Protection of Wine from Protein Haze Using Schizosaccharomyces japonicus Polysaccharides

 , ,
, ,  , , and
, , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Wine
2.3. Fermentation Conditions
2.4. Polysaccharides Recovery and Purification
2.5. Wine Treatments
2.6. Polysaccharides Characterization
2.6.1. Polysaccharides Quantification
2.6.2. Monosaccharide Composition
2.6.3. Protein Quantification
2.7. Wine Protein Heat Test and Treatment of the Derived Fractions
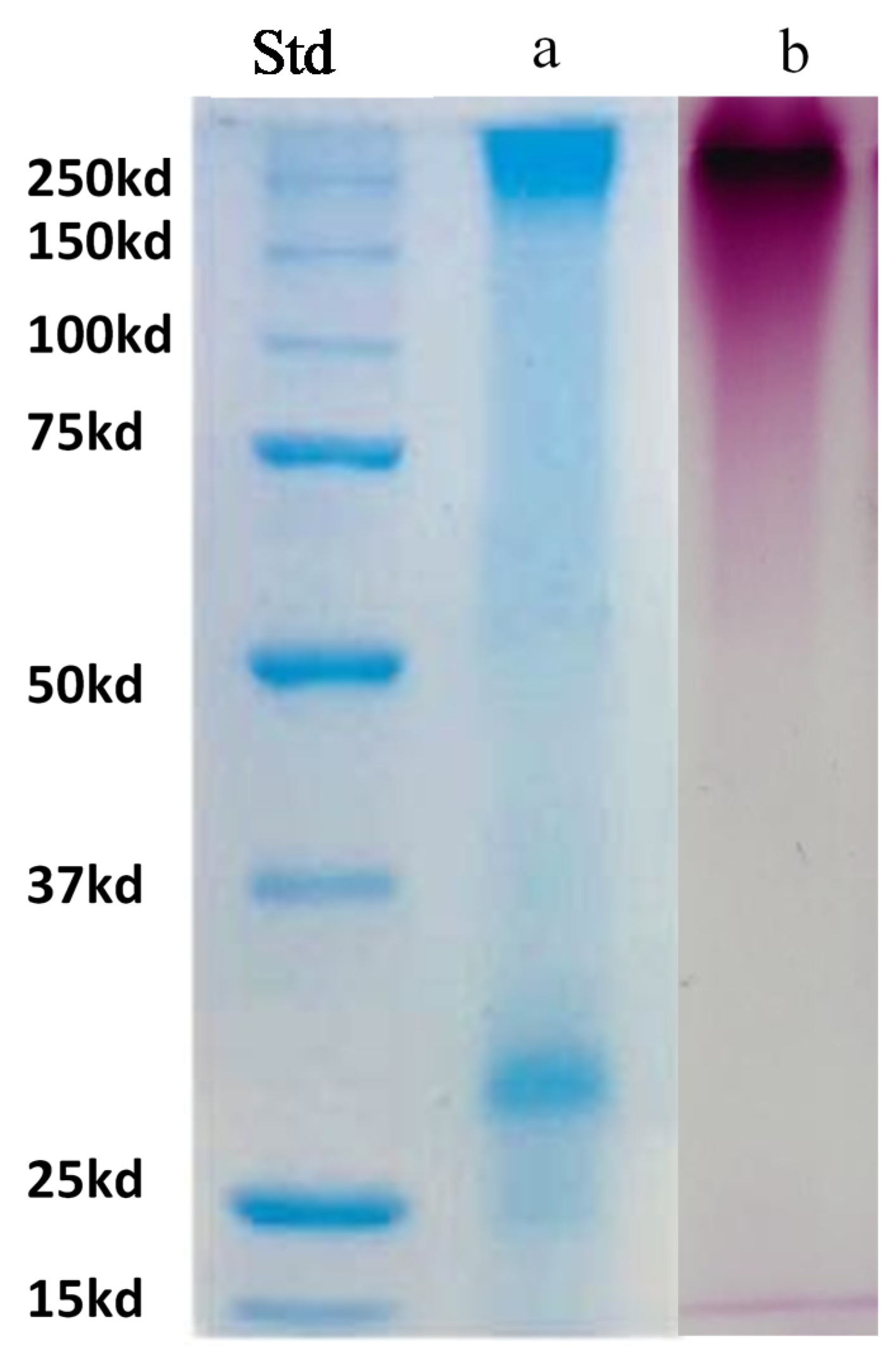
2.8. Proteins Profiling by Gel Electrophoresis
2.9. Dynamic Light Scattering (DLS)
3. Results and Discussion
3.1. Fermentation Performance
3.2. Polysaccharides Quantification and Characterization
3.3. Impact of PSs on Wine Protein Stability
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dordoni, R.; Colangelo, D.; Giribaldi, M.; Giuffrida, M.G.; De Faveri, D.M.; Lambri, M. Effect of Bentonite Characteristics on Wine Proteins, Polyphenols, and Metals under Different pH Conditions. Am. J. Enol. Vitic. 2015, 66, 518–530. [Google Scholar] [CrossRef]
- Making Good Wine. Available online: https://www.panmacmillan.com.au/9781405036016/ (accessed on 1 May 2004).
