Autochthonous and Probiotic Lactic Acid Bacteria Employed for Production of “Advanced Traditional Cheeses”

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
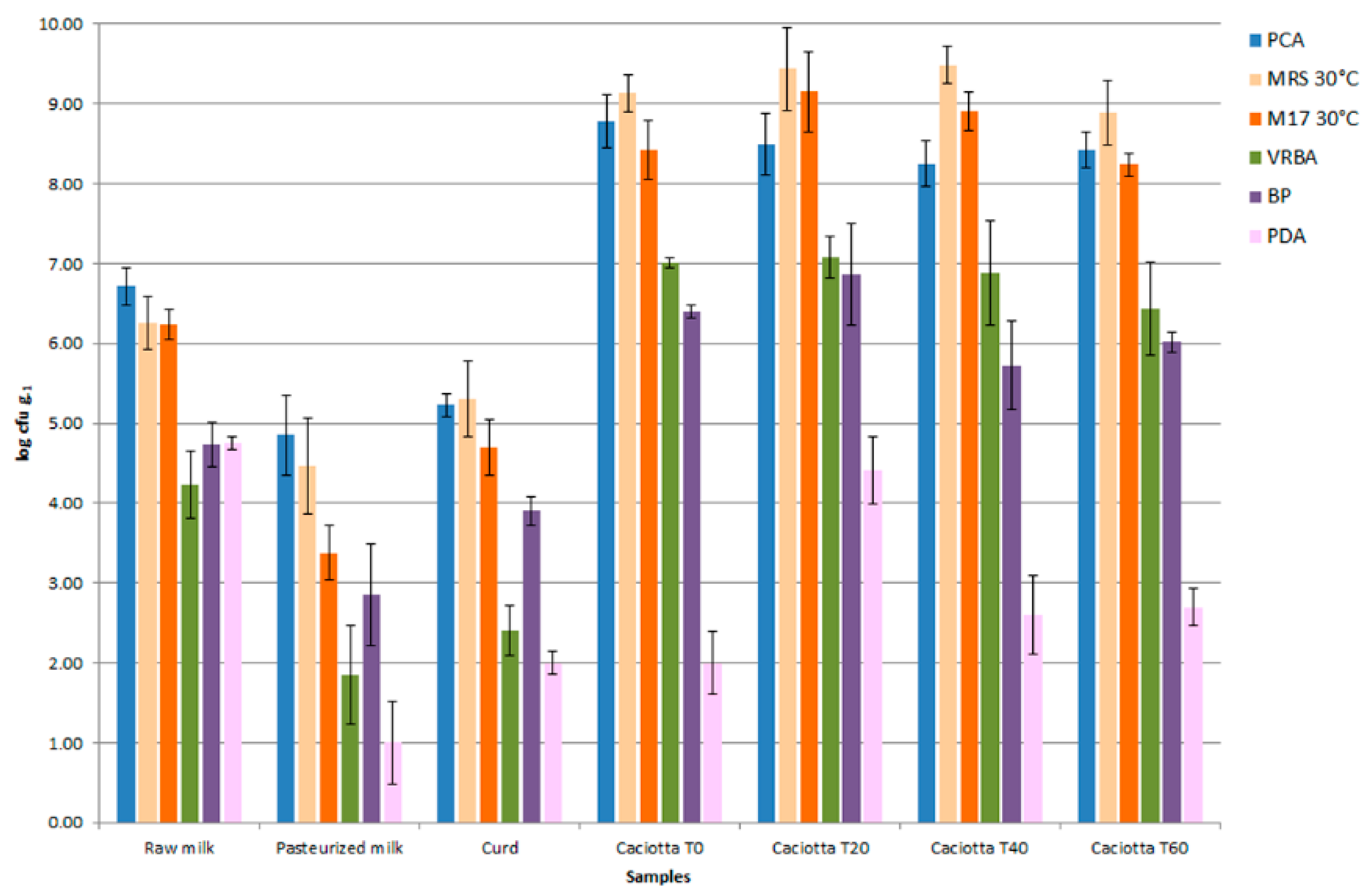
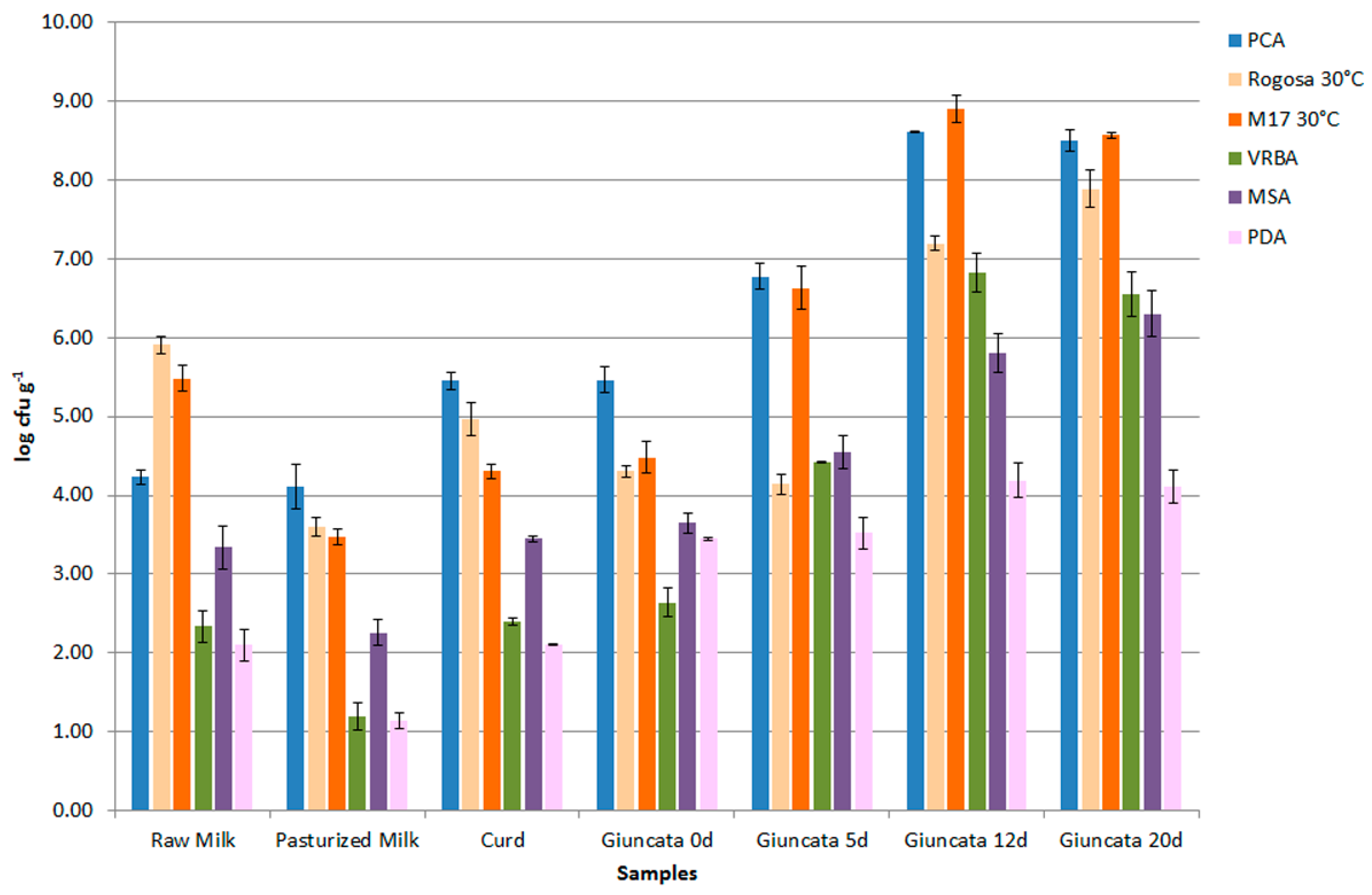
2.1. Cheese-Making and Sampling
2.2. Enumeration and Isolation of Microorganisms and Preliminary Characterization of Isolates
2.3. Genomic DNA Extraction and Molecular Analysis of Isolates
2.4. Technological Characterization of Bacterial Strains
2.5. Pilot-Scale Cheese-Making of Giuncata and Caciotta Leccese with Autochthonous and Probiotic Strains
2.6. Sensory Analysis
2.7. Advanced Cheese Feeding Procedure
3. Results and Discussion
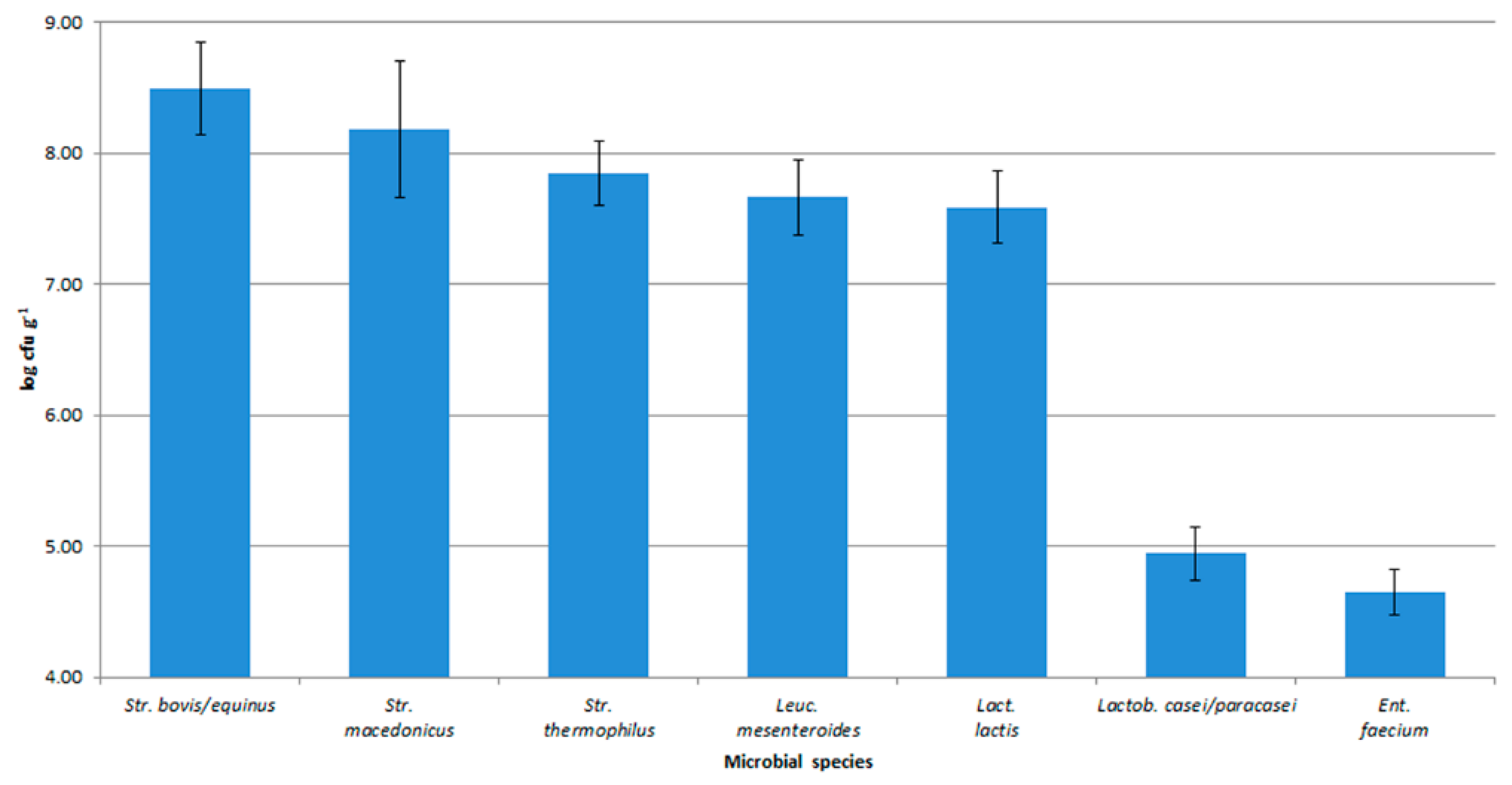
3.1. Isolation and Molecular Characterization of Microorganisms from Cheeses
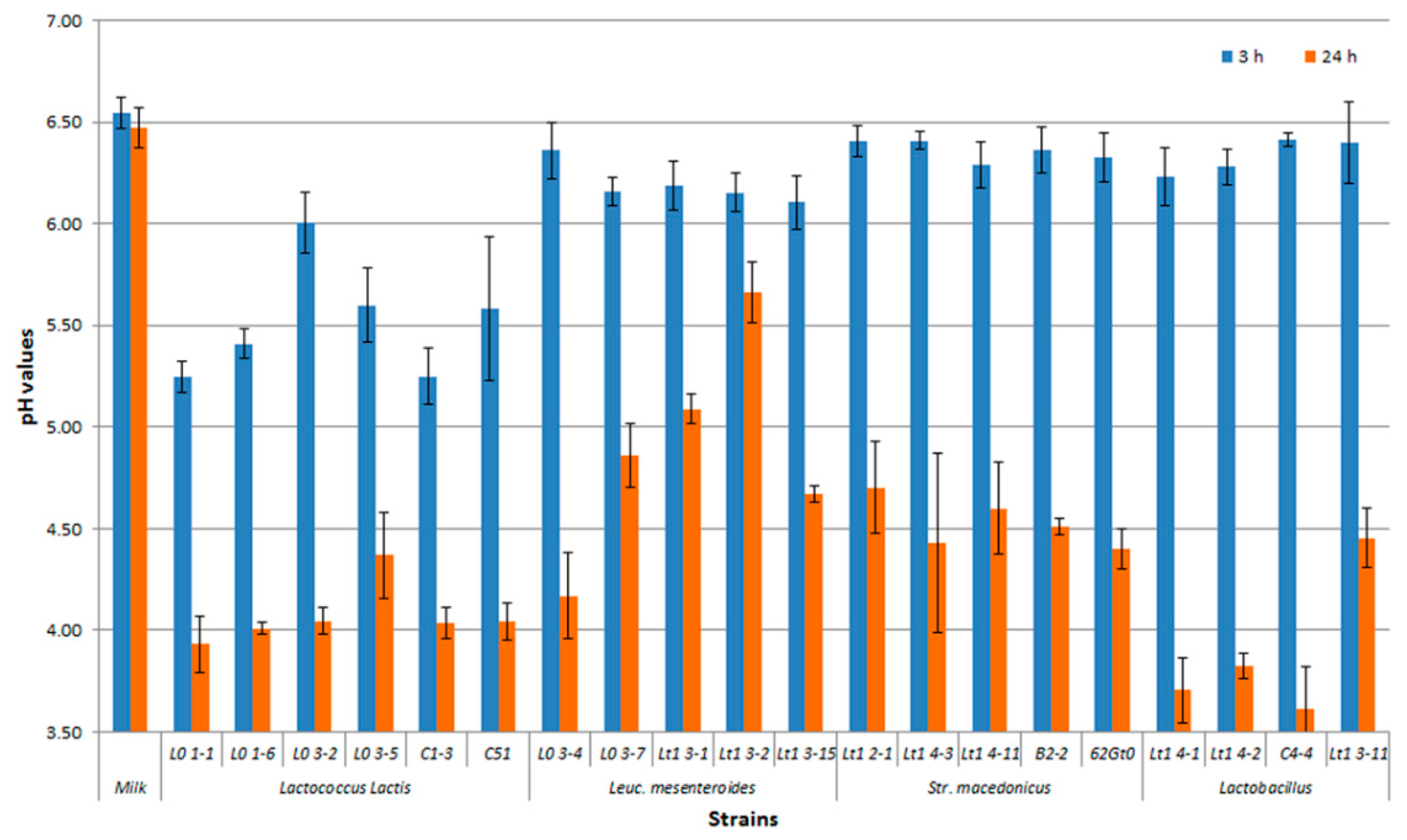
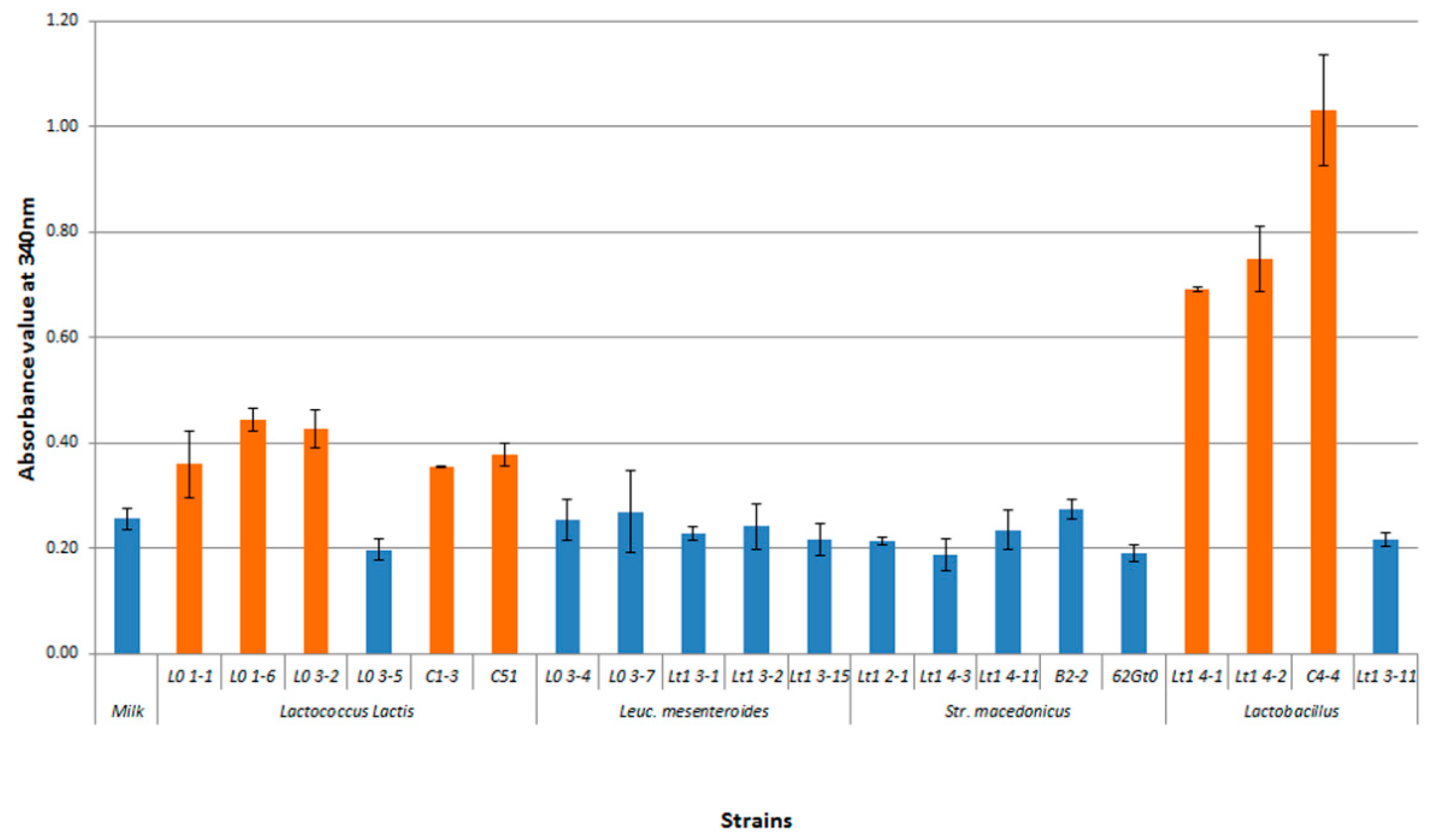
3.2. Technological Characterization of Bacterial Strains
3.3. Experimental Pilot-Scale Making of Giuncata and Caciotta Amended with Pro-Technological Strains and with the Probiotic Strain
3.4. Sensory Analyses
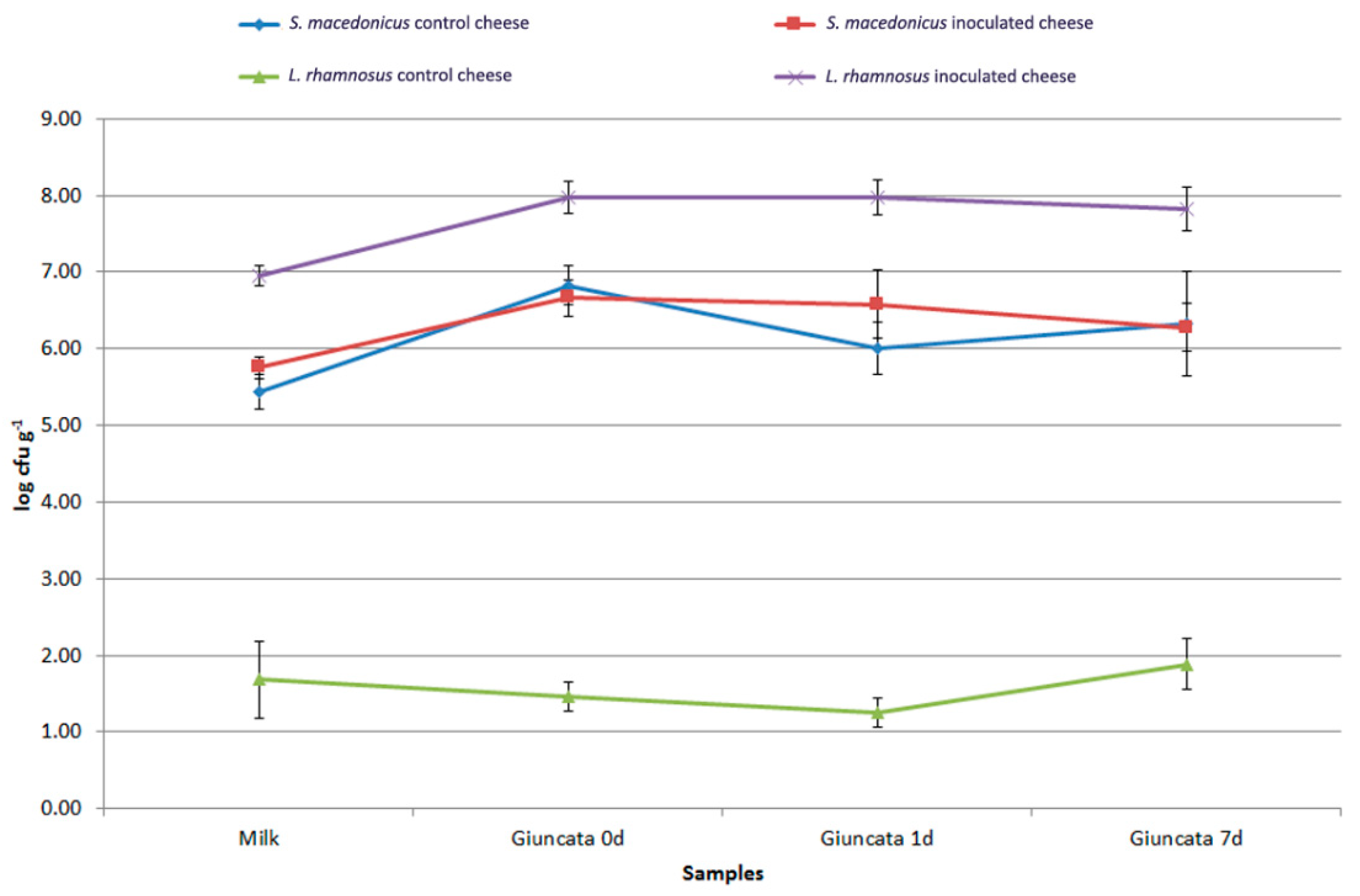
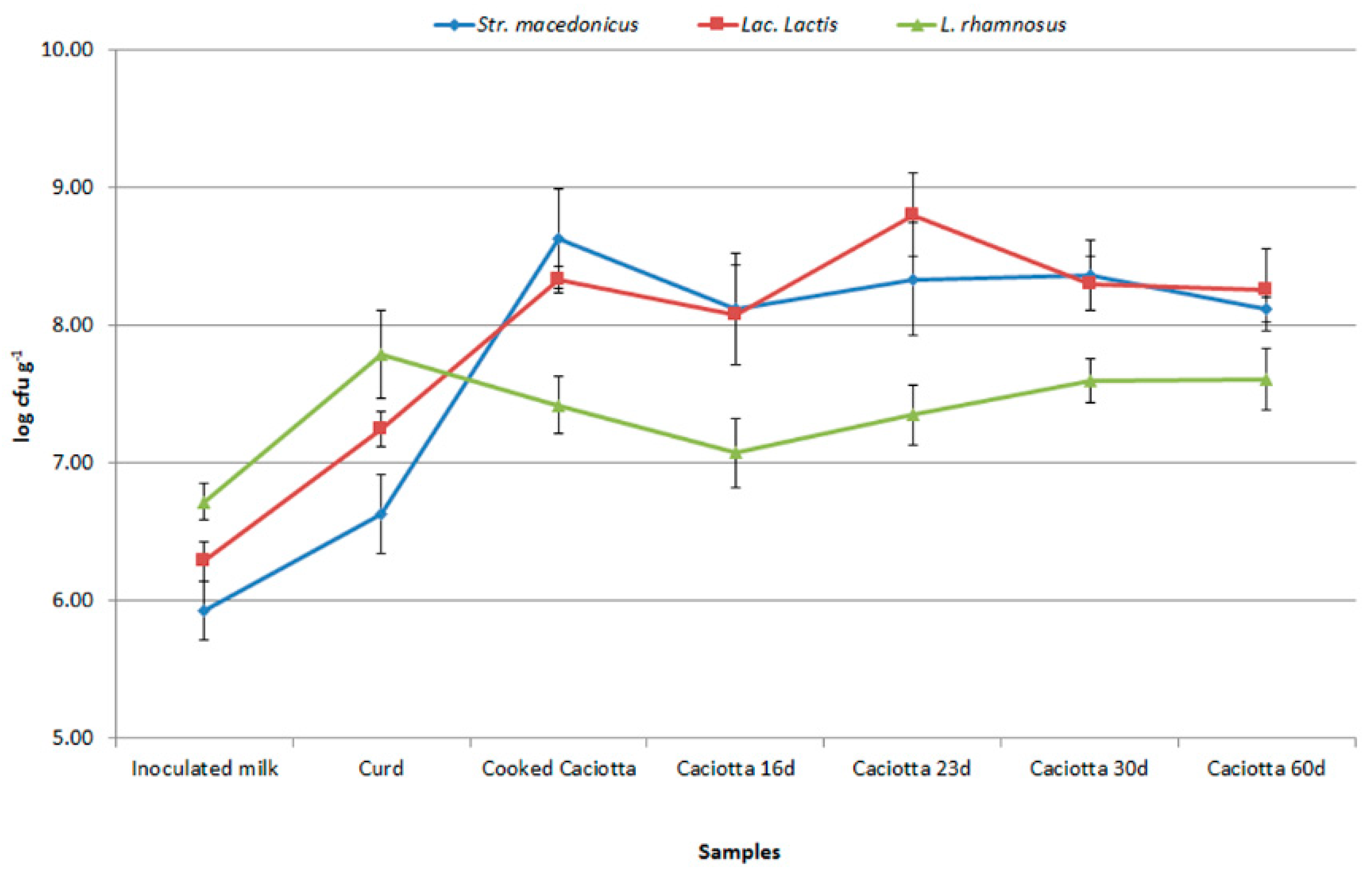
3.5. Persistence of Pro-Technological and Probiotic Strains
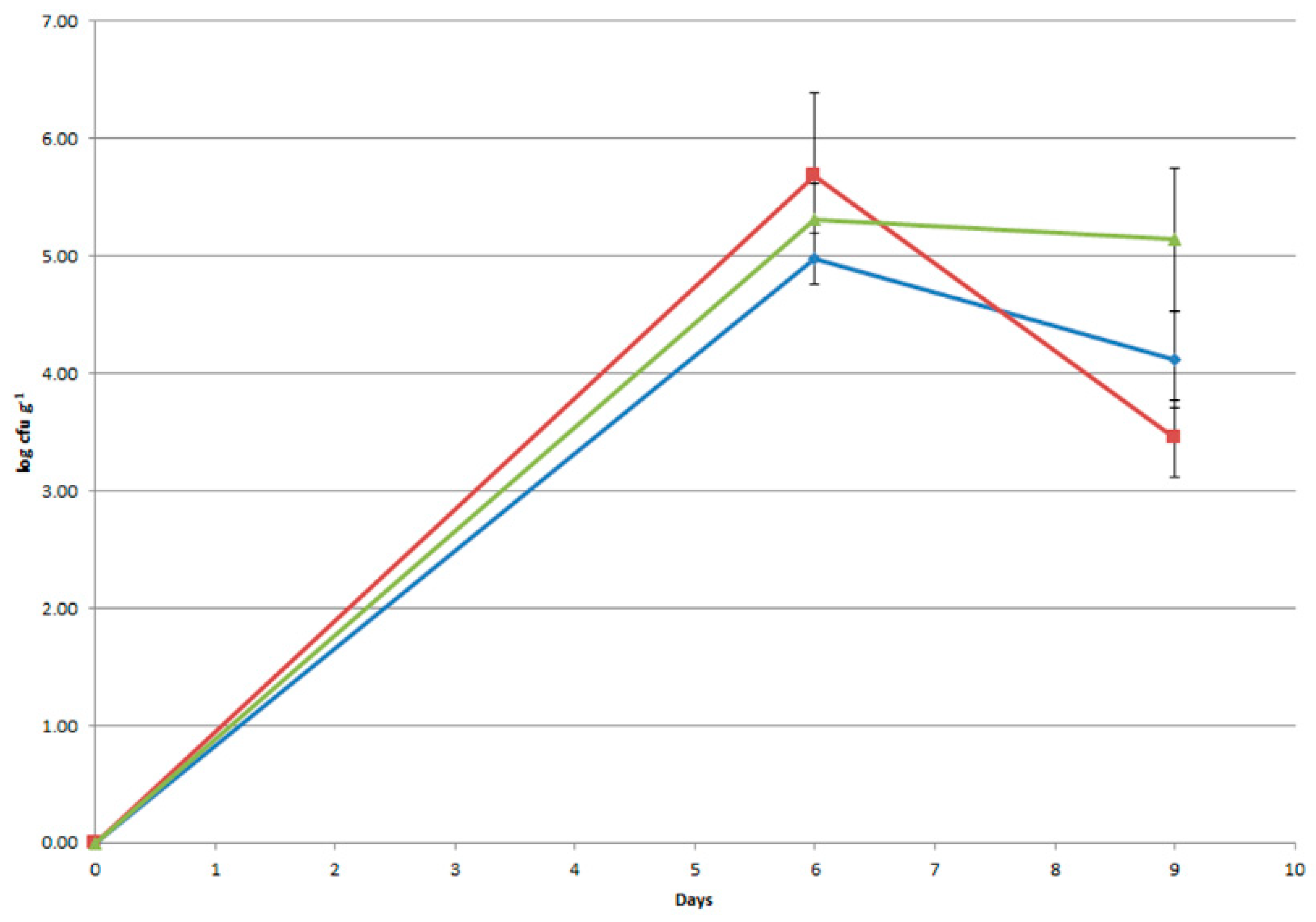
3.6. Monitoring of the Probiotic Strain in Human Trials
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Di Cagno, R.; De Pasquale, I.; De Angelis, M.; Buchin, S.; Calasso, M.; Fox, P.F.; Gobbetti, M. Manufacture of Italian Caciotta-type cheeses with adjuncts and attenuated adjuncts of selected non-starter lactobacilli. Int. Dairy J. 2011, 21, 254–260. [Google Scholar] [CrossRef]
