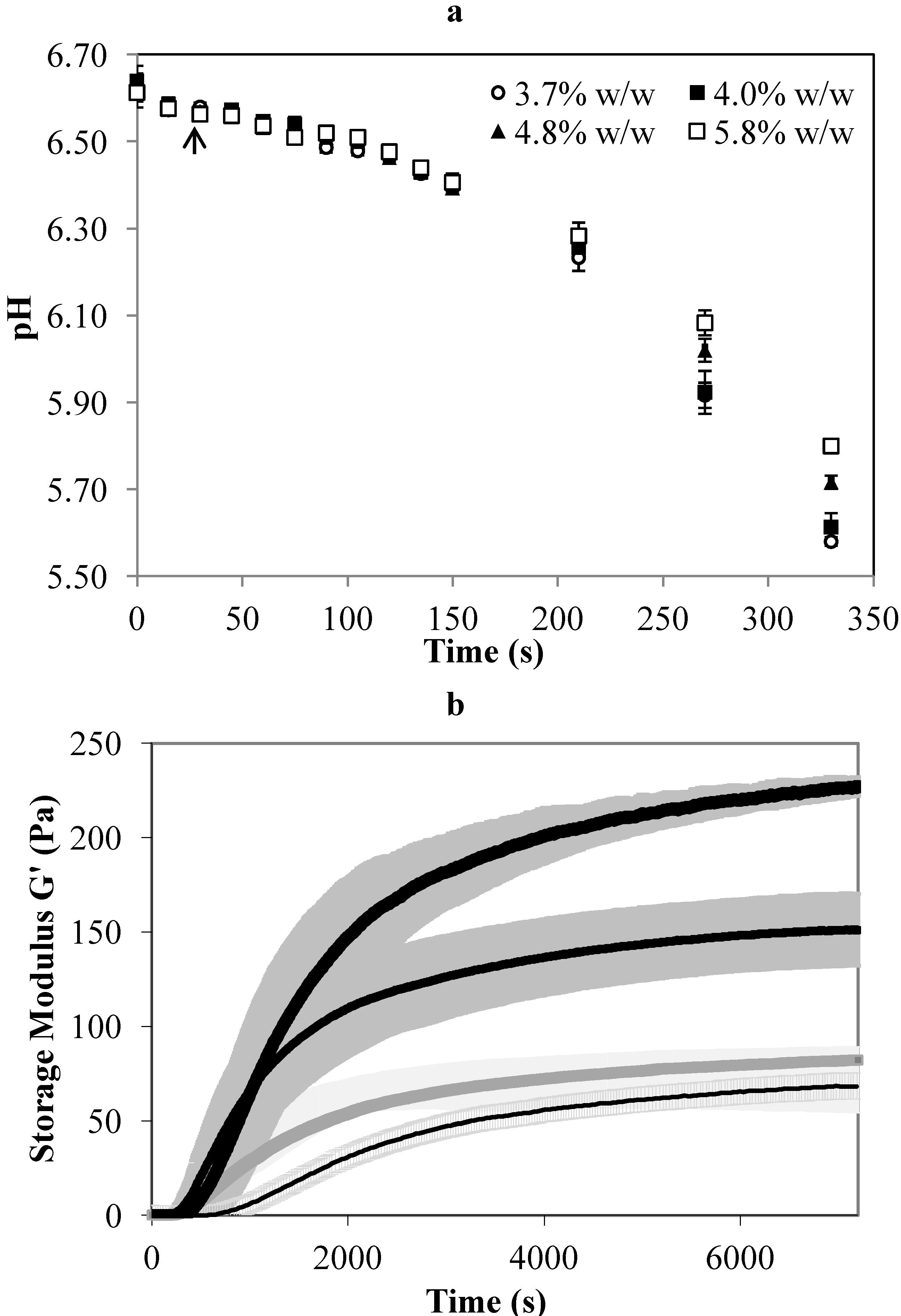
The storage modulus (G’) of the gel formed from milk with a higher protein concentration was significantly higher than for milk with a lower protein concentration (
Figure 1b). The rate of gel firming also increased rapidly in milk with a higher protein concentration, as reflected by the gradient of the slope of the storage modulus and the decrease in the time required to obtain a given value of G’. The G’ increased rapidly between 200 and 400 s for all gels (
Figure 1b), indicating a similar onset of gelation.
In our previous experiments, the time from rennet addition to cutting, known as the setting time of the gel, was set at 2700 s corresponding to a G’ of 60 Pa, as this gave a Cheddar cheese with acceptable moisture content using standardised milk with a protein concentration of 4% w/w [
15]. To maintain a similar level of stiffness of 60 Pa, the setting time of the gel made with 4.8% or 5.8% w/w milk protein was set to 1200 s (
Figure 1b). This adjustment prevents the curd becoming too firm and avoids curd fines in the whey. Another way to reduce the firmness of a gel is to reduce the coagulation temperature or reduce the amount of rennet used [
1,
19]. The concentration of rennet added per unit mass of milk was similar here (
Table 1), but the rennet concentration per kg of milk protein was consequently lower in the milk with a higher protein concentration (
Table 1). Further reducing the rennet for high protein treatments would impact on the residual rennet in the final cheese. For this reason, the same concentration of rennet per unit mass of milk was used in this study. The time required for the unconcentrated milk to reach G’ = 60 Pa was about 5000 s (
Figure 1b). During the actual cheese-making experiment, however, the milk in the cheese vat reached a gel consistency of medium firmness and was ready for cutting at 2700 s. The longer time indicated by the rheology analysis could possibly be due to a small degree of shearing applied to the sample during analysis that disturbed the gel formation. This was not apparent in the concentrated sample, but the setting time indicated by the rheology analysis in this study was less accurate for gel made with the unconcentrated milk.
The processing time from the addition of starter culture to whey draining or milling in the pilot scale cheese-making was not significantly different (p > 0.05) (data not shown), suggesting that adjustments made to the starter concentration and the setting time described above were effective.
3.3. Composition of Cheese-Milk, Sweet Whey and Salty Whey
Ultrafiltration is known to alter the composition of milk. Approximately 81% of the protein within the LCUFR used in this study was casein as compared to 77% in unconcentrated milk. The LCUFR also contains less non protein nitrogen (NPN) arising from low molecular weight nitrogen containing compounds, such as creatine and creatinine and urea [
20] as a portion of the NPN permeates through the UF membrane during ultrafiltration [
21]. Consequently the concentration of NPN as a percentage of total nitrogen (TN) differed significantly in the LCUFR and unconcentrated milk from 2.5% w/w to 4.6% w/w, respectively.