- Høj, P.B.; Tattersall, D.B.; Adams, K.; Pocock, K.F.; Hayasaka, Y.; van Heeswijck, R.; Waters, E. The ‘haze proteins’ of wine—A summary of properties, factors affecting their accumulation in grapes, and the amount of bentonite required for their removal from wine. In Proceedings of the ASEV 50th Anniversary Meeting, Seattle, WA, USA, 19–23 June 2000. [Google Scholar]
- Lambri, M.; Dordoni, R.; Silva, A.; De Faveri, D.M. Odor-active compound adsorption onto bentonite in a model white wine solution. Chem. Eng. Trans. 2013, 32, 1741–1746. [Google Scholar]
- Francis, I.L.; Sefton, M.A.; Williams, P.J. The sensory effects of pre- or post-fermentation thermal processing on Chardonnay and Semillon wines. Am. J. Enol. Vitic. 1994, 45, 243–251. [Google Scholar]
- Pocock, K.; Høj, P.; Adams, K.; Kwiatkowski, M.; Waters, E. Combined heat and proteolytic enzyme treatment of white wines reduces haze forming protein content without detrimental effect. Aust. J. Grape Wine Res. 2003, 9, 56–63. [Google Scholar] [CrossRef]
- Hsu, J.-C.; Heatherbell, D.A.; Flores, J.H.; Watson, B.T. Heat-unstable proteins in grape juice and wine. II. Characterization and removal by ultrafiltration. Am. J. Enol. Vitic. 1987, 38, 17–22. [Google Scholar]
- Flores, J.H.; Heatherbell, D.A.; McDaniel, M.R. Ultrafiltration of wine: Effect of ultrafiltration on white Riesling and Gewürztraminer wine composition and stability. Am. J. Enol. Vitic. 1990, 41, 207–214. [Google Scholar]
- Marangon, M.; Van Sluyter, S.C.; Robinson, E.M.; Muhlack, R.A.; Holt, H.E.; Haynes, P.A.; Godden, P.W.; Smith, P.; Waters, E. Degradation of white wine haze proteins by Aspergillopepsin I and II during juice flash pasteurization. Food Chem. 2012, 135, 1157–1165. [Google Scholar] [CrossRef]
- Van Sluyter, S.C.; Warnock, N.I.; Schmidt, S.A.; Anderson, P.A.; Van Kan, J.A.L.; Bacic, A.; Waters, E.J. Aspartic Acid Protease from Botrytis cinerea Removes Haze-Forming Proteins during White Winemaking. J. Agric. Food Chem. 2013, 61, 9705–9711. [Google Scholar] [CrossRef]
- Sarmento, M.R.; Boulton, R.B.; Oliveira, J.C. Selection of low swelling materials for protein adsorption from white wines. Int. J. Food Sci. Technol. 2000, 35, 41–47. [Google Scholar] [CrossRef]
- Pashova, V.; Güell, C.; López, F. White Wine Continuous Protein Stabilization by Packed Column. J. Agric. Food Chem. 2004, 52, 1558–1563. [Google Scholar] [CrossRef]
- Salazar, F.N.; Achaerandio, I.; Labbé, M.A.; Güell, C.; López, F. Comparative study of protein stabilization in white wine using zirconia and bentonite: Physiochemical and wine sensory analysis. J. Agric. Food Chem. 2006, 54, 9955–9958. [Google Scholar] [CrossRef] [PubMed]
- Marangon, M.; Lucchetta, M.; Waters, E. Protein stabilization of white wines using zirconium dioxide enclosed in a metallic cage. Aust. J. Grape Wine Res. 2010, 17, 28–35. [Google Scholar] [CrossRef]
- Lucchetta, M.; Pocock, K.F.; Waters, E.; Marangon, M. Use of Zirconium Dioxide during Fermentation as an Alternative to Protein Fining with Bentonite for White Wines. Am. J. Enol. Vitic. 2013, 64, 400–404. [Google Scholar] [CrossRef]
- Mercurio, M.; Mercurio, V.; Gennaro, B.; Gennaro, M.; Grifra, C.; Langella, A.; Morra, V. Natural zeolites and white wines from Campania region (Southern Italy): A new contribution for solving some oenological problems. Period Miner. 2010, 79, 95–112. [Google Scholar]