- Olarte, C.; Gonzalez-Fandos, E.; Gimenez, M.; Sanz, S.; Portu, J. The growth of Listeria monocytogenes in fresh goat cheese (Cameros cheese) packaged under modified atmospheres. Food Microbiol. 2002, 19, 75–82. [Google Scholar] [CrossRef]
- De Angelis, M.; de Candia, S.; Calasso, M.P.; Faccia, M.; Guinee, T.P.; Simonetti, M.C.; Gobbetti, M. Selection and use of autochthonous multiple strain cultures for the manufacture of high-moisture traditional Mozzarella cheese. Int. J. Food Microbiol. 2008, 125, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Morea, M.; Matarante, A.; Di Cagno, R.; Baruzzi, F.; Minervini, F. Contribution of autochthonous non-starter lactobacilli to proteolysis in Caciocavallo Pugliese cheese. Int. Dairy J. 2007, 17, 525–534. [Google Scholar] [CrossRef]
- Quero, G.M.; Fusco, V.; Cocconcelli, P.S.; Owczarek, L.; Borcakli, M.; Fontana, C.; Skapska, S.; Jasinska, U.T.; Ozturk, T.; Morea, M. Microbiological, physico-chemical, nutritional and sensory characterization of traditional Matsoni: Selection and use of autochthonous multiple strain cultures to extend its shelf-life. Food Microbiol. 2014, 38, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Di Lena, M.; Quero, G.M.; Santovito, E.; Verran, J.; De Angelis, M.; Fusco, V. A selective medium for isolation and accurate enumeration of Lactobacillus casei-group lactobacilli in probiotic milks and dairy products. Int. Dairy J. 2015, 47, 27–36. [Google Scholar] [CrossRef]
- Milk and Milk Products. Guidance on Methods of Sampling; Standard 50C:1995; International Dairy Federation, Ed.; IDF: Brussels, Belgium, 1995. [Google Scholar]
- ISO (International Organization for Standardization) 8261:2001. Milk and Milk-Products—General Guidance for the Preparation of Test Samples, Initial Suspensions and Decimal Dilutions for Microbiological Examination; International Organization for Standardization: Geneva, Switzerland, 2001. [Google Scholar]