The impact of standardization of cheese-milk on casein and NPN content is shown in
Table 2. As the casein concentration increased with LCUFR addition, the ratio of casein nitrogen over TN increased and the ratio of NPN over TN decreased. These different CN/TN or NPN/TN concentrations in the cheese-milk are expected to impact the final cheese yield [
22].
Table 2.
Protein composition, total solids and total calcium content of cheese-milk used for pilot scale cheese-making #.
Table 2.
Protein composition, total solids and total calcium content of cheese-milk used for pilot scale cheese-making #.
| Batch | Casein (% w/w) | CN/TN (% w/w) | NPN/TN (% w/w) | Total solids (% w/w) | Total calcium (mg/kg) | P/F ratio |
|---|
| 1 | 2.83 ± 0.04 d | 76.7 ± 0.00 d | 4.61 ± 0.00 a | 13.6 ± 0.14 d | 985 ± 62 d | 0.84 ± 0.00 |
| 2 | 3.12 ± 0.01 c | 77.5 ± 0.15 c | 4.19 ± 0.09 b | 14.4 ± 0.07 c | 1108 ± 31 c | 0.83 ± 0.01 |
| 3 | 3.78 ± 0.28 b | 78.9 ± 0.19 b | 3.42 ± 0.10 c | 15.9 ± 1.20 b | 1223 ± 52 b | 0.85 ± 0.04 |
| 4 | 4.60 ± 0.04 a | 79.9 ± 0.00 a | 2.83 ± 0.02 d | 18.1 ± 0.53 a | 1500 ± 34 a | 0.83 ± 0.00 |
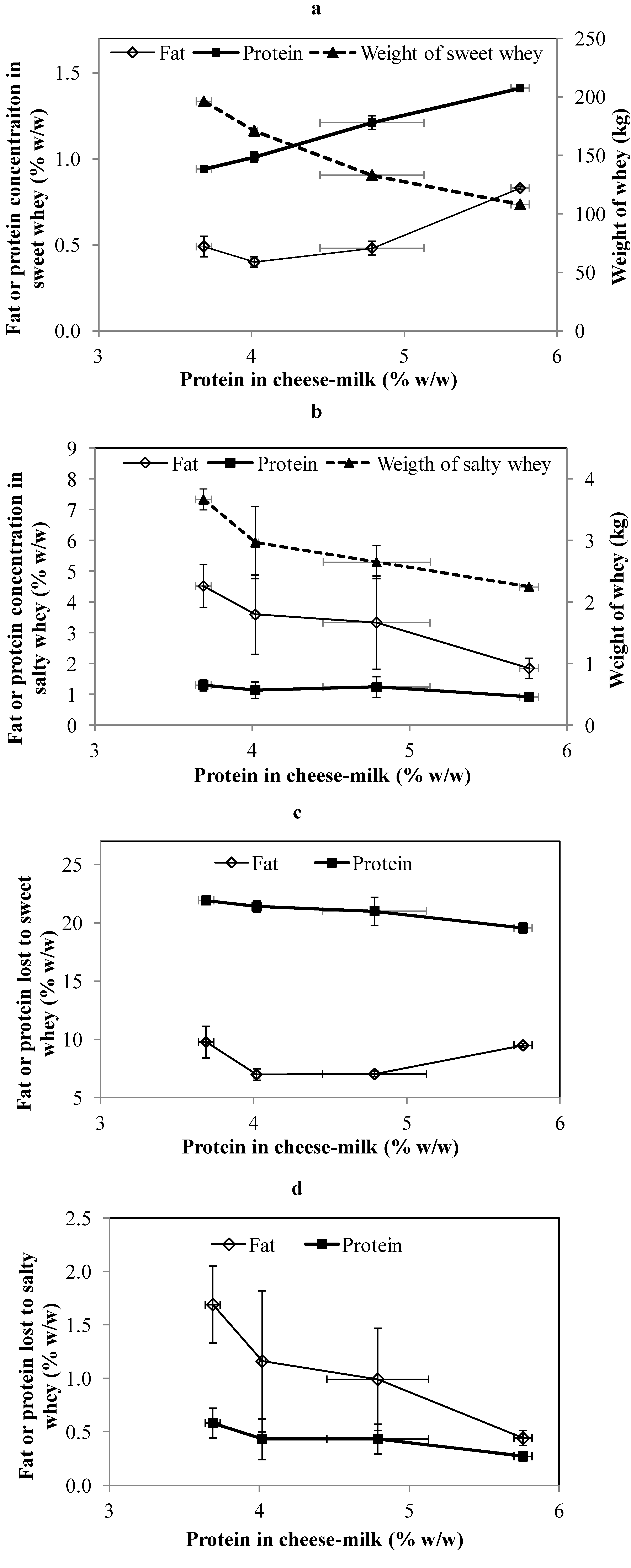
The weight and composition of the whey generated during cheese-making at pilot scale is shown in
Figure 2a,b. The weight of sweet whey and salty whey was significantly reduced (
p < 0.05) for samples made with higher protein milk. This was expected and is consistent with previous reports that less processing of whey is required when milk with a higher protein concentration is used for cheese-making [
2]. The percentage of fat lost to the sweet whey was significantly higher (
p < 0.05) in cheese made from milk without the addition of LCUFR (
Figure 2c). Greater fat loss also occurred when the concentration of milk protein was the highest (5.8% w/w) despite the increase in CN/TN, with minimum levels of fat loss occurring for samples made using cheese-milk with 4% or 4.8% (w/w) protein (
Figure 2c). Conversely, while the percentage of protein in the sweet whey increased continuously as the protein in the cheese milk increased (
Figure 2a), the total protein lost to this whey stream reduced continuously as the protein concentration in the cheese milk increased (
Figure 2c).
Figure 2.
The composition and weight of sweet whey (a) and salty whey (b) collected during cheese-making at pilot scale using cheese-milk with different protein concentrations. The percentage of fat lost or protein lost to the sweet whey (c) and salty whey (d) were calculated on the basis of total fat or protein in the cheese-milk. Results are expressed as mean ± standard deviation of mean (n = 3).
Figure 2.
