Functional Food from Endangered Ecosystems: Atriplex portulacoides as a Case Study


Abstract
:1. Introduction
2. Morphology and Systematics of Atriplex portulacoides
3. Use History of A. portulacoides
4. Eco-Physiology of A. portulacoides and the Implications for Its Chemical Composition
Metabolism of Heavy Metals
5. Chemical Biomass Composition and Related Nutritional Value
5.1. Lipids
5.2. Carotenoids
5.3. Phenolic Compounds
6. Productivity and Cultivability
6.1. Exploitation as Forage
6.2. Human Nutrition
7. Safety Specifications and Recommendations
8. Toxicology
9. Conclusions and Perspectives
Funding
Conflicts of Interest
Abbreviations
| BW | body weight |
| DW | dry weight |
| EFSA | European Food Safety Authority |
| FW | fresh weight |
| GSH | glutathione |
| LCAA | long chain aliphatic alcohol |
| PSU | practical salinity unit |
| PUFA | Polyunsaturated fatty acid |
References
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Ventura, Y.; Wuddineh, W.A.; Myrzabayeva, M.; Alikulov, Z.; Khozin-Goldberg, I.; Shpigel, M.; Samocha, T.M.; Sagi, M. Effect of seawater concentration on the productivity and nutritional value of annual Salicornia and perennial Sarcocornia halophytes as leafy vegetable crops. Sci. Hortic. 2011, 128, 189–196. [Google Scholar] [CrossRef]
- Norman, H.C.; Masters, D.G.; Barrett-Lennard, E.G. Halophytes as forages in saline landscapes: Interactions between plant genotype and environment change their feeding value to ruminants. Environ. Exp. Bot. 2013, 92, 96–109. [Google Scholar] [CrossRef]
- Maciel, E.; Leal, M.; Lillebø, A.; Domingues, P.; Domingues, M.; Calado, R. Bioprospecting of marine macrophytes using MS-based lipidomics as a new approach. Mar. Drugs 2016, 14, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreira, L.; Resek, E.; Rodrigues, M.J.; Rocha, M.I.; Pereira, H.; Bandarra, N.; da Silva, M.M.; Varela, J.; Custódio, L. Halophytes: Gourmet food with nutritional health benefits? J. Food Compos. Anal. 2017, 59, 35–42. [Google Scholar] [CrossRef]
- Pacicco, L.; Bodesmo, M.; Torricelli, R.; Negri, V. A methodological approach to identify agro-biodiversity hotspots for priority in situ conservation of plant genetic resources. PLoS ONE 2018, 13, e0197709. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodivers. Conserv. 2019, 28, 1343–1360. [Google Scholar] [CrossRef]
- French, K.E. Species composition determines forage quality and medicinal value of high diversity grasslands in lowland England. Agric. Ecosyst. Environ. 2017, 241, 193–204. [Google Scholar] [CrossRef]
- Chapman, V.J. Halimione portulacoides (L.). Aell. J. Ecol. 1950, 38, 214–222. [Google Scholar] [CrossRef]
- Anoè, N.; Calzavara, D.; Salviato, L.; Zanaboni, A. Flora e vegetazione delle barene. In Gli Ambienti Salmastri della Laguna di Venezia; Soc Veneziana di Scienze Naturali: Venezia, Italy, 2001; Volume 26, pp. 9–84. [Google Scholar]
- Kadereit, G.; Borsch, T.; Weising, K.; Freitag, H. Phylogeny of Amaranthaceae and Chenopodiaceae and the evolution of C4 photosynthesis. Int. J. Plant Sci. 2003, 164, 959–986. [Google Scholar] [CrossRef]
- APG (Angiosperm Phylogeny Group) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [CrossRef] [Green Version]
- Moquin-Tandon, A. Chenopodearum Monographica Enumeratio; Loss: Paris, France, 1840; 182p. [Google Scholar]
- Aellen, P. Revision der australischen und neuseelandischen Chenopodiaceae. Bot. Jahrb. Syst. 1938, 68, 345–435. [Google Scholar]
- Acta Plantarum, from 2007+ Etimologia dei Nomi Botanici e Micologici. Available online: http://www.actaplantarum.org/etimologia/etimologia.php (accessed on 6 March 2020).