- Sperber, W.H.; Swan, J. Hot loop test for the determination of carbon dioxide production from glucose by lactic acid bacteria. Appl. Environ. Microbiol. 1976, 31, 990–991. [Google Scholar]
- Fusco, V.; Quero, G.M.; Morea, M.; Blaiotta, G.; Visconti, A. Rapid and reliable identification of Staphylococcus aureus harbouring the enterotoxin gene cluster (egc) and quantitative detection in raw milk by real time PCR. Int. J. Food Microbiol. 2011, 144, 528–537. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Morea, M.; Stea, G.; Visconti, A. Novel PCR-based identification of Weissella confusa using an AFLP-derived marker. Int. J. Food Microbiol. 2011, 145, 437–443. [Google Scholar] [CrossRef]
- Jensen, M.A.; Webster, J.A.; Straus, N. Rapid identification of bacteria on the basis of polymerase chain reaction-amplified ribosomal DNA spacer polymorphisms. Appl. Environ. Microbiol. 1993, 59, 945–952. [Google Scholar] [PubMed]
- Ercolini, D.; Fusco, V.; Blaiotta, G.; Coppola, S. Sequence heterogeneity in the lacSZ operon of Streptococcus thermophilus and its use in PCR systems for strain differentiation. Res. Microbiol. 2005, 156, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Papadelli, M.; Manolopoulou, E.; Kalantzopoulos, G.; Tsakalidou, E. Rapid detection and identification of Streptococcus macedonicus by species-specific PCR and DNA hybridisation. Int. J. Food Microbiol. 2003, 81, 231–239. [Google Scholar] [CrossRef]
- Coppola, S.; Fusco, V.; Andolfi, R.; Aponte, M.; Blaiotta, G.; Ercolini, D.; Moschetti, G. Evaluating microbial diversity during the manufacture of “fior di latte di agerola”, a traditional raw milk cheese of Naples area. J. Dairy Res. 2006, 73, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Aponte, M.; Fusco, V.; Andolfi, R.; Coppola, S. Lactic acid bacteria occurring during manufacture and ripening of Provolone del Monaco cheese: Detection by different analytical approaches. Int. Dairy J. 2008, 18, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Blaiotta, G.; Fusco, V.; Ercolini, D.; Aponte, M.; Pepe, O.; Villani, F. Diversity of Lactobacillus strains based on partial HSP60 gene sequences and design of PCR-RFLP assays for species identification and differentiation. Appl. Environ. Microbiol. 2008, 74, 208–215. [Google Scholar] [CrossRef]
- Baruzzi, F.; Poltronieri, P.; Quero, G.M.; Morea, M.; Morelli, L. An in vitro protocol for direct isolation of potential probiotic lactobacilli from raw bovine milk and traditional fermented milks. Appl. Microbiol. Biotechnol. 2011, 90, 331–342. [Google Scholar] [CrossRef]
- Dabour, N.; LaPointe, G. Identification and molecular characterization of the chromosomal exopolysaccharide biosynthesis gene cluster from Lactococcus lactis subsp. cremoris SMQ-461. Appl. Environ. Microbiol. 2005, 71, 7414–7425. [Google Scholar] [CrossRef]
- Mozzi, F.; Torino, M.I.; de Valdez, G.F. Identification of Exopolysaccharide-Producing Lactic Acid Bacteria; Food Microbiology Protocols; Humana Press: Totowa, NJ, USA, 2001; pp. 183–190. [Google Scholar]
- Kempler, G.M.; McKay, L.L. Improved medium for detection of citrate-fermenting Streptococcus lactis subsp. diacetylactis. Appl. Environ. Microbiol. 1980, 39, 926–927. [Google Scholar]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Baruzzi, F.; Matarante, A.; Morea, M.; Cocconcelli, P.S. Microbial community dynamics during the Scamorza Altamurana cheese natural fermentation. J. Dairy Sci. 2002, 85, 1390–1397. [Google Scholar] [CrossRef]