The composition and weight of sweet whey (a) and salty whey (b) collected during cheese-making at pilot scale using cheese-milk with different protein concentrations. The percentage of fat lost or protein lost to the sweet whey (c) and salty whey (d) were calculated on the basis of total fat or protein in the cheese-milk. Results are expressed as mean ± standard deviation of mean (n = 3).
The variability in fat composition measured for the salty whey was quite high. This could be due to the sampling technique used to obtain the salty whey at a pilot scale, as it was noted that some fat adhered to the cheese-press instead of flowing out with the salty whey, leading to greater variability. There is still clear evidence, however, of both a decline in the concentration of fat in the salty whey and the total fat loss (
Figure 2b,d). The trends in the protein data are less clear but there is still some evidence of a decrease in protein loss into the salty whey as the protein content of the original cheese milk increases (
Figure 2d).
3.4. Fat and Protein Retention, the Composition and Yield of Cheese
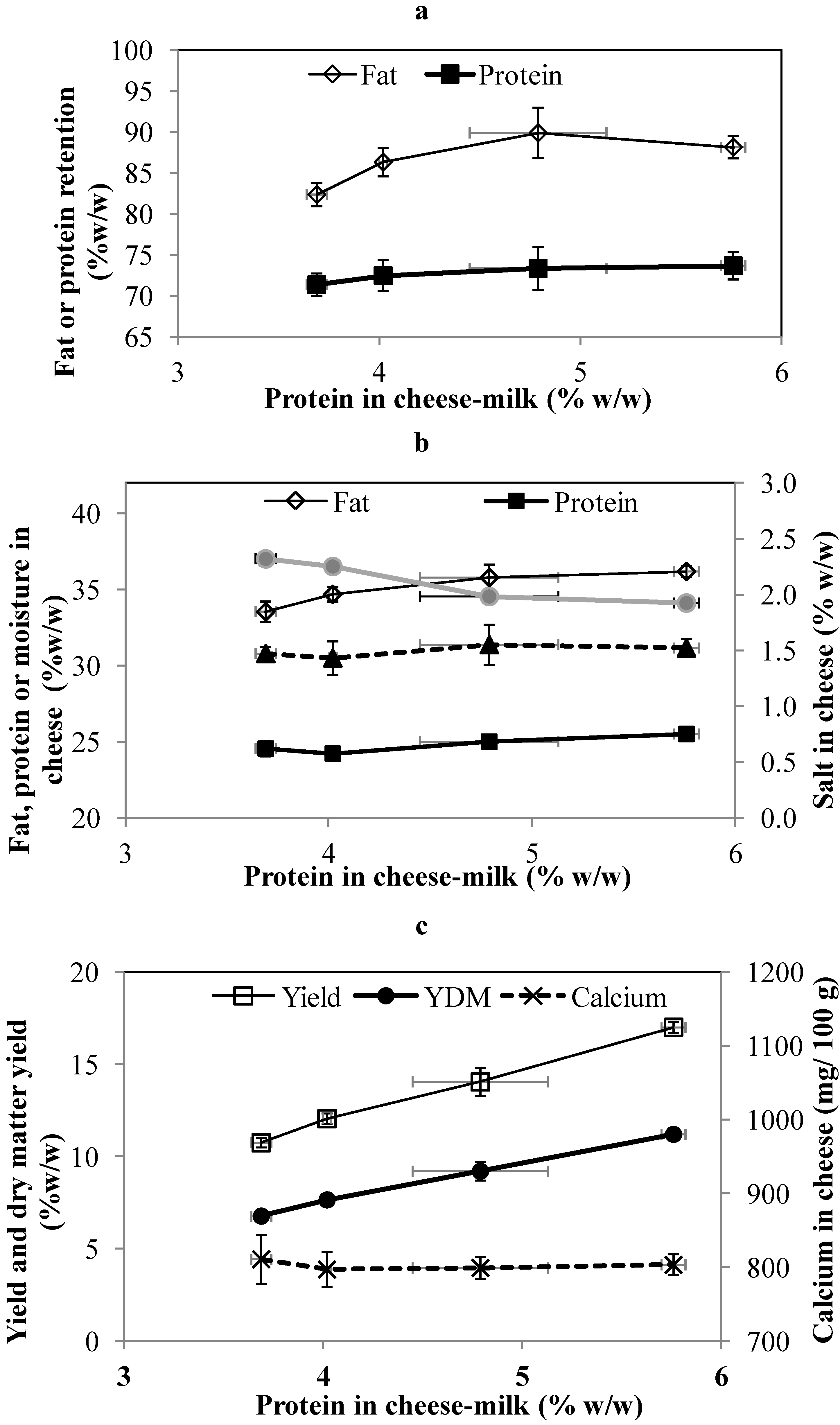
Consistent with the changes in the sweet and salty whey, the percentage of protein retained in the cheese increased with the protein in the cheese milk, although the trend is within experimental error (
Figure 3a). The percentage of fat retained in the cheese also increases up to a level of 4.8% w/w total protein in the cheese milk. No further increase is observed at 5.8 w/w protein. In a similar study, Govindasamy-Lucey
et al. [
3] reported no significant difference in fat retention when increasing the milk protein concentration from 3.2% to 5.0% w/w using whole milk UF retentate [
3]. Overall, the mean values of fat retention for all cheeses (
Figure 3a) were within the range of values (~83%–92% w/w) reported elsewhere for full fat Cheddar cheese [
23].
The composition of all cheeses was also within the acceptable range for Cheddar cheese. The fat content of the cheese made with LCUFR standardized cheese-milk with 4.8% or 5.8% w/w protein was significantly higher (
p < 0.05) than cheese made using unconcentrated cheese-milk (
Figure 3b). This could be due to the higher fat retention but also the lower moisture content of these cheeses (
Figure 3b). The effect on cheese moisture suggests that the addition of LCUFR could be used as a tool to reduce the moisture content of Cheddar cheese when seasonal differences make it difficult to achieve a low moisture Cheddar [
24]. The protein and salt content were not significantly different between different treatments (
p > 0.05,
Figure 3b). Our results are in agreement with a previous study [
1], where milk protein level was increased in the range of 30–70 g/L with little effect on Cheddar composition.
The weight of the cheeses made using cheese-milk with ~3.7%, 4%, 4.8% and 5.8% w/w protein were 24.4 ± 1.7, 24.1 ± 0.56, 22.5 ± 1.3 and 23.0 ± 0.41 kg respectively. The yield calculation on the basis of milk volume and the dry matter yield increased linearly with protein content (
Figure 3c), due to the higher protein and total solids content of the LCUFR standardized cheese-milk (
Table 2).
