1. Introduction
Olfactory cues of palatable food appear to work as appetizers while anticipating food intake. The smell of freshly baked bread entices you to buy and eventually eat a loaf.
From the day we are born we experience food odours during anticipation and consumption of food. Consumption is followed by nutritional consequences related to satiety, such as the digestion of available macronutrients. Over the course of our life we learn to use sensory food cues to (accurately) anticipate the energy density and taste (sweet/savoury) of the foods we are about to eat [
1,
2]. Through cephalic phase responses, our body is able to prepare for what will be ingested [
3,
4,
5]. Food odours thus are an important guide in our food-rich environment, but their exact role needs to be clarified.
Olfactory food cues presented in the anticipatory phase of eating are found to increase the appetite for congruent products and decrease the appetite for incongruent products [
6,
7]. This phenomenon is referred to as sensory-specific appetite. Besides product-specific effects, Ramaekers
et al. [
6,
7] found that savoury odours increased the appetite for (other) savoury foods and decreased the appetite for sweet foods, and
vice versa. In earlier research, exposure to food odours (pizza, cookies) increased appetite, liking and craving for the food that was smelled in restrained eaters [
8,
9]. Moreover, Gaillet
et al. [
10,
11] found that non-attentively perceived odours in the environment affected food choice. Participants placed in a waiting room with pear odour, chose fruity desserts more often compared to participants that had been waiting in an unscented room. Brief exposure to the smell and sight of pizza increased prospective intake for pizza and other savoury foods, but not for sweet foods [
12,
13]. After pizza cueing, the amount of pizza participants thought they could eat accurately predicted how much pizza they would actually eat [
12]. Fedoroff
et al. [
8,
9] and Larsen
et al. [
14] also found that participants ate more of an odour-cued food compared to non-cued food. Increased appetite likely parallels the effects of odour exposure on food choice, prospective intake and actual intake for matching products (
i.e, sweet/savoury, similar energy density, fruity).
The results of Ramaekers
et al. [
6,
7] indicate the presence of sensory-specific appetite for taste category (sweet
vs. savoury). Taste is important in the prediction of macronutrient content of a food. Savoury tastes are thought to indicate a high-protein content, whereas sweet tastes are suggested to point to a high-carbohydrate content, e.g., [
15,
16,
17]. Energy density of a food, such as fat content, is ecologically relevant as well. It has been suggested that humans are able to detect fat content of a food using their sense of smell [
18]. Anticipation of the energy density of a food is important in the process of energy-intake regulation.
Internal cues of hunger and satiety impact how much we eat [
19,
20]. It is likely that these internal cues also play a modulating role in food-cue reactivity during the anticipation phase of eating and thereby influence our appetite (responses) and drive to ingest energy.
In this study we aim to replicate the influence of olfactory cues on sensory-specific appetite for a certain taste category and extend those findings to energy-density categories of foods. Additionally, we are interested whether hunger state plays a modulatory role in this effect. We expect that for both taste and energy density, exposure to food odours will lead to an increased appetite for products that are congruent to the odour and a decreased appetite for products that are incongruent to the odour. Additionally, we hypothesize that sensory-specific effects on appetite are thought to be more relevant and thus more pronounced in a hungry compared to a satiated state.
3. Results
3.1. Hunger State (Manipulation Check)
Hunger ratings confirmed that feelings of hunger were significantly different between sessions, and according to the appropriate hunger states (hungry
vs. satiated; see
Table 2). In the hungry conditions the participants felt hungrier, less full, and indicated a higher prospective consumption and desire to eat (all
p < 0.001). Ratings of thirst indicated that participants were equally thirsty in both test sessions.
3.2. General Appetite
General appetite (100 mm VAS) differed significantly after exposure to different odours (p = 0.015) and also between hunger states (p < 0.001). General appetite after smelling chocolate (46 ± 3), beef (42 ± 4), melon (44 ± 3) and cucumber (47 ± 3) was significantly higher than after smelling fresh green (36 ± 3) or baseline reference (36 ± 3; all p < 0.01). General appetite in the hungry state (62 ± 3) was significantly higher compared to the satiated state (21 ± 3; p < 0.001).