- Lamy, F.; Galaup, F. Compte rendu des communications, Carhaix. In Colloqui et Tournee Pratique Sur la Lipolyse du Lait; GIE: Louisville, KY, USA, 1986; pp. 9–12. [Google Scholar]
- Tagg, J.R.; McGiven, A.R. Assay system for bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [PubMed]
- The Disk Diffusion Method Developed by the British Society for Antimicrobial Chemotherapy. Available online: http://www.bsac.org.uk/eucastbsac-disc-diffusion-method (accessed on 21 July 2019).
- Tosi, L.; Berruti, G.; Danielsen, M.; Wind, A.; Huys, G.; Morelli, L. Susceptibility of Streptococcus thermophilus to antibiotics. Antonie Leeuwenhoek 2007, 92, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Pacini, F.; Cariolato, D.; Andrighetto, C.; Lombardi, A. Occurrence of Streptococcus macedonicus in Italian cheeses. FEMS Microbiol. Lett. 2006, 261, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.J.H.; Timmins, M.J. Differentiation of Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus by polymerase chain reaction. Lett. Appl. Microbiol. 1999, 29, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, E.; Settanni, L.; Carlin, S.; Cavazza, A.; Poznanski, E. A factory-scale application of secondary adjunct cultures selected from lactic acid bacteria during Puzzone di Moena cheese ripening. J. Dairy Sci. 2008, 91, 2981–2991. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, H.J.M.; Gibson, G.R.; Elfferich, P.; Raangs, G.C.; Wildeboer-Veloo, A.C.M.; Argaiz, A.; Roberfroid, M.B.; Welling, G.W. Comparison of viable cell counts and fluorescence in situ hybridization using specific rRNA-based probes for the quantification of human fecal bacteria. FEMS Microbiol. Lett. 2000, 183, 125–129. [Google Scholar] [CrossRef]