The concentration of total calcium in the cheese-milk increased with the increased concentration of protein (
Table 2), although the ratio of total calcium (TC) over total protein (TP) in the cheese-milk was similar at 24 mg/g to 28 mg/g (
p > 0.05). The concentration of TC in the whey was not significantly different (
p > 0.05). The TC in the whey was 380 ± 32, 392 ± 40, 383 ± 26 and 416 ± 29 mg/kg from cheese-making with 3.7%, 4%, 4.8% and 5.8 % w/w milk protein, respectively. Increasing the concentration of milk protein did not affect the total calcium in the final cheese (
Figure 3c). Less milk was added to the cheese vat for the treatments with increasing concentrations of protein. This adjustment led to similar amount of TC at a range between 230 g to 200 g in all vats regardless of protein concentration.
Figure 3.
The fat and protein retention (a), composition (b) and yield (c) of cheese prepared at pilot scale using cheese-milk with different protein concentrations. Fat retention or protein retention in the cheese were calculated on the basis of total fat or protein in the cheese-milk. Results are expressed as mean ± standard deviation of mean (n = 3).
Figure 3.
The fat and protein retention (a), composition (b) and yield (c) of cheese prepared at pilot scale using cheese-milk with different protein concentrations. Fat retention or protein retention in the cheese were calculated on the basis of total fat or protein in the cheese-milk. Results are expressed as mean ± standard deviation of mean (n = 3).
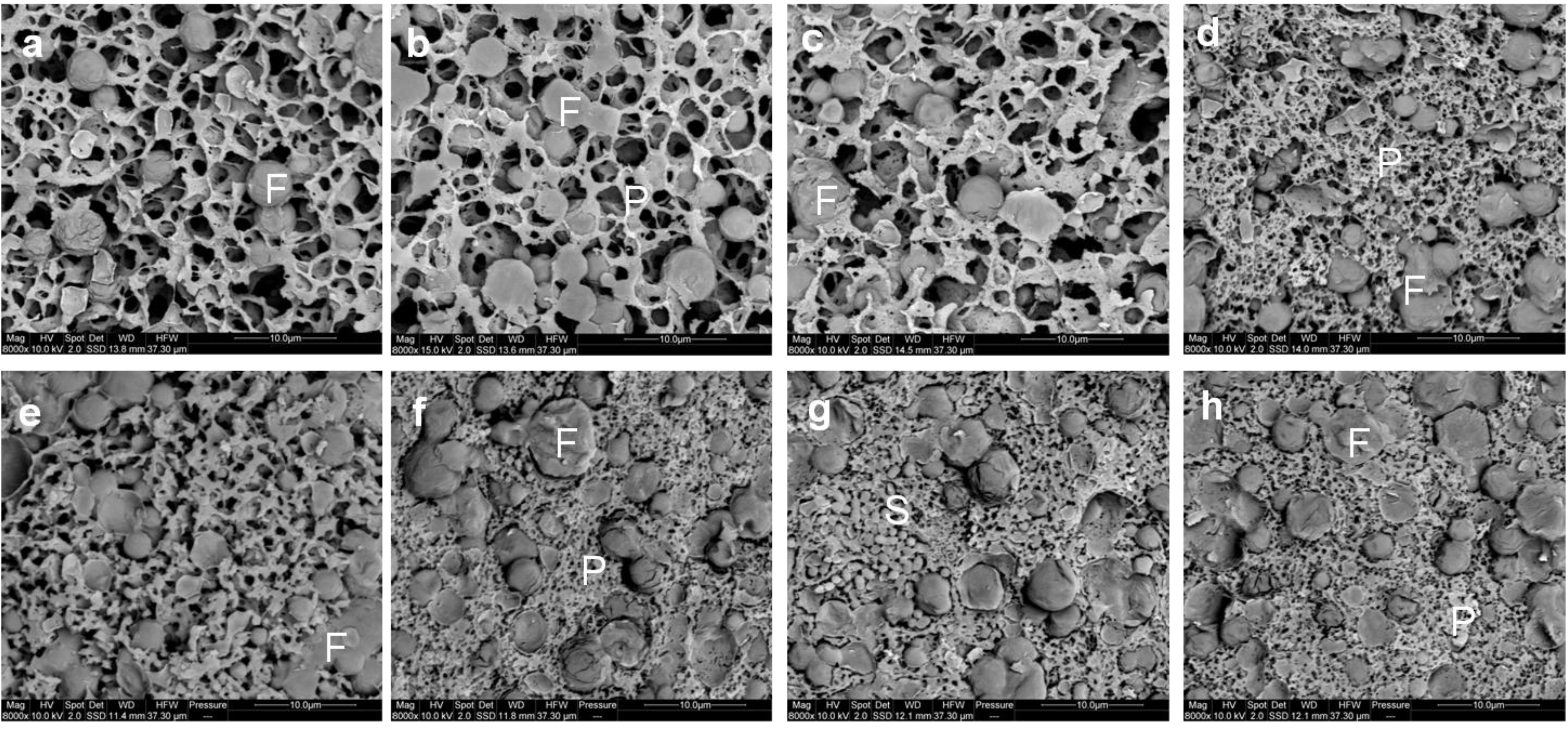
3.5. Microstructure of Gel, Curd and Cheese
The microstructure of the gel observed using cryo SEM is shown in
Figure 4a–d. After the addition of rennet, casein micelles aggregate to form a protein network in which the fat globules are entrapped. Qualitatively, there was no clear distinction between the microstructure of the gels made using cheese-milk with ~3.7%–4.8% w/w milk protein (
Figure 4a–c respectively). The microstructure of the gel with 5.8% w/w milk protein, however, was denser with smaller pores (indicated by the black areas in
Figure 4d). This dense network might arise due to the increased aggregation of casein micelles. As discussed previously, the high concentration of protein in the samples decreased the mean free distance between casein micelles, resulting in a densely aggregated CM network.
Figure 4.
Cryo SEM micrographs of gels (a–d) and cooked curd (e–h) made using (a,e) unconcentrated cheese-milk with ~3.7% w/w protein or cheese-milk standardized with LCUFR to (b,f) 4% w/w, (c,g) 4.8% w/w or (d,h) 5.8% w/w protein, where F = fat, P = protein network and S= starter culture bacteria. The gel samples were fixed in slushed liquid nitrogen when it reached the cutting time at 60 Pa. Scale bars within the images are 10 μm in length.