- Kühn, U.; Bittrich, V.; Carolin, R.; Freitag, H.; Hedge, I.C.; Uotila, P.; Wilson, P.G. Chenopodiaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Rohwer, J.G., Bittrich, V., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; Volume 2, pp. 253–281. [Google Scholar]
- Kadereit, G.; Mavrodiev, E.V.; Zacharias, E.H.; Sukhorukov, A.P. Molecular phylogeny of Atripliceae (Chenopodioideae, Chenopodiaceae): Implications for systematics, biogeography, flower and fruit evolution, and the origin of C4 photosynthesis. Am. J. Bot. 2010, 97, 1664–1687. [Google Scholar] [CrossRef] [PubMed]
- Bair, A.; Howe, M.; Roth, D.; Taylor, R.; Ayers, T.; Kiger, R.W. Vascular Plants of Arizona: Portulacaceae. Canotia 2006, 2, 1–22. [Google Scholar]
- Boestfleisch, C.; Papenbrock, J. Changes in secondary metabolites in the halophytic putative crop species Crithmum maritimum L., Triglochin maritima L. and Halimione portulacoides (L.) Aellen as reaction to mild salinity. PLoS ONE 2017, 12, e0176303. [Google Scholar] [CrossRef] [Green Version]
- Kubiak-Martens, L.; Brinkkemper, O.; Oudemans, T.F. What’s for dinner? Processed food in the coastal area of the northern Netherlands in the Late Neolithic. Veg. Hist. Archaeobot. 2015, 24, 47–62. [Google Scholar] [CrossRef]
- Custódio, M.; Villasante, S.; Cremades, J.; Calado, R.; Lillebø, A.I. Unravelling the potential of halophytes for marine integrated multi-trophic aquaculture (IMTA) a perspective on performance, opportunities and challenges. Aquac. Environ. Interact. 2017, 9, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Guarrera, P.M.; Savo, V. Wild food plants used in traditional vegetable mixtures in Italy. J. Ethnopharmacol. 2016, 185, 202–234. [Google Scholar] [CrossRef]
- Mangia, N.P.; Murgia, M.A.; Fancello, F.; Mouannes, E.; Deiana, P. Tecnologia e controllo microbiologico della mreca, alimento tradizionale a base di Mugil cephalus. Ind. Aliment. 2014, 547, 21–24. [Google Scholar]
- Stienstra, A.W. Mineral composition of Halimione portulacoides (L.) Aellen leaves. Hydrobiologia 1977, 52, 89–91. [Google Scholar] [CrossRef]
- Jensen, A. On the ecophysiology of Halimione portulacoides. Vegetatio 1985, 61, 231–240. [Google Scholar] [CrossRef]
- Crain, C.M.; Silliman, B.R.; Bertness, S.L.; Bertness, M.D. Physical and Biotic Drivers of Plant Distribution across Estuarine Salinity Gradients. Ecology 2004, 85, 2539–2549. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Davy, A.J.; Fernández-Muñoz, F.; Castellanos, E.M.; Luque, T.; Figueroa, M.E. Growth and photosynthetic responses to salinity of the salt-marsh shrub Atriplex portulacoides. Ann. Bot. 2007, 100, 555–563. [Google Scholar]
- Bessieres, M.A.; Gibon, Y.; Lefeuvre, J.C.; Larher, F. A single-step purification for glycine betaine determination in plant extracts by isocratic HPLC. J. Agric. Food Chem. 1999, 47, 3718–3722. [Google Scholar] [CrossRef] [PubMed]
- Storey, R.; Ahma, N.; Jones, R.W. Taxonomic and ecological aspects of the distribution of glycinebetaine and related compounds in plants. Oecologia 1977, 27, 319–332. [Google Scholar] [CrossRef]
- Rabbani, G.; Choi, I. Roles of osmolytes in protein folding and aggregation in cells and their biotechnological applications. Int. J. Biol. Macromol. 2018, 109, 483–491. [Google Scholar] [CrossRef]
- Benzarti, M.; Rejeb, K.B.; Messedi, D.; Mna, A.B.; Hessini, K.; Ksontini, M.; Abdelly, C.; Debez, A. Effect of high salinity on Atriplex portulacoides: Growth, leaf water relations and solute accumulation in relation with osmotic adjustment. S. Afr. J. Bot. 2014, 95, 70–77. [Google Scholar] [CrossRef]