- Flórez, A.B.; Mayo, B. Microbial diversity and succession during the manufacture and ripening of traditional, Spanish, blue-veined Cabrales cheese, as determined by PCR-DGGE. Int. J. Food Microbiol. 2006, 110, 165–171. [Google Scholar] [CrossRef]
- Fuka, M.M.; Wallisch, S.; Engel, M.; Welzl, G.; Havranek, J.; Schlotera, M. Dynamics of bacterial communities during the ripening process of different Croatian cheese types derived from raw ewe’s milk cheeses. PLoS ONE 2013, 8, e80734. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | RM | PM | Samples 1 | Total | ||||
|---|---|---|---|---|---|---|---|---|
| C | G | G5d | G12d | G20d | ||||
| Lactococcus lactis | 6 (−3) | 7 (−4) | 6 (−3) | 1(−3) | 12 (−5) | 4 (−6) | 35 | |
| subsp. lactis | ||||||||
| Lactobacillus casei | 1 (−3) | 1 | ||||||
| Streptococcus parauberis | 1 (−3) | 2 (−3) | 10 (−3) | 3( −5) | 2 (−5) | 18 | ||
| Streptococcus thermophilus | 1 (−3) | 12 (−2) | 3 (−5) | 11 (−4) | 10 (−3) | 2 (−3) | 39 | |
| Streptococcus macedonicus | 2 (−3) | 1 (−3) | 3 (−6) | 12 (−4) | 11 (−3) | 2 (−3) | 31 | |
| Leuconostoc mesenteroides | 1 (−3) | 3 (−6) | 4 | |||||
| subsp. mesenteroides | ||||||||
| Enterococcus faecium | 1 (−3) | 1 (−3) | 2 | |||||
| Streptococcus bovis | 1 (−3) | 1 (−3) | 2 | |||||
| Species | Strain | Source |
|---|---|---|
| Lactococcus lactis | L0 1-1 | Caciotta 24 h after salting |
| L0 1-6 | Caciotta 24 h after salting | |
| L0 3-2 | Caciotta 24 h after salting | |
| L0 3-5 | Caciotta 24 h after salting | |
| C51 | Giuncata curd | |
| Leuconostoc mesenteroides | L0 3-4 | Caciotta 24 h after salting |
| L0 3-7 | Caciotta 24 h after salting | |
| Lt1 3-1 | 20 days ripened Caciotta | |
| Lt1 3-2 | 20 days ripened Caciotta | |
| Lt1 3-15 | 20 days ripened Caciotta | |
| Streptococcus macedonicus | Lt1 2-1 | 20 days ripened Caciotta |