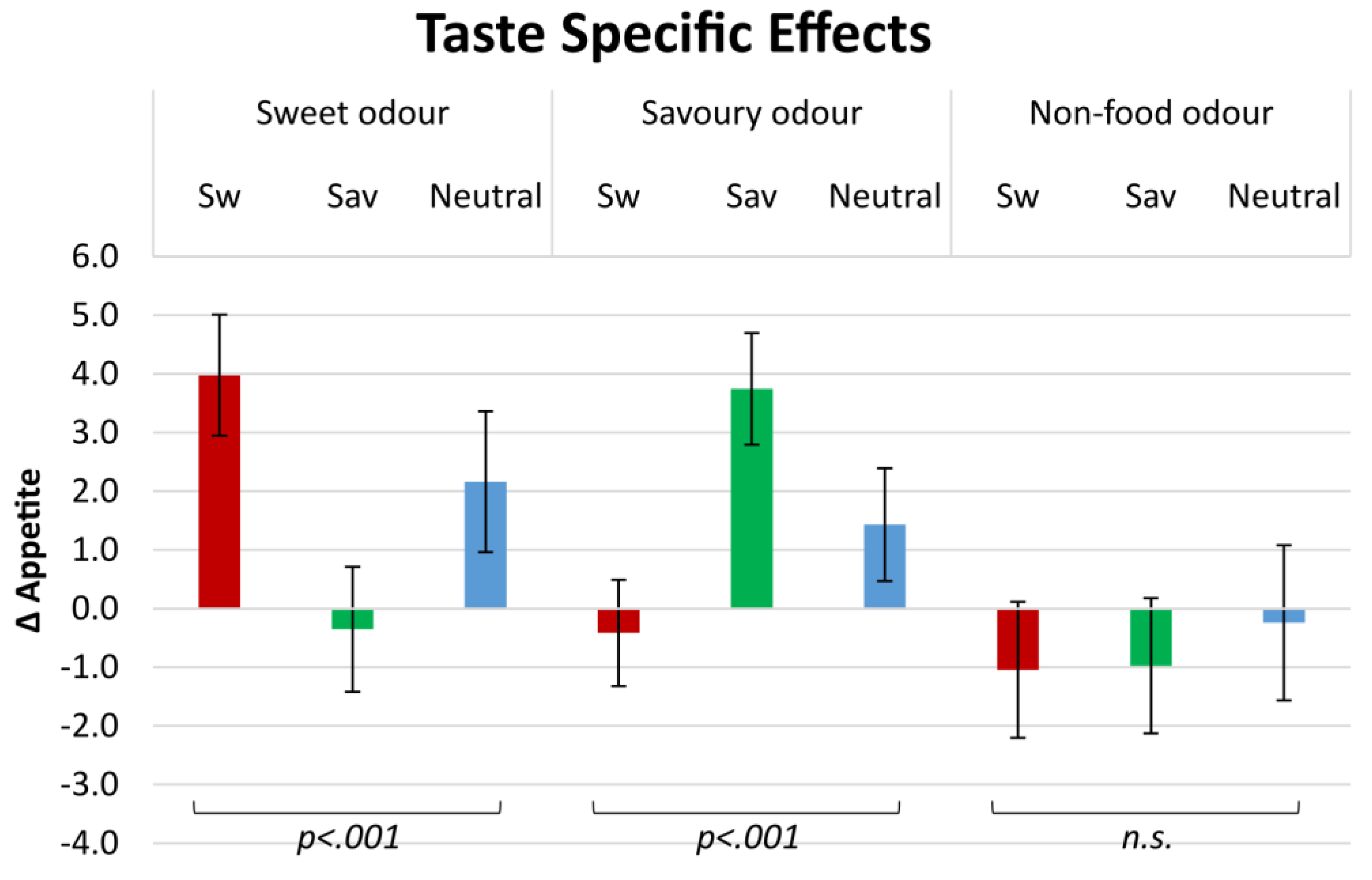
3.3. Sensory-Specific Appetite (SSA): Taste Category
Our results (see
Figure 1) show that by smelling Sw odours, appetite changes significantly for products of different taste categories (
F(2, 432) = 27.46,
p < 0.001). After exposure to Sw odours, Δ appetite was significantly higher for Sw products (4.0 ± 1.1) than for Sav (−0.4 ± 1.1;
p < 0.001) or neutral products (2.2 ± 1.2;
p = 0.022), and Δ appetite for Sav products was also significantly lower than appetite for neutral products (
p = 0.001).