- Cott, G.M.; Reidy, D.T.; Chapman, D.V.; Jansen, M.A. Waterlogging affects the distribution of the saltmarsh plant Atriplex portulacoides (L.) Aellen. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 336–342. [Google Scholar] [CrossRef]
- Reboredo, F. Zinc compartmentation in Halimione portulacoides (L.) Aellen and some effects on leaf ultrastructure. Environ. Sci. Pollut. Res. 2012, 19, 2644–2657. [Google Scholar] [CrossRef]
- Duarte, B.; Carreiras, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Matos, A.R.; Marques, S.C.; Caçador, I. Halophyte fatty acids as biomarkers of anthropogenic-driven contamination in Mediterranean marshes: Sentinel species survey and development of an integrated biomarker response (IBR) index. Ecol. Indic. 2018, 87, 86–96. [Google Scholar] [CrossRef]
- Sousa, A.I.; Caçador, I.; Lillebø, A.I.; Pardal, M.A. Heavy metal accumulation in Halimione portulacoides: Intra-and extra-cellular metal binding sites. Chemosphere 2008, 70, 850–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, R.; Pereira, S.; Lima, A.; Corticeiro, S.; Válega, M.; Pereira, E.; Duarte, A.; Figueira, E. Accumulation, distribution and cellular partitioning of mercury in several halophytes of a contaminated salt marsh. Chemosphere 2009, 76, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Válega, M.; Lima, A.I.G.; Figueira, E.M.A.P.; Pereira, E.; Pardal, M.A.; Duarte, A.C. Mercury intracellular partitioning and chelation in a salt marsh plant, Halimione portulacoides (L.) Aellen: Strategies underlying tolerance in environmental exposure. Chemosphere 2009, 74, 530–536. [Google Scholar]
- Válega, M.; Lillebø, A.I.; Pereira, M.E.; Caçador, I.; Duarte, A.C.; Pardal, M.A. Mercury in salt marshes ecosystems: Halimione portulacoides as biomonitor. Chemosphere 2008, 73, 1224–1229. [Google Scholar] [CrossRef] [Green Version]
- Negrin, V.L.; Teixeira, B.; Godinho, R.M.; Mendes, R.; Vale, C. Phytochelatins and monothiols in salt marsh plants and their relation with metal tolerance. Mar. Pollut. Bull. 2017, 121, 78–84. [Google Scholar] [CrossRef]
- Cartaxana, P.; Catarino, F. Allocation of nitrogen and carbon in an estuarine salt marsh in Portugal. J. Coast. Conserv. 1997, 3, 27–34. [Google Scholar] [CrossRef]
- Harrouni, M.C.; Daoud, S.; Koyro, H.W. Effect of Seawater Irrigation on Biomass Production and Ion Composition of Seven Halophytic Species in Morocco. In Cash Crop Halophytes: Recent Studies; Kratochwil, A., Lieth, H., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 59–70. [Google Scholar]
- Neves, J.P.; Ferreira, L.F.; Simões, M.P.; Gazarini, L.C. Primary production and nutrient content in two salt marsh species, Atriplex portulacoides L. and Limoniastrum monopetalum L., in Southern Portugal. Estuaries Coasts 2007, 30, 459–468. [Google Scholar] [CrossRef]
- Reboredo, F. Some observations on the effects of iron on the leaf ultrastructure of Halimione portulacoides. J. Plant Physiol. 1997, 151, 581–589. [Google Scholar] [CrossRef]
- Vilela, C.; Santos, S.A.; Coelho, D.; Silva, A.M.; Freire, C.S.; Neto, C.P.; Silvestre, A.J. Screening of lipophilic and phenolic extractives from different morphological parts of Halimione portulacoides. Ind. Crop. Prod. 2014, 52, 373–379. [Google Scholar] [CrossRef]