- Mierczynska-Vasilev, A.; Wahono, S.K.; Smith, P.A.; Bindon, K.; Vasilev, K. Using Zeolites To Protein Stabilize White Wines. ACS Sustain. Chem. Eng. 2019, 7, 12240–12247. [Google Scholar] [CrossRef]
- Vincenzi, S.; Mosconi, S.; Zoccatelli, G.; Pellegrina, C.D.; Veneri, G.; Chignola, R.; Peruo, A.; Curioni, A.; Rizzi, C. Development of a new procedure for protein recovery and quantification in wine. Am. J. Enol. Vitic. 2005, 56, 182–187. [Google Scholar]
- Colangelo, D.; Torchio, F.; De Faveri, D.M.; Lambri, M. The use of chitosan as alternative to bentonite for wine fining: Effects on heat-stability, proteins, organic acids, colour, and volatile compounds in an aromatic white wine. Food Chem. 2018, 264, 301–309. [Google Scholar] [CrossRef]
- Vincenzi, S.; Polesani, M.; Curioni, A. Removal of specific protein components by chitin enhances protein stability in a white wine. Am. J. Enol. Vitic. 2005, 56, 246–254. [Google Scholar]
- Spagna, G.; Pifferi, P.G.; Rangoni, C.; Mattivi, F.; Nicolini, G.; Palmonari, R. The stabilization of white wines by adsorption of phenolic compounds on chitin and chitosan. Food Res. Int. 1996, 29, 241–248. [Google Scholar] [CrossRef]
- Ratnayake, S.; Stockdale, V.; Grafton, S.; Munro, P.; Robinson, A.L.; Pearson, W.; McRae, J.M.; Bacic, A. Carrageenans as heat stabilizers of white wine. Aust. J. Grape Wine Res. 2019, 25, 439–450. [Google Scholar] [CrossRef]
- Marangon, M.; Stockdale, V.J.; Munro, P.; Trethewey, T.; Schulkin, A.; Holt, H.E.; Smith, P. Addition of Carrageenan at Different Stages of Winemaking for White Wine Protein Stabilization. J. Agric. Food Chem. 2013, 61, 6516–6524. [Google Scholar] [CrossRef] [PubMed]
- Marangon, M.; Lucchetta, M.; Duan, D.; Stockdale, V.; Hart, A.; Rogers, P.; Waters, E. Protein removal from a Chardonnay juice by addition of carrageenan and pectin. Aust. J. Grape Wine Res. 2012, 18, 194–202. [Google Scholar] [CrossRef]
- LeDoux, V.; Dulau, L.; Dubourdieu, D. Interprétation de l’amélioration de la stabilité protéique des vins au cours de l’élevage sur lies. J. Int. Sci. Vigne Vin. 1992, 26, 239. [Google Scholar] [CrossRef]
- Waters, E.J.; Wallace, W.; Tate, M.E.; Williams, P.J. Isolation and partial characterization of a natural haze protective factor from wine. J. Agric. Food Chem. 1993, 41, 724–730. [Google Scholar] [CrossRef]
- Waters, E.J.; Pellerin, P.; Brillouet, J.-M. A Saccharomyces mannoprotein that protects wine from protein haze. Carbohydr. Polym. 1994, 23, 185–191. [Google Scholar] [CrossRef]
- Moine-Ledoux, V.; Dubourdieu, D. An invertase fragment responsible for improving the protein stability of dry white wines. J. Sci. Food Agric. 1999, 79, 537–543. [Google Scholar] [CrossRef]
- Dupin, I.V.; McKinnon, B.M.; Ryan, C.; Boulay, M.; Markides, A.J.; Jones, G.P.; Williams, P.J.; Waters, E. Saccharomyces cerevisiae mannoproteins that protect wine from protein haze: Their release during fermentation and lees contact and a proposal for their mechanism of action. J. Agric. Food Chem. 2000, 48, 3098–3105. [Google Scholar] [CrossRef]
- Brown, S.L.; Stockdale, V.J.; Pettolino, F.; Pocock, K.F.; Lopes, M.D.B.; Williams, P.J.; Bacic, A.; Fincher, G.B.; Høj, P.B.; Waters, E. Reducing haziness in white wine by overexpression of Saccharomyces cerevisiae genes YOL155c and YDR055w. Appl. Microbiol. Biotechnol. 2007, 73, 1363–1376. [Google Scholar] [CrossRef]