Similarly, Δ appetite after smelling Sav odours was significantly different between the product-taste categories (F(2, 404) = 12.08, p < 0.001). It was higher for Sav products (3.7 ± 1.2) than for Sw (−0.4 ± 1.3; p < 0.001) or neutral products (1.4 ± 1.3; p = 0.003), but did not differ between Sw and neutral products (p = 0.017).
Δ Appetite after smelling an NF odour was not significantly different for the different products (F(2, 197) = 0.28, p = 0.750). For Sw products (−1.0 ± 0.9), Sav products (−1.0 ± 0.9) or neutral products (−0.2 ± 0.9; all p > 0.05).
In addition to an increase in general appetite when hungry (
Section 3.2), participants’ raw appetite ratings for products were higher in the hungry state (see
Figure S1). However, there were no significant differences in specific appetite (the difference between appetite after exposure to the no-odour control and appetite rated after exposure to an odour) between the hungry and satiated condition (difference in Δ appetite between hunger states after Sw odour: 0.7 mm; after Sav odour: 0.0 mm; after NF odour: 2.6 mm; all
p > 0.05).
In order to exclude the possibility that the effects described above were driven by the specific odour-product match (e.g., exposure to chocolate odour = appetite for chocolate pieces), these specific matches were excluded from the dataset and the analyses were rerun.
We found a significant main effect of Sw odour exposure on Δ appetite for different product taste categories (F(2, 403) = 19.29, p < 0.001). Δ Appetite after Sw odours remained significantly higher for Sw products (3.1 ± 1.1) than for Sav products (−0.4 ± 1.1; p < 0.001), but did not differ from Δ appetite for neutral products (2.0 ± 1.1; p = 0.191.). Further, Δ appetite for Sav products also differed from Δ appetite for neutral products (p = 0.003).
Δ Appetite after smelling Sav odours was also significantly different between product categories that were based on taste (F(2, 383) = 5.68, p = 0.004). Δ appetite for Sav products (2.8 ± 1.3) remained significantly higher than for Sw products (−0.4 ± 1.3; p < 0.001), but was not significantly different from Δ appetite for neutral products (1.2 ± 1.3; p = 0.052). Δ Appetite for Sw products was significantly different from Δ appetite for neutral products (p = 0.033).