- Boughalleb, N.; Trabelsi, L.; Harzallah-Skhiri, F. Antifungal activity from polar and non-polar extracts of some Chenopodiaceae wild species growing in Tunisia. Nat. Prod. Res. 2009, 23, 988–997. [Google Scholar] [CrossRef]
- Maciel, E.; Lillebø, A.; Domingues, P.; Da Costa, E.; Calado, R.; Domingues, M.R.M. Polar lipidome profiling of Salicornia ramosissima and Halimione portulacoides and the relevance of lipidomics for the valorization of halophytes. Phytochemistry 2018, 153, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Dinan, L.; Whiting, P.; Scott, A.J. Taxonomic distribution of phytoecdysteroids in seeds of members of the Chenopodiaceae. Biochem. Syst. Ecol. 1998, 26, 553–576. [Google Scholar] [CrossRef]
- Parr, M.K.; Botrè, F.; Naß, A.; Hengevoss, J.; Diel, P.; Wolber, G. Ecdysteroids: A novel class of anabolic agents? Biol. Sport 2015, 32, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Ben Nejma, A.B.; Nguir, A.; Jannet, H.B.; Daïch, A.; Othman, M.; Lawson, A.M. New septanoside and 20-hydroxyecdysone septanoside derivative from Atriplex portulacoides roots with preliminary biological activities. Bioorg. Med. Chem. Lett. 2015, 25, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Zakar, T.; Laczko-Dobos, H.; Toth, T.N.; Gombos, Z. Carotenoids assist in cyanobacterial photosystem II assembly and function. Front. Plant Sci. 2016, 7, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Sun, Z.; Sun, P.; Chen, T.; Chen, F. Microalgal carotenoids: Beneficial effects and potential in human health. Food Funct. 2014, 5, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Israr, M.; Duarte, A.C.; Pereira, M.E.; Ahmad, I. Halimione portulacoides (L.) physiological/biochemical characterization for its adaptive responses to environmental mercury exposure. Environ. Res. 2014, 131, 39–49. [Google Scholar] [CrossRef]
- Duarte, B.; Santos, D.; Marques, J.C.; Caçador, I. Ecophysiological adaptations of two halophytes to salt stress: Photosynthesis, PS II photochemistry and anti-oxidant feedback–implications for resilience in climate change. Plant Physiol. Biochem. 2013, 67, 178–188. [Google Scholar] [CrossRef]
- Duarte, B.; Silva, H.; Dias, J.M.; Sleimi, N.; Marques, J.C.; Caçador, I. Functional and ecophysiological traits of Halimione portulacoides and Sarcocornia perennis ecotypes in Mediterranean salt marshes under different tidal exposures. Ecol. Res. 2018, 33, 1145–1156. [Google Scholar] [CrossRef]
- Duarte, B.; Santos, D.; Marques, J.C.; Caçador, I. Impact of heat and cold events on the energetic metabolism of the C3 halophyte Halimione portulacoides. Estuarine Coast. Shelf Sci. 2015, 167, 166–177. [Google Scholar] [CrossRef]
- Benzarti, M.; Rejeb, K.B.; Debez, A.; Messedi, D.; Abdelly, C. Photosynthetic activity and leaf antioxidative responses of Atriplex portulacoides subjected to extreme salinity. Acta Physiol. Plant. 2012, 34, 1679–1688. [Google Scholar] [CrossRef]
- Zengin, G.; Aumeeruddy-Elalfi, Z.; Mollica, A.; Yilmaz, M.A.; Mahomoodally, M.F. In vitro and in silico perspectives on biological and phytochemical profile of three halophyte species—A source of innovative phytopharmaceuticals from nature. Phytomedicine 2018, 38, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Seker, H.; Yolcu, H.; Acikgoz, E. Primary growth parameters of three alfalfa cultivars adapted to highland climatic conditions. J. Agron. Crop Sci. 2015, 201, 219–227. [Google Scholar] [CrossRef]