Figure 4.
Cryo SEM micrographs of gels (a–d) and cooked curd (e–h) made using (a,e) unconcentrated cheese-milk with ~3.7% w/w protein or cheese-milk standardized with LCUFR to (b,f) 4% w/w, (c,g) 4.8% w/w or (d,h) 5.8% w/w protein, where F = fat, P = protein network and S= starter culture bacteria. The gel samples were fixed in slushed liquid nitrogen when it reached the cutting time at 60 Pa. Scale bars within the images are 10 μm in length.
It is interesting to note that the gel stiffness was similar for all samples regardless of the protein concentration during cutting but the structure of the gel made from milk with 5.8% protein was very different. The stiffness of the gel could be attributed to two factors; the number of aggregating CM and the strength of the bonds between the CM and the relative importance of these factors may differ between treatments.
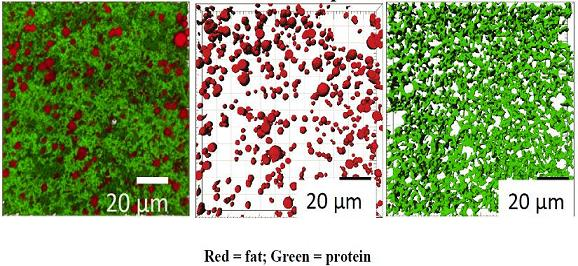
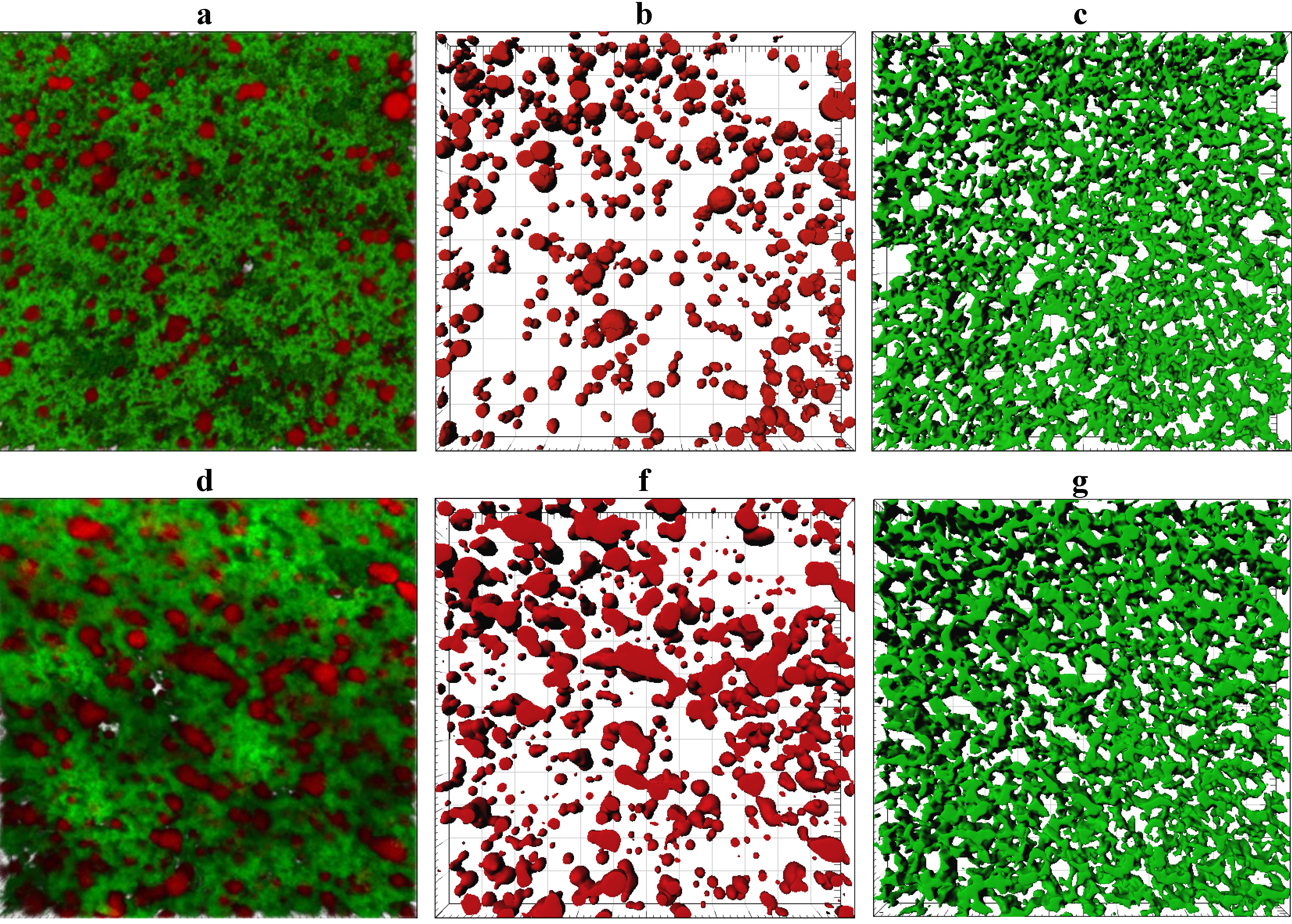
Representative CLSM images showing the microstructure of gel made from ~3.7% and 5.8% w/w milk protein are shown in
Figure 5 and a complete set of 2D CLSM images is given in
supplementary Figure S1. Despite a difference of ~2% in fat content between cheese-milk with ~3.7% w/w and 5.8% w/w protein samples, the number of fat globules and the total volume of the fat globules were not significantly different within the gel (
p > 0.05,
supplementary Figure S2a,e). The image analysis output shows a variation greater than 2% which made it hard to detect the small variation in fat content. The sphericity, fat droplet volume and fat droplet diameter were also similar (
p > 0.05,
supplementary Figure S2b–d). A lower porosity was observed for the gel with 5.8% w/w milk protein (
supplementary Figure S2f), consistent with the denser structure observed by cryo SEM.
Figure 5.
Confocal laser scanning microscopy (CLSM) microstructure of gels made using unconcentrated cheese-milk with ~3.7% w/w milk protein (a–c) or cheese-milk standardized with LCUFR to 5.8% w/w milk protein (d–f). All images are 3D reconstructions consisting of 40 layers where the separation between layers is 0.25 µm giving a total observation depth of 10 µm. Nile red stained fat appears red and fast green stained protein appears green. Images b–f are the rendered volume of the CLSM images, where b and e show the rendered volume of the fat; c and f show the rendered volume of the protein. All scale bars are 20 µm in length.