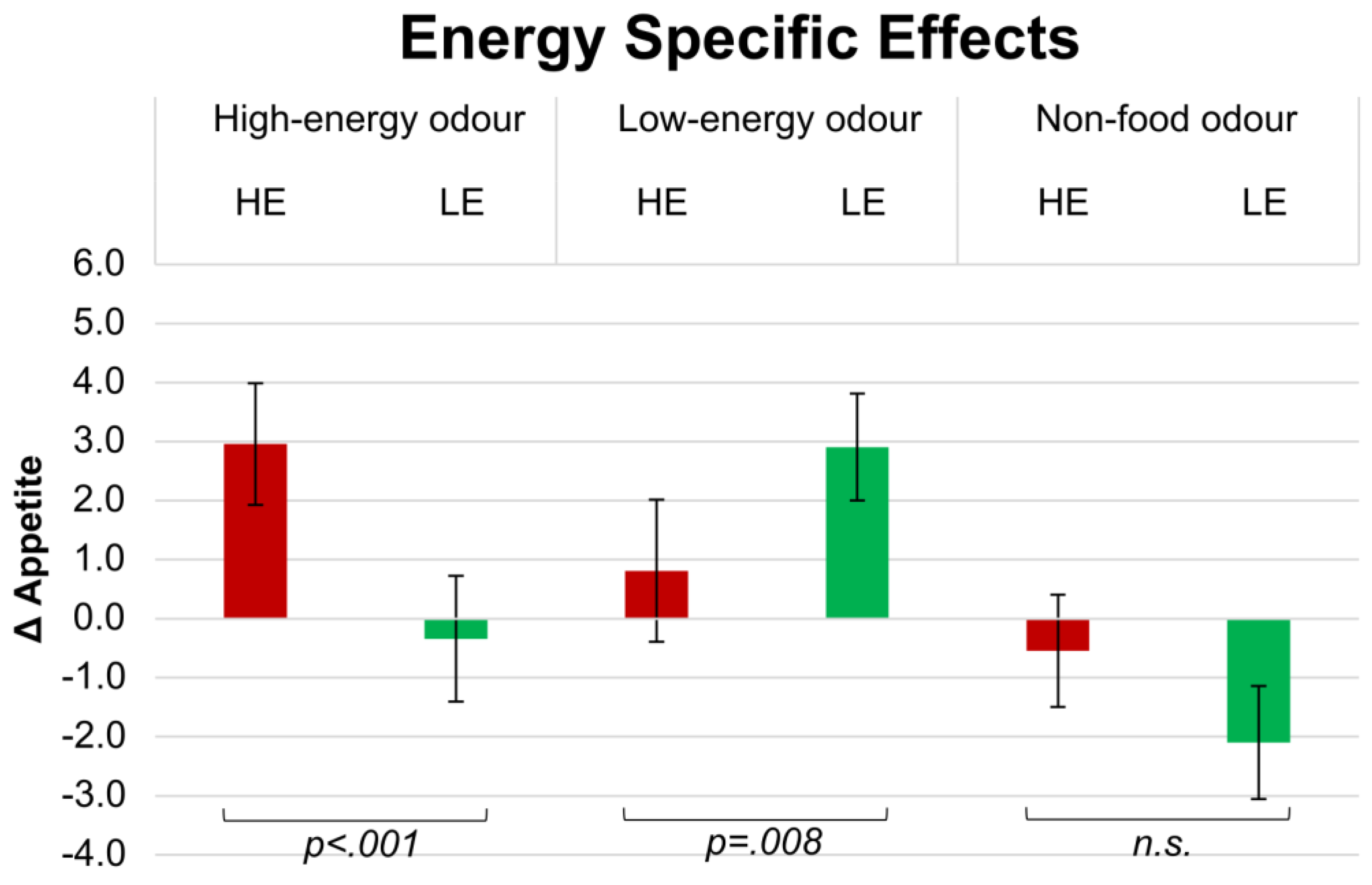
3.4. SSA: Energy Category
After smelling HE odours, Δ appetite for HE products (3.0 ± 1.1) was significantly higher than for LE (−0.3 ± 1.2;
p < 0.001; see
Figure 2).
Paired comparisons also revealed that Δ appetite rated after smelling LE odours was significantly higher for LE products (2.9 ± 1.2) than for HE products (0.8 ± 1.1; p = 0.008).
Δ Appetite rated after smelling an NF odour was not significantly different than for HE products (−0.5 ± 0.9) and LE products (−2.1 ± 1.1).
In
Figure S2 it is visible that raw appetite ratings for products are higher in the hungry state. Specific appetite (the difference between appetite after exposure to the no-odour control and appetite rated after exposure to an odour) was not significantly influenced by hunger state (difference in Δ appetite between hunger states after HE odour: 0.7 mm; after LE odour: 0.0 mm; both
p > 0.05).