| Lt1 4-3 | 20 days ripened Caciotta | |
| Lt1 4-11 | 20 days ripened Caciotta | |
| B2-2 | Thermized milk | |
| 62Gt0 | Ready produced Giuncata | |
| Streptococcus thermophilus | Lt1 2-5 | 20 days ripened Caciotta |
| B2-3 | Thermized milk | |
| Lactobacillus delbrueckii | Lt1 4-1 | 20 days ripened Caciotta |
| Lt1 4-2 | 20 days ripened Caciotta | |
| Lactobacillus paracasei | Lt1 3-11 | 20 days ripened Caciotta |
| Trial | S. macedonicus | Lc. lactis |
|---|---|---|
| Control (C) | ||
| Fresh Giuncata Leccese (T0) * | <100 | <100 |
| Fresh Giuncata Leccese (T1) ** | <100 | <100 |
| Fresh Giuncata Leccese (T4) *** | <100 | <100 |
| Str. macedonicus 62GT0 (ASC2) | ||
| Fresh Giuncata Leccese (T0) | 6.95 × 106 | |
| Fresh Giuncata Leccese (T1) | 1.37 × 107 | |
| Fresh Giuncata Leccese (T4) | 4.95 × 107 | |
| Lc. lactis LC51 (ASC1) | ||
| Fresh Giuncata Leccese (T0) | 9.63 × 108 | |
| Fresh Giuncata Leccese (T1) | 2.30 × 109 | |
| Fresh Giuncata Leccese (T4) | 6.45 × 109 | |
| Lc. lactis LC51 + Str. macedonicus 62GT0 (ASC3) | ||
| Fresh Giuncata Leccese (T0) | 1.30 × 107 | 9.49 × 108 |
| Fresh Giuncata Leccese (T1) | 2.32 × 107 | 2.23 × 109 |
| Fresh Giuncata Leccese (T4) | 8.14 × 107 | 6.30 × 109 |
| Assessors | Sensory Descriptors | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flat | Sweet | Acidic | Bitter | Creamy | Soft | Gummy | Hard | |||||||||
| CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | |
| 1 | ± | - | ± | ± | - | ± | ± | - | + | + | + | + | - | - | - | - |
| 2 | - | - | ± | - | - | - | - | - | + | + | + | + | - | - | - | - |
| 3 | ± | - | ± | ± | - | ± | ± | - | + | + | + | + | - | - | - | - |
| 4 | - | - | ± | ± | - | - | - | - | + | + | + | + | - | - | - | - |
| 5 | ± | - | + | - | - | ± | ± | - | + | + | + | + | - | - | - | - |