- Jensen, A. Seasonal changes in near infrared reflectance ratio and sanding crop biomass in a salt marsh community dominated by Halimione portulacoides (L.) AELLEN. New Phytol. 1980, 86, 57–67. [Google Scholar] [CrossRef]
- Benito, I.; Onaindia, M. Biomass and aboveground production of four angiosperms in Cantabrian (N. Spain) salt marshes. Vegetatio 1991, 96, 165–175. [Google Scholar] [CrossRef]
- Groenendijk, A.M. Primary production of four dominant salt-marsh angiosperms in the SW Netherlands. Vegetatio 1984, 57, 143–152. [Google Scholar] [CrossRef]
- Sardo, V. Halophytes and Salt-Tolerant Glycophytes a Potential Resource. In The Use of non-Conventional Water Resources; CIHEAM/EU DG Research: Bari, Italy, 2005; pp. 87–98. [Google Scholar]
- Le Houérou, H.N. Forage Halophytes and Salt-Tolerant Fodder Crops in the Mediterranean Basin. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Squires, V.R., Ayoub, A.T., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 123–137. [Google Scholar]
- Ventura, Y.; Sagi, M. Halophyte crop cultivation: The case for Salicornia and Sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar] [CrossRef]
- Bakker, J.P.; Bos, D.; De Vries, Y. To Graze or not to Graze: That is the Question. In Proceedings of the 10th International Scientific Wadden Sea Symposium, Groningen, The Netherlands, 31 October–3 November 2000; Wolff, W.J., Essink, K., Kellermann, A., Van Leeuwe, M.A., Eds.; University of Groningen: Groningen, The Netherlands, 2003. [Google Scholar]
- Laffaille, P.; Lefeuvre, J.C.; Feunteun, E. Impact of sheep grazing on juvenile sea bass, Dicentrarchus labrax L., in tidal salt marshes. Biol. Conserv. 2000, 96, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Anaya, A.; Herrero, J.; Rosell, C.; Couto, S.; García-Serrano, A. Food habits of wild boars (Sus scrofa) in a Mediterranean coastal wetland. Wetlands 2008, 28, 197–203. [Google Scholar] [CrossRef]
- Bos, D.; Loonen, M.J.; Stock, M.; Hofeditz, F.; Van der Graaf, A.J.; Bakker, J.P. Utilisation of Wadden Sea salt marshes by geese in relation to livestock grazing. J. Nat. Conserv. 2005, 13, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kuijper, D.P.J.; Bakker, J.P. Top-down control of small herbivores on salt-marsh vegetation along a productivity gradient. Ecology 2005, 86, 914–923. [Google Scholar] [CrossRef] [Green Version]
- Attia-Ismail, S.A. Halophytes as Forages. In New Perspectives in Forage Crops; Loiola Edvan, R., Rocha Bezerra, L., Eds.; IntechOpen: London, UK, 2018; pp. 69–87. [Google Scholar] [CrossRef] [Green Version]
- Gihad, E.A.; Shaer, H.E. Utilization of Halophytes by Livestock on Rangelands. Problems and Prospects. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Squires, V.R., Ayoub, A.T., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 77–96. [Google Scholar]
- Miyamoto, S.; Glenn, E.P.; Singh, N.T. Utilization of Halophytic Plants for Fodder Production with Brackish Water in Subtropic Deserts. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Squires, V.R., Ayoub, A.T., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 43–75. [Google Scholar]
- Pacchioli, M.T.; Fattori, G. Gli Alimenti per la Vacca da Latte: I Foraggi; Centro Ricerche Produzioni Animali—C.R.P.A.: Reggio Emilia, Italy, 2014; 81p. [Google Scholar]
- Bortolazzo, E.; Dal Prà, A.; Davolio, R.; Ruozzi, F. Produttività e composizione dell’erba medica. In L’informatore Agrario; Edizioni L’Informatore Agrario Srl: Verona, Italy, 2016; pp. 40–43. [Google Scholar]