As mentioned in
section 3.3, the analyses were rerun, excluding specific matches from the dataset, to account for the possibility that the effects described above were merely driven by the specific odour-product match.
Paired comparisons showed that after smelling HE odours, Δ appetite for HE products (2.3 ± 1.1) was significantly different from Δ appetite for LE products (−0.7 ± 1.1; p < 0.001).
Δ Appetite after smelling LE products was not different for LE products (1.8 ± 1.2) and HE products (0.6 ± 1.1).
4. Discussion
With this research, we wanted to determine whether odours signalling specific categories (sweet/savoury taste; high-/low-energy density; non-food) can induce sensory-specific changes in appetite. Furthermore, we were interested in whether the strength of this effect was dependent on hunger state. Results reveal that food odours increase appetite for products that are similar, both in taste and energy density. Hunger state did not significantly affect odour-induced sensory-specific appetite.
Our results indicate that odours that signal sweet products and odours that signal savoury products affect appetite in an opposite, but similar way. After smelling a sweet food odour, appetite for sweet products increases more compared to appetite for neutral (bread, croissant, and pancake) and savoury products. Similarly, after smelling a savoury food odour, appetite for savoury products increases significantly more than for neutral or sweet products. In line with this, the smell of flowers (fresh green), a non-food related odour, slightly decreased appetite ratings for sweet, savoury and neutral foods equally. These results are concurrent with those of Ramaekers
et al. [
6,
7]. We suggest that the level of congruency between the odour and the product determines the strength of the appetizing effect of the odours. Interestingly, after removing the specific odour-product matches (e.g., chocolate odour = appetite for chocolate pieces) from our dataset, appetite for taste-congruent products remained higher compared to non-congruent products. Olfactory sensory-specific appetite is a phenomenon that not only applies to a specific product, but also to categories of products related to taste quality (sweet
vs. savoury). By repeated exposure to the combination of an odour and the nutritional consequences after eating the food we smell, it is proposed that we learn to use (olfactory) food cues as predictors for macronutrient content [
1,
2]. These associations are most often described in terms of taste quality. Sweet taste is related to a high-carbohydrate content, whilst savoury taste is associated with a high-protein content [
15,
16,
24].
To our knowledge, this is the first study investigating olfactory sensory-specific appetite effects for energy density (high-energy vs. low-energy). When odours and products were categorized according to their associated energy density, we saw that appetite changed in a similar pattern as for taste category. Smelling odours associated with high-energy dense food increased the appetite for high-energy products significantly more than for low-energy dense products. The reverse occurred after smelling odours of low-energy dense foods. There was no difference in appetite for high-energy dense and low-energy dense products after smelling a non-food related odour. After removing the specific odour-product matches from the dataset, the pattern remained the same for high-energy dense odours. However, low-energy odours did not induce category-specific appetite effects. High-energy dense odours appear to be more potent appetizers than low-energy dense odours. Sensory-specific appetite seems to exist for specific odour-product matches and for the high energy-density category more in general.
When comparing these effects to the effects of the taste category, it seems that olfactory signals of taste lead to larger increases in appetite than for olfactory signals of energy density. After removal of the specific odour-product matches, an interesting pattern in the results emerges. Category-broad effects for sweet, savoury and high-energy dense odours remain, whereas a significant category-broad effect for low-energy food odours is absent. Based on these results, we speculate that odours signal macronutrient content. Low-energy food cues signal products that do not have substantial content. This can explain the lack of category-broad effect for low-energy cues. As mentioned for the taste category, it is thought that sweet signals carbohydrates (sweet) and savoury signals protein [
15,
16,
24]. On the contrary, the energy density of a food is not based on a single macronutrient
per se, but can be composed of different macronutrient sources, mainly fats and carbohydrates. In essence, teasing apart energy and macronutrient contribution is complex since they are interrelated. A systematic and more complete research approach is necessary to uncover the appetizing effects of olfactory stimuli representing proteins, carbohydrates and fats. Deprivation or supplementation of a certain macronutrient by an experimental diet could unveil whether appetizing effects of odours are modulated by macronutrient status. This will help to determine whether the function of macronutrient signalling can indeed be ascribed to olfactory cues in the anticipatory phase of eating.