| 6 | ± | ± | + | - | ± | + | - | - | + | + | + | + | - | - | - | - |
| Assessors | Sensory Descriptors | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flat | Sweet | Acidic | Bitter | Creamy | Soft | Gummy | Hard | |||||||||
| CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | CC | ASC | |
| 1 | - | ± | - | - | - | - | ± | - | ± | + | + | + | - | - | - | - |
| 2 | - | ± | - | - | ± | - | - | - | - | + | - | + | ± | - | - | - |
| 3 | - | + | - | - | - | - | ± | - | - | + | - | + | ± | - | - | - |
| 4 | - | ± | - | - | - | - | - | - | ± | ++ | + | + | - | - | - | - |
| 5 | - | ± | - | - | - | - | ± | - | - | + | + | + | - | - | - | - |
| 6 | - | + | - | - | ± | - | - | - | - | + | - | + | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, V.; Quero, G.M.; Poltronieri, P.; Morea, M.; Baruzzi, F. Autochthonous and Probiotic Lactic Acid Bacteria Employed for Production of “Advanced Traditional Cheeses”. Foods 2019, 8, 412. https://doi.org/10.3390/foods8090412
Fusco V, Quero GM, Poltronieri P, Morea M, Baruzzi F. Autochthonous and Probiotic Lactic Acid Bacteria Employed for Production of “Advanced Traditional Cheeses”. Foods. 2019; 8(9):412. https://doi.org/10.3390/foods8090412
Chicago/Turabian StyleFusco, Vincenzina, Grazia Marina Quero, Palmiro Poltronieri, Maria Morea, and Federico Baruzzi. 2019. "Autochthonous and Probiotic Lactic Acid Bacteria Employed for Production of “Advanced Traditional Cheeses”" Foods 8, no. 9: 412. https://doi.org/10.3390/foods8090412
APA StyleFusco, V., Quero, G. M., Poltronieri, P., Morea, M., & Baruzzi, F. (2019). Autochthonous and Probiotic Lactic Acid Bacteria Employed for Production of “Advanced Traditional Cheeses”. Foods, 8(9), 412. https://doi.org/10.3390/foods8090412