Figure 5.
Confocal laser scanning microscopy (CLSM) microstructure of gels made using unconcentrated cheese-milk with ~3.7% w/w milk protein (a–c) or cheese-milk standardized with LCUFR to 5.8% w/w milk protein (d–f). All images are 3D reconstructions consisting of 40 layers where the separation between layers is 0.25 µm giving a total observation depth of 10 µm. Nile red stained fat appears red and fast green stained protein appears green. Images b–f are the rendered volume of the CLSM images, where b and e show the rendered volume of the fat; c and f show the rendered volume of the protein. All scale bars are 20 µm in length.
CLSM provides a poorer resolution of the protein network compared to cryo SEM but a better overview of the distribution of fat and an even distribution of fat was seen within the gel made using the unconcentrated milk (
Figure 5b). The porous nature of the protein network in this sample (
Figure 5c), however, suggests some of this fat may be lost to the whey after cutting. In contrast, the fat globules tended to pool together at higher protein concentrations and the fat distribution was more heterogeneous (
Figure 5e).
Due to the nature of the pilot study occurring at a manufacturing site, samples of cooked curd, milled curd and cheese were stored at 4 °C for 2 days prior to observation by CLSM. A separate laboratory study was conducted to assess the impact of sample storage and ensure that any small changes in structure did not cause differences between treatments. A small degree of compaction was observed for all these samples regardless of protein content (data not shown) and a small degree of syneresis observed for the cooked curd. The trend of samples made from 5.8% w/w protein displaying a denser structure was not found to be obscured by storage.
The microstructure of the cooked curd samples produced at a pilot scale as observed by cryo SEM, is shown in
Figure 4e–h. The heat treatment applied during cooking at 38 °C resulted in further fusion of CM particles within each sample compared to the gel (
Figure 4a–d). Some bacterial cells could also be observed within the cooked curd sample (
Figure 4g). The only difference in the observed microstructure was the bigger pores remaining within the cooked curd prepared using the unconcentrated cheese-milk (
Figure 4e), as compared to cooked curd made using milk with a higher protein concentration (
Figure 4f–h).
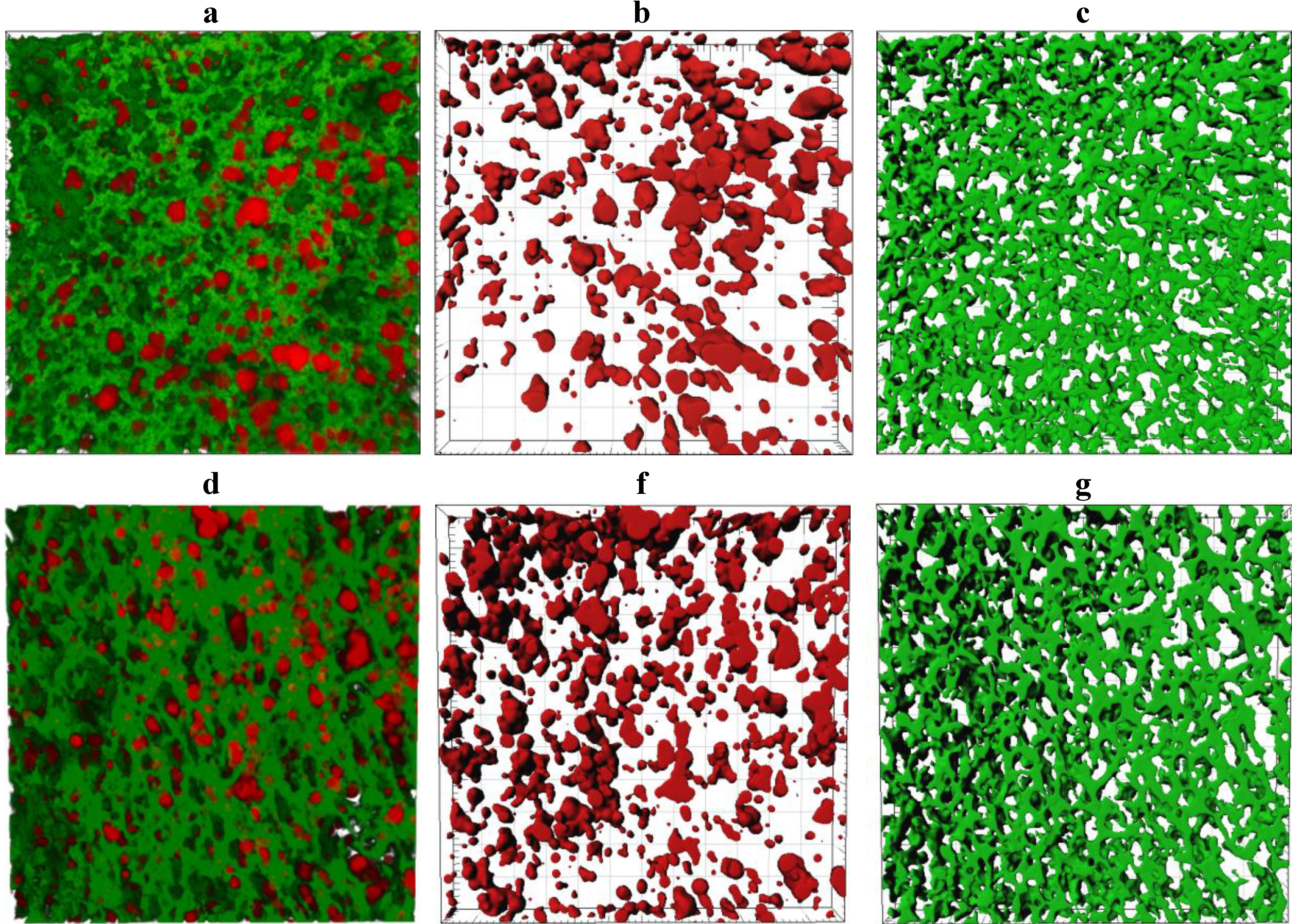
Representative CLSM images showing the microstructure of the cooked curd produced at a pilot scale are shown in
Figure 6. The number of fat globules decreased significantly (
p < 0.05) after cooking, possibly due to the coalescence of fat globules (
supplementary Figure S2a). The sphericity of the fat globules was reduced in some cases (
supplementary Figure S2b) and a concurrent increase in the size of the fat globules was observed (
supplementary Figure S2c,d). The porosity of the cooked curd samples was not significantly different (
p > 0.05) for different treatments but the protein strands within the cooked curd made with ~3.7% w/w milk protein (
Figure 6c) were qualitatively thinner and less dense than strands observed within cooked curd made with 5.8% w/w milk protein (
Figure 6f).