Intake regulation is largely dependent on feelings of hunger and satiation [
25,
26]. Individuals that maintain their energy balance should be able to adjust their food intake according to their hunger status and resist eating beyond satiety when tempted by environmental food cues [
8]. Hunger ratings provided by the participants in our study confirmed that feelings of hunger (hunger, fullness, prospective consumption, desire to eat) were significantly different in the two hunger states (hungry
vs. satiated; see
Table 2). Surprisingly, hunger state did not modulate odour-induced specific appetite: Δ Appetite for specific products after odour exposure was no different when participants were satiated or not. The theory that tempting food cues in the environment stimulate eating in the absence of hunger has been posed before [
27]. This idea is supported by research by Nijs
et al. [
28], where participants showed a similar bias in orientation and maintained attention to visual food cues in both a hungry and satiated condition. Appetite responses of our participants also show that the impact of food odour exposure is independent of hunger state.
Selective detection of nutritious foods has been a beneficial trait for survival throughout evolution. We propose that in the anticipation phase of eating, (olfactory) food cues induce appetite specific to the macronutrient content that is signalled by the odour. Besides changes in appetite behaviour [
8,
9], previous research revealed very rapid physiological changes (e.g., salivation, endocrine responses; [
4,
29]) following food odour exposure. It is not clear whether food cues signalling a specific macronutrient content cause specific physiological responses to facilitate macronutrient uptake. Over the course of evolution, it would seem advantageous to have built-in systems (physiological, behavioural) that work together to obtain food sources in the environment.
Although olfactory food cues in our surroundings (ambient exposure) clearly have a specific effect on appetite, the effect on food preferences or choice is less consistent. Food odours do not seem to affect the preference for one type of food over another, as measured by a forced choice computer task [
19], but do influence what we decide to eat when choosing dishes from a menu [
10,
11]. These inconsistencies in choice are likely related to variations in context (controlled
vs. real-life). Ferriday and Brunstrom [
12,
13] showed that smelling and viewing pizza increases the amount of pizza we think we can eat (prospective consumption), a measure closely related to appetite. This effect also transferred to dishes that were similar to pizza (“scrambled egg, chips and baked beans”, “pasta and tomato sauce”), but not to dishes that were dissimilar (cake).What is more, after smelling a food (in combination with the sight), participants ate more of the cued food [
8,
9,
12,
14]. However, findings on intake are not consistently repeated [
19] and seem to have specific prerequisites with regard to the context, cue exposure and also personality characteristics (restraint, impulsivity). Altogether, it seems that olfactory food cues have a more clear-cut role in the phase leading up to meal initiation [
30], tempting us to start eating what is in front of us. Their influence may wane in later phases, when other cues such as flavour (taste, retronasal odour), satiety signals and intake-inhibiting behaviours come into play, that work toward cessation of eating. In the context of food scarcity and low food security, it is highly beneficial to have the ability to detect nutritious food sources. Subsequent increases in appetite and changes in physiology may promote and facilitate adequate food intake and uptake, thereby increasing chances of survival. In most Western societies, however, energy-dense foods are readily available. In this case, appetizing olfactory food cues are part of an environment that promotes overconsumption, ultimately contributing to a higher incidence of nutrition-related diseases (e.g., obesity, diabetes).
We cannot exclude that our sample size was too small to detect modulatory effects of hunger state on specific appetite. However, we do not find this plausible since significant effects of hunger state on general appetite were found. Confirmation of the absence of this effect may be provided by future research in a larger study population. Additionally, in the current study we only included females, limiting the possibility to generalize our findings to a broader population that includes males.





{kind=link}
{kind=link}