- Hojilla-Evangelista, M.P.; Selling, G.W.; Hatfield, R.; Digman, M. Extraction, composition, and functional properties of dried alfalfa (Medicago sativa L.) leaf protein. J. Sci. Food Agric. 2017, 97, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Swingle, R.S.; Glenn, E.P.; Riley, J.J. Halophytes in Mixed Feeds for Livestock. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Squires, V.R., Ayoub, A.T., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 97–100. [Google Scholar]
- CSIRO. Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Collingwood, Australia, 2007; 270p. [Google Scholar]
- Cambrollé, J.; Mancilla-Leytón, J.M.; Muñoz-Vallés, S.; Luque, T.; Figueroa, M.E. Tolerance and accumulation of copper in the salt-marsh shrub Halimione portulacoides. Mar. Pollut. Bull. 2012, 64, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.S. Variability in mineral and trace element content of dairy cattle feeds. J. Dairy Sci. 1975, 58, 1538–1548. [Google Scholar] [CrossRef]
- Kincaid, R.L.; Cronrath, J.D. Amounts and distribution of minerals in Washington forages. J. Dairy Sci. 1983, 66, 821–824. [Google Scholar] [CrossRef]
- Massaro, M.; De Caterina, R. Vasculoprotective effects of oleic acid: Epidemiological background and direct vascular antiatherogenic properties. Nutr. Metab. Cardiovasc. Dis. 2002, 12, 42–51. [Google Scholar]
- Fleming, J.A.; Kris-Etherton, P.M. The evidence for α-linolenic acid and cardiovascular disease benefits: Comparisons with eicosapentaenoic acid and docosahexaenoic acid. Adv. Nutr. 2014, 5, 863S–876S. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Liu, Y.Y.; Wang, X.; Yang, N.; Zhu, H.B.; Zuo, P.P. Protective effects of octacosanol on 6-hydroxydopamine-induced Parkinsonism in rats via regulation of ProNGF and NGF signaling. Acta Pharmacol. Sin. 2010, 31, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Park, S.; Han, D.S.; Park, T. Octacosanol supplementation increases running endurance time and improves biochemical parameters after exhaustion in trained rats. J. Med. Food 2003, 6, 345–351. [Google Scholar] [CrossRef]
- Xu, Z.; Fitz, E.; Riediger, N.; Moghadasian, M.H. Dietary octacosanol reduces plasma triacylglycerol levels but not atherogenesis in apolipoprotein E–knockout mice. Nutr. Res. 2007, 27, 212–217. [Google Scholar] [CrossRef]
- McBride, P.T.; Clark, L.; Krueger, G. Evaluation of triacontanol-containing compounds as anti-inflammatory agents using guinea pig models. J. Investig. Dermatol. 1987, 89, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Sugano, M.; Morioka, H.; Ikeda, I. A comparison of hypocholesterolemic activity of β-sitosterol and β-sitostanol in rats. J. Nutr. 1977, 107, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S.; Manayi, A.; Gohari, A.R.; Abdollahi, M. The story of Beta-sitosterol—A review. Eur. J. Med. Plants 2014, 4, 590–609. [Google Scholar] [CrossRef]
- Ikeda, I.; Morioka, H.; Sugano, M. The effect of dietary β-sitosterol and β-sitostanol on the metabolism of cholesterol in rats. Agric. Biol. Chem. 1979, 43, 1927–1933. [Google Scholar] [CrossRef]
- Samarakoon, S.R.; Fernando, N.; Ediriweera, M.K.; Adhikari, A.; Wijayabandara, L.; de Silva, E.D.; Tennekoon, K.H. Isolation of hopenone-I from the leaves of mangrove plant Scyphiphora hydrophyllacea and its cytotoxic properties. Br. J. Pharm. Res. 2016, 10, 1–6. [Google Scholar] [CrossRef]