Figure 6.
CLSM microstructure of cooked curd made using unconcentrated cheese-milk with ~3.7% w/w milk protein (a–c) or cheese-milk standardized with LCUFR to 5.8% w/w milk protein (d–f). All images are 3D reconstructions consisting of 40 layers where the separation between layers is 0.25 µm giving a total observation depth of 10 µm. Nile red stained fat appears red and fast green stained protein appears green. Images b–f are the rendered volume of the CLSM images, where b and e show the rendered volume of the fat; c and f show the rendered volume of the protein. All scale bars are 20 µm in length.
Figure 6.
CLSM microstructure of cooked curd made using unconcentrated cheese-milk with ~3.7% w/w milk protein (a–c) or cheese-milk standardized with LCUFR to 5.8% w/w milk protein (d–f). All images are 3D reconstructions consisting of 40 layers where the separation between layers is 0.25 µm giving a total observation depth of 10 µm. Nile red stained fat appears red and fast green stained protein appears green. Images b–f are the rendered volume of the CLSM images, where b and e show the rendered volume of the fat; c and f show the rendered volume of the protein. All scale bars are 20 µm in length.
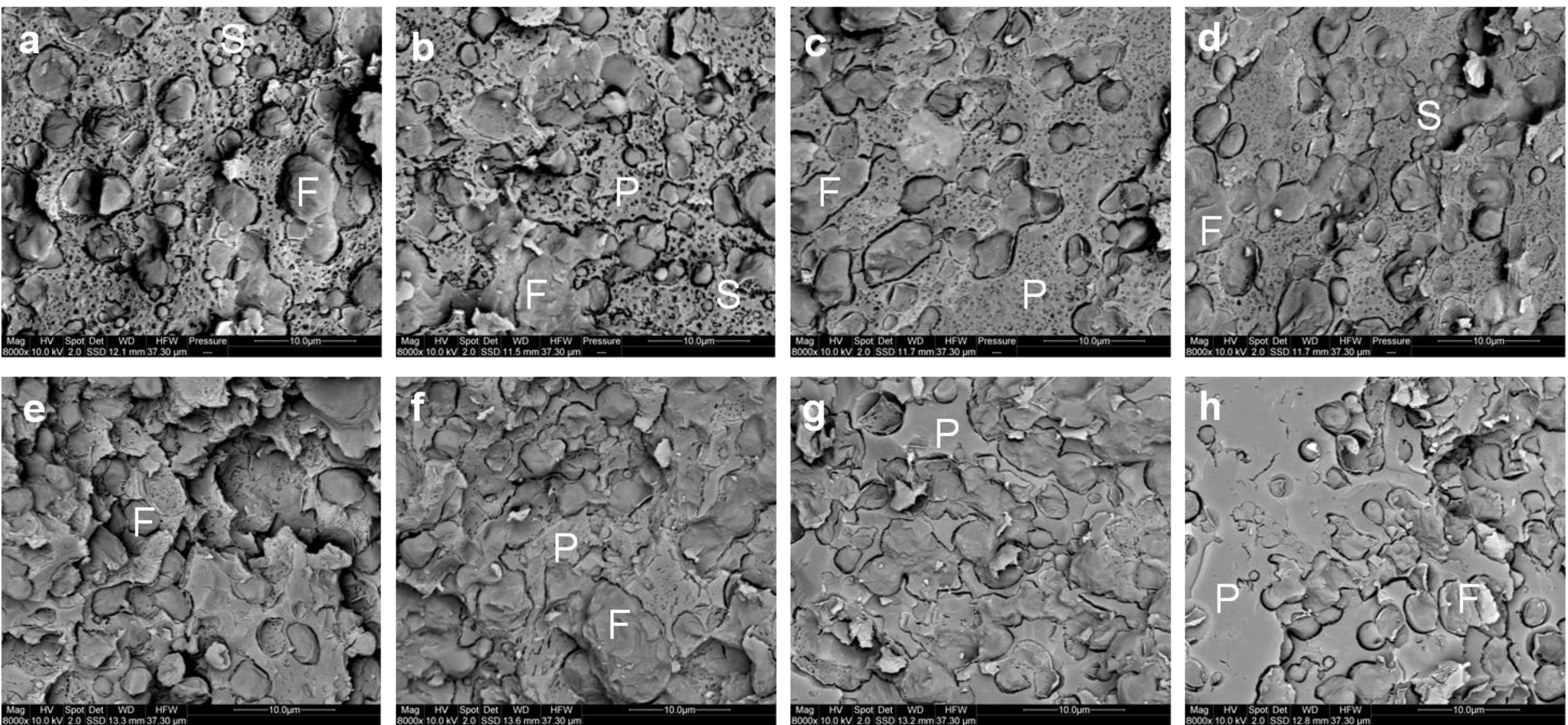
The microstructure of the curd after cheddaring and milling at a pilot scale observed by cryo SEM is shown in
Figure 7a–d. Micro-pores of about 1 µm diameter could readily be observed within the milled curd made with a lower concentration of milk protein (
Figure 7a,b). This was consistent with the higher moisture content of the lower protein cheeses (
Figure 3b). Conversely smaller micro-pores of diameter <1 µm were observed within the milled curd made from a higher concentration of milk protein (
Figure 7c,d). The starter culture bacteria could also be observed randomly within the curds formed in all treatments.
Figure 7.
Cryo SEM micrographs of the milled curd (a–d) and Cheddar cheese (e–h) made using (a,e) unconcentrated cheese-milk with ~3.7% w/w protein or cheese-milk standardized with LCUFR to (b,f) 4% w/w, (c,g) 4.8% w/w or (d,h) 5.8% w/w protein, where F = fat, P = protein network and S = starter culture bacteria. Scale bars within the images are 10 μm in length.
Figure 7.