- Carvalho, E.; Mateus, N.; Plet, B.; Pianet, I.; Dufourc, E.; De Freitas, V. Influence of Wine Pectic Polysaccharides on the Interactions between Condensed Tannins and Salivary Proteins. J. Agric. Food Chem. 2006, 54, 8936–8944. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, I.; Williams, P.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E. The mouth-feel properties of polysaccharides and anthocyanins in a wine like medium. Food Chem. 2004, 85, 519–525. [Google Scholar] [CrossRef]
- Chalier, P.; Angot, B.; Delteil, D.; Doco, T.; Günata, Z. Interactions between aroma compounds and whole mannoprotein isolated from Saccharomyces cerevisiae strains. Food Chem. 2007, 100, 22–30. [Google Scholar] [CrossRef]
- Guadalupe, Z.; Ayestarán, B. Polysaccharide Profile and Content during the Vinification and Aging of Tempranillo Red Wines. J. Agric. Food Chem. 2007, 55, 10720–10728. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ramos, D.; Cebollero, E.; Gonzalez, R. A Recombinant Saccharomyces cerevisiae Strain Overproducing Mannoproteins Stabilizes Wine against Protein Haze. Appl. Environ. Microbiol. 2008, 74, 5533–5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubbers, S.; Léger, B.; Charpentier, C.; Feuillat, M. Effet colloïdes protecteurs d’extraits de parois de levures sur la stabilité tartrique d’un vin modèle. J. Int. Sci. Vigne Vin 1993, 27, 13–22. [Google Scholar]
- Vidal, S.; Williams, P.; Doco, T.; Moutounet, M.; Pellerin, P. The polysaccharides of red wine: Total fractionation and characterization. Carbohydr. Polym. 2003, 54, 439–447. [Google Scholar] [CrossRef]
- Klis, F.M.; Boorsma, A.; Grootab, P.W. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Llaubères, R.M.; Dubourdieu, D.; Villetaz, J.C. Exocellular polysaccharides from Saccharomyces cerevisiae in wine. J. Sci. Food Agric. 1987, 41, 277–286. [Google Scholar] [CrossRef]
- Charpentier, C.; Feuillat, M. Wine Microbiology and Biotechnology; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 225–242. [Google Scholar]
- Boivin, S.; Feuillat, M.; Alexandre, H.; Charpentier, C. Effect of must turbidity on cell wall porosity and macromolecules excretion of Saccharomyces cerevisiae cultivated on grape juice. Am. J. Enol. Vitic. 1998, 49, 325–332. [Google Scholar]
- Charpentier, C.; Dos Santos, A.; Feuillat, M. Release of macromolecules by Saccharomyces cerevisiae during ageing of French flor sherry wine “Vin jaune”. Int. J. Food Microbiol. 2004, 96, 253–262. [Google Scholar] [CrossRef]
- Waters, E.; Alexander, G.; Muhlack, R.; Pocock, K.; Colby, C.; O’Neill, B.; Høj, P.; Jones, P. Preventing protein haze in bottled white wine. Aust. J. Grape Wine Res. 2005, 11, 215–225. [Google Scholar] [CrossRef]
- Ribeiro, T.; Fernandes, C.; Nunes, F.M.; Filipe-Ribeiro, L.; Cosme, F. Influence of the structural features of commercial mannoproteins in white wine protein stabilization and chemical and sensory properties. Food Chem. 2014, 159, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Young, M.; Davies, M.J.; Bailey, D.; Gradwell, M.J.; Smestad-Paulsen, B.; Wold, J.K.; Barnes, R.M.; Hounsell, E.F. Characterization of oligosaccharides from an antigenic mannan of Saccharomyces cerevisiae. Glycoconj. J. 1998, 15, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Rosi, I.; Gheri, A.; Domizio, P.; Fia, G. Production de macromolecules parietals de Saccharomyces cerevisiae au cours de la fermentation et leur influence sur la fermentation malolactique. Rev. Des. Oenol. 2000, 94, 18–20. [Google Scholar]