Cryo SEM micrographs of the milled curd (a–d) and Cheddar cheese (e–h) made using (a,e) unconcentrated cheese-milk with ~3.7% w/w protein or cheese-milk standardized with LCUFR to (b,f) 4% w/w, (c,g) 4.8% w/w or (d,h) 5.8% w/w protein, where F = fat, P = protein network and S = starter culture bacteria. Scale bars within the images are 10 μm in length.
The micro-pores seen by cryo SEM (
Figure 7) could not readily be observed within the CLSM images and fewer differences between the protein treatments in the milled curd microstructure could be observed by CLSM, as also reflected in the data obtained by image analysis for these samples (
supplementary Figures S1 and S2).
The microstructure of the cheese after pressing is shown in
Figure 7e–h. Pressing completely closes the micro-pores observed earlier within the milled curd (
Figure 7a–d). Qualitatively, the microstructure observed by cryo SEM or CLSM for cheese made using the unconcentrated cheese-milk and cheese-milk with 5.8 % w/w protein were not different (
Figure 7e–h and
supplementary Figure S1). The size of the fat globules within cheeses made from cheese-milk with 5.8% w/w protein, however, was significantly bigger than in cheese made using unconcentrated milk (
supplementary Figure S2c,d). This difference may influence the flavour perception and texture of the cheese.
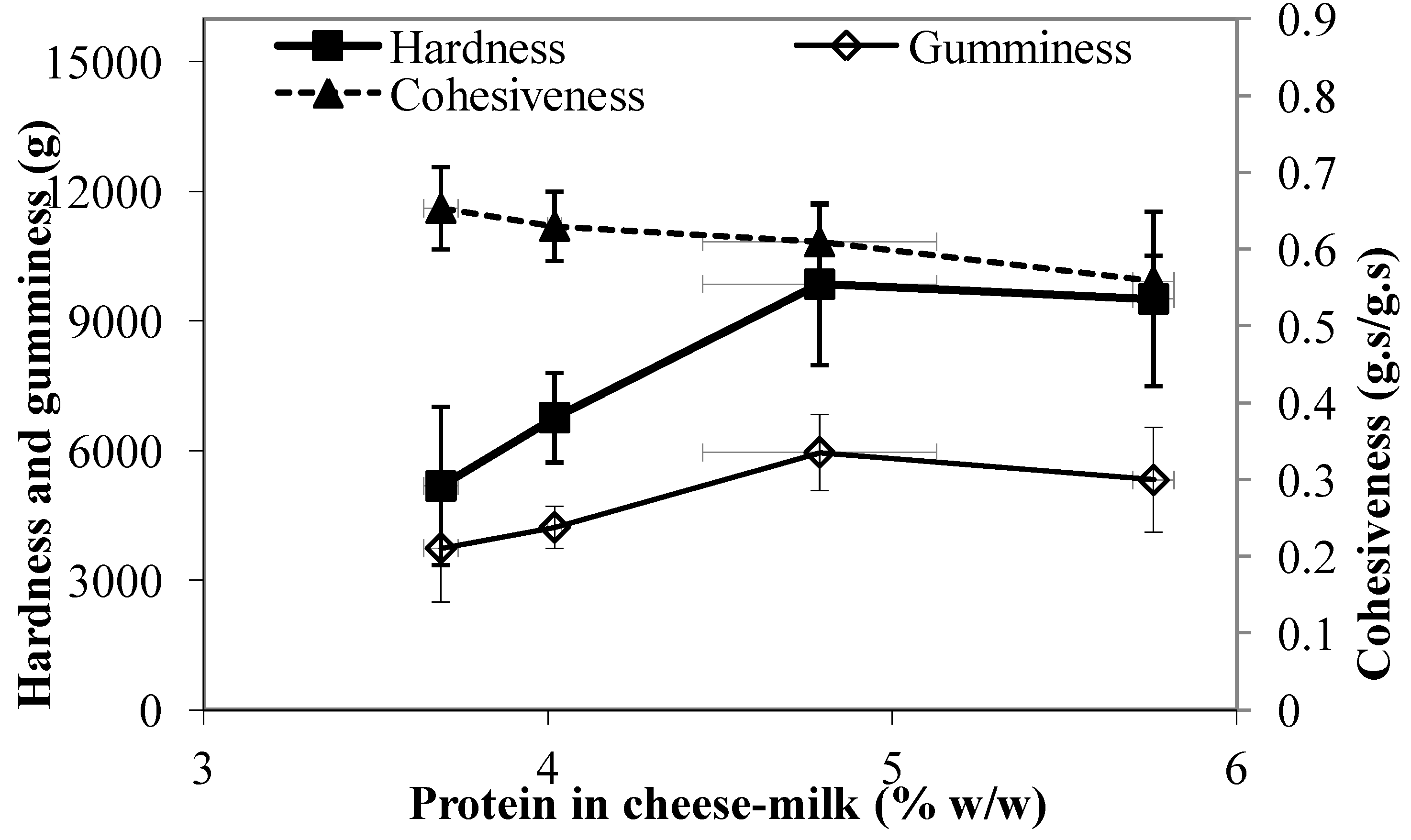
3.6. Texture Profile Analysis of the Cheese
The textural profile of the cheese measured within a week of cheese manufacture is shown in
Figure 8. The resistance to compression also known as the hardness, increased as the milk protein level increased (
p < 0.05) (
Figure 8). This result indicates that the cheese structure was more rigid, possibly due to the higher concentration of protein in the cheese-milk during gelation, the difference in microstructure or the lower moisture content of the cheese made using milk with greater protein. No significant measured differences in gumminess were observed (
Figure 8). Interestingly, the cohesiveness for cheese made from 5.8% w/w milk protein was significantly lower (
p < 0.05) than for cheese made from unconcentrated milk (
Figure 8). Further, we observed that cheeses made with 5.8% w/w milk protein crumbled more easily during the compression test.
Figure 8.
Texture profile analysis showing the hardness, gumminess, and cohesiveness of Cheddar cheeses made using unconcentrated cheese-milk with ~3.7% w/w milk protein or cheese-milk standardized with LCUFR to 4% w/w, 4.8% w/w or 5.8% w/w milk protein. Data are presented as mean ± standard deviation of mean (n = 3).
Figure 8.
Texture profile analysis showing the hardness, gumminess, and cohesiveness of Cheddar cheeses made using unconcentrated cheese-milk with ~3.7% w/w milk protein or cheese-milk standardized with LCUFR to 4% w/w, 4.8% w/w or 5.8% w/w milk protein. Data are presented as mean ± standard deviation of mean (n = 3).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