- Romani, C.; Domizio, P.; Lencioni, L.; Gobbi, M.; Comitini, F.; Ciani, M.; Mannazzu, I. Polysaccharides and glycerol production by non-Saccharomyces wine yeasts in mixed fermentation. Quad. Vitic. Enol. Univ. Torino 2010, 31, 185–189. [Google Scholar]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Comitini, F.; Gobbi, M.; Lencioni, L.; Mannazzu, I.; Ciani, M. Potential spoilage non-Saccharomyces yeasts in mixed cultures with Saccharomyces cerevisiae. Ann. Microbiol. 2010, 61, 137–144. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Liu, Y.; Bisson, L.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2016, 61, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Domizio, P.; Lencioni, L.; Calamai, L.; Portaro, L.; Bisson, L. Evaluation of the Yeast Schizosaccharomyces japonicus for Use in Wine Production. Am. J. Enol. Vitic. 2018, 69, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Spiropoulos, A.; Tanaka, J.; Flerianos, I.; Bisson, L.F. Characterization of hydrogen sulfide formation in commercial and natural wine isolates of Saccharomyces. Am. J. Enol. Vitic. 2000, 51, 233–248. [Google Scholar]
- Romani, C.; Lencioni, L.; Bartolini, A.B.; Ciani, M.; Mannazzu, I.; Domizio, P. Pilot Scale Fermentations of Sangiovese: An Overview on the Impact of Saccharomyces and Non-Saccharomyces Wine Yeasts. Fermentation 2020, 6, 63. [Google Scholar] [CrossRef]
- Chamizo, S.; Adessi, A.; Mugnai, G.; Simiani, A.; De Philippis, R. Soil Type and Cyanobacteria Species Influence the Macromolecular and Chemical Characteristics of the Polysaccharidic Matrix in Induced Biocrusts. Microb. Ecol. 2018, 78, 482–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- McRae, J.M.; Barricklow, V.; Pocock, K.; Smith, P. Predicting protein haze formation in white wines. Aust. J. Grape Wine Res. 2018, 24, 504–511. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Packer, N.H.; Ball, M.S.; Devine, P.L.; Patton, W.F. The Protein Protocols Handbook Walker. In Detection of Glycoproteins in Gels and Blots; Humana Press: Totowa, NJ, USA, 2002; pp. 761–772. [Google Scholar]
- Ju, R.T.C.; Frank, C.W.; Gast, A.P. CONTIN analysis of colloidal aggregates. Langmuir 1992, 8, 2165–2171. [Google Scholar] [CrossRef]
- Manners, D.J.; Meyer, M.T. The molecular structures of some glucans from the cell walls of Schizosaccharomyces pombe. Carbohydr. Res. 1977, 57, 189–203. [Google Scholar] [CrossRef]
- Hart, C.; Schulenberg, B.; Steinberg, T.H.; Leung, W.-Y.; Patton, W.F. Detection of glycoproteins in polyacrylamide gels and on electroblots using Pro-Q Emerald 488 dye, a fluorescent periodate Schiff-base stain. Electrophoresis 2003, 24, 588–598. [Google Scholar] [CrossRef]
- Pellerin, P.; Waters, E.; Brillouet, J.-M.; Moutounet, M. Effet de polysaccharides sur la formation de trouble protéique dans un vin blanc. J. Int. Sci. Vigne Vin 1994, 28, 213. [Google Scholar] [CrossRef]
- Iland, P.G.; Ewart, A.J.W.; Sitters, J.H.; Markides, A.J.; Bruer, N.G.C. Techniques for Chemical Analysis and Quality Monitoring During Winemaking; Patrick Iland Wine Promotions: Campbelltown, Australia, 2000. [Google Scholar]
- Robinson, S.P.; Davies, C. Molecular biology of grape berry ripening. Aust. J. Grape Wine Res. 2000, 6, 175–188. [Google Scholar] [CrossRef]
- Ferreira, R.B.; A Piçarra-Pereira, M.; Monteiro, S.A.; Loureiro, V.B.; Teixeira, A.R. The wine proteins. Trends Food Sci. Technol. 2001, 12, 230–239. [Google Scholar] [CrossRef]
- Van Sluyter, S.C.; McRae, J.M.; Falconer, R.J.; Smith, P.; Bacic, A.; Waters, E.J.; Marangon, M. Wine Protein Haze: Mechanisms of Formation and Advances in Prevention. J. Agric. Food Chem. 2015, 63, 4020–4030. [Google Scholar] [CrossRef] [PubMed]
- Cosme, F.; Fernandes, C.; Ribeiro, T.; Filipe-Ribeiro, L.; Nunes, F.M. White Wine Protein Instability: Mechanism, Quality Control and Technological Alternatives for Wine Stabilization—An Overview. Beverages 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Dufrechou, M.; Vernhet, A.; Roblin, P.; Sauvage, F.-X.; Poncet-Legrand, C. White Wine Proteins: How Does the pH Affect Their Conformation at Room Temperature? Langmuir 2013, 29, 10475–10482. [Google Scholar] [CrossRef] [PubMed]
- Dufrechou, M.; Doco, T.; Poncet-Legrand, C.; Sauvage, F.-X.; Vernhet, A. Protein/Polysaccharide Interactions and Their Impact on Haze Formation in White Wines. J. Agric. Food Chem. 2015, 63, 10042–10053. [Google Scholar] [CrossRef] [PubMed]
- Vernhet, A.; Pellerin, P.; Prieur, C.; Osmianski, J.; Moutounet, M. Charge properties of some grape and wine polysaccharide and polyphenolic fractions. Am. J. Enol. Vitic. 1996, 47, 25–30. [Google Scholar]
- Hassan, P.A.; Rana, S.; Verma, G. Making Sense of Brownian Motion: Colloid Characterization by Dynamic Light Scattering. Langmuir 2014, 31, 3–12. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Parameters | Values |
|---|---|
| pH | 3.33 ± 0.07 |
| Ethanol % (v/v) | 12.35 ± 0.14 |
| Residual sugars (g/L) | 0.69 ± 0.02 |
| Titratable acidity (as tartaric acid) (g/L) | 5.14 ± 0.07 |
| Volatile acidity (as acetic acid) (g/L) | 0.23 ± 0.01 |
| Δ * NTU | 14.00 ± 0.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Millarini, V.; Ignesti, S.; Cappelli, S.; Ferraro, G.; Adessi, A.; Zanoni, B.; Fratini, E.; Domizio, P. Protection of Wine from Protein Haze Using Schizosaccharomyces japonicus Polysaccharides. Foods 2020, 9, 1407. https://doi.org/10.3390/foods9101407
Millarini V, Ignesti S, Cappelli S, Ferraro G, Adessi A, Zanoni B, Fratini E, Domizio P. Protection of Wine from Protein Haze Using Schizosaccharomyces japonicus Polysaccharides. Foods. 2020; 9(10):1407. https://doi.org/10.3390/foods9101407
Chicago/Turabian StyleMillarini, Valentina, Simone Ignesti, Sara Cappelli, Giovanni Ferraro, Alessandra Adessi, Bruno Zanoni, Emiliano Fratini, and Paola Domizio. 2020. "Protection of Wine from Protein Haze Using Schizosaccharomyces japonicus Polysaccharides" Foods 9, no. 10: 1407. https://doi.org/10.3390/foods9101407
APA StyleMillarini, V., Ignesti, S., Cappelli, S., Ferraro, G., Adessi, A., Zanoni, B., Fratini, E., & Domizio, P. (2020). Protection of Wine from Protein Haze Using Schizosaccharomyces japonicus Polysaccharides. Foods, 9(10), 1407. https://doi.org/10.3390/foods9101407