- Perera, L.M.S.; Veloz, A.I.R.; GoisRuiz, A.L.T.; Foglio, M.A.; de Oliveira Sousa, I.M.; Tinti, S.V.; Possenti, A.; de Carvalho, J.E. In vitro and In vivo Antiproliferative activity of extracts and fractions of leaves and stem from Tabebuia hypoleuca (C. Wright). Urb. Drug Discov. 2019, 13, 10–21. [Google Scholar]
- Merhan, O. Biochemistry and Antioxidant Properties of Carotenoids. In Carotenoids; Cvetkovic, D., Nikolic, G., Eds.; IntechOpen: London, UK, 2017; pp. 51–66. [Google Scholar]
- Nakajima, Y.; Shimazawa, M.; Otsubo, K.; Ishibashi, T.; Hara, H. Zeaxanthin, a retinal carotenoid, protects retinal cells against oxidative stress. Curr. Eye Res. 2009, 34, 311–318. [Google Scholar] [CrossRef]
- Saud, S.M.; Young, M.R.; Jones-Hall, Y.L.; Ileva, L.; Evbuomwan, M.O.; Wise, J.; Colburn, N.H.; Kim, Y.S.; Bobe, G. Chemopreventive activity of plant flavonoid isorhamnetin in colorectal cancer is mediated by oncogenic Src and β-catenin. Cancer Res. 2013, 73, 5473–5484. [Google Scholar] [CrossRef] [Green Version]
- Chirumbolo, S. Anti-inflammatory action of isorhamnetin. Inflammation 2014, 37, 1200–1201. [Google Scholar] [CrossRef]
- Chi, G.; Zhong, W.; Liu, Y.; Lu, G.; Lü, H.; Wang, D.; Sun, F. Isorhamnetin protects mice from lipopolysaccharide-induced acute lung injury via the inhibition of inflammatory responses. Inflamm. Res. 2016, 65, 33–41. [Google Scholar] [CrossRef]
- Bruno, A.; Rossi, C.; Marcolongo, G.; Di Lena, A.; Venzo, A.; Berrie, C.P.; Corda, D. Selective in vivo anti-inflammatory action of the galactolipid monogalactosyldiacylglycerol. Eur. J. Pharmacol. 2005, 524, 159–168. [Google Scholar] [CrossRef]
- Banskota, A.H.; Gallant, P.; Stefanova, R.; Melanson, R.; O’Leary, S.J. Monogalactosyldiacylglycerols, potent nitric oxide inhibitors from the marine microalga Tetraselmis chui. Nat. Prod. Res. 2013, 27, 1084–1090. [Google Scholar] [CrossRef]
- Valencia, E.; Hardy, G. Practicalities of glutathione supplementation in nutritional support. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, D.; Grosini, M.; Giardina, S.; Michelotti, A.; Carrabetta, M.; Seneci, A.; Verri, M.; Dossena, M.; Marzatico, F. Bioavailability study of an innovative orobuccal formulation of glutathione. Oxidative Med. Cell. Longev. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.; Gangadhar, K.; Vizetto-Duarte, C.; Wubshet, S.; Nyberg, N.; Barreira, L.; Varela, J.; Custódio, L. Maritime halophyte species from southern Portugal as sources of bioactive molecules. Mar. Drugs 2014, 12, 2228–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanella, L.; Pertile, P. Extracts of Halimione Portulacoides and Their Application. U.S. Patent WO2015/040010A1, 26 March 2015. [Google Scholar]
- EFSA-NDA panel. Safety of betaine as a novel food pursuant to Regulation (EC) No 258/97. Scientific opinion adopted on 17 October 2017. EFSA J. 2017, 15, 5057. [Google Scholar] [CrossRef]
- Srivastava, G.K.; Alonso-Alonso, M.L.; Fernandez-Bueno, I.; Garcia-Gutierrez, M.T.; Rull, F.; Medina, J.; Coco, R.M.; Pastor, J.C. Comparison between direct contact and extract exposure methods for PFO cytotoxicity evaluation. Sci. Rep. 2018, 8, 1495. [Google Scholar] [CrossRef]
- Cattani, M.; Mantovani, R.; Schiavon, S.; Bittante, G.; Bailoni, L. Recovery of n-3 polyunsaturated fatty acids and conjugated linoleic acids in ripened cheese obtained from milk of cows fed different levels of extruded flaxseed. J. Dairy Sci. 2014, 97, 123–135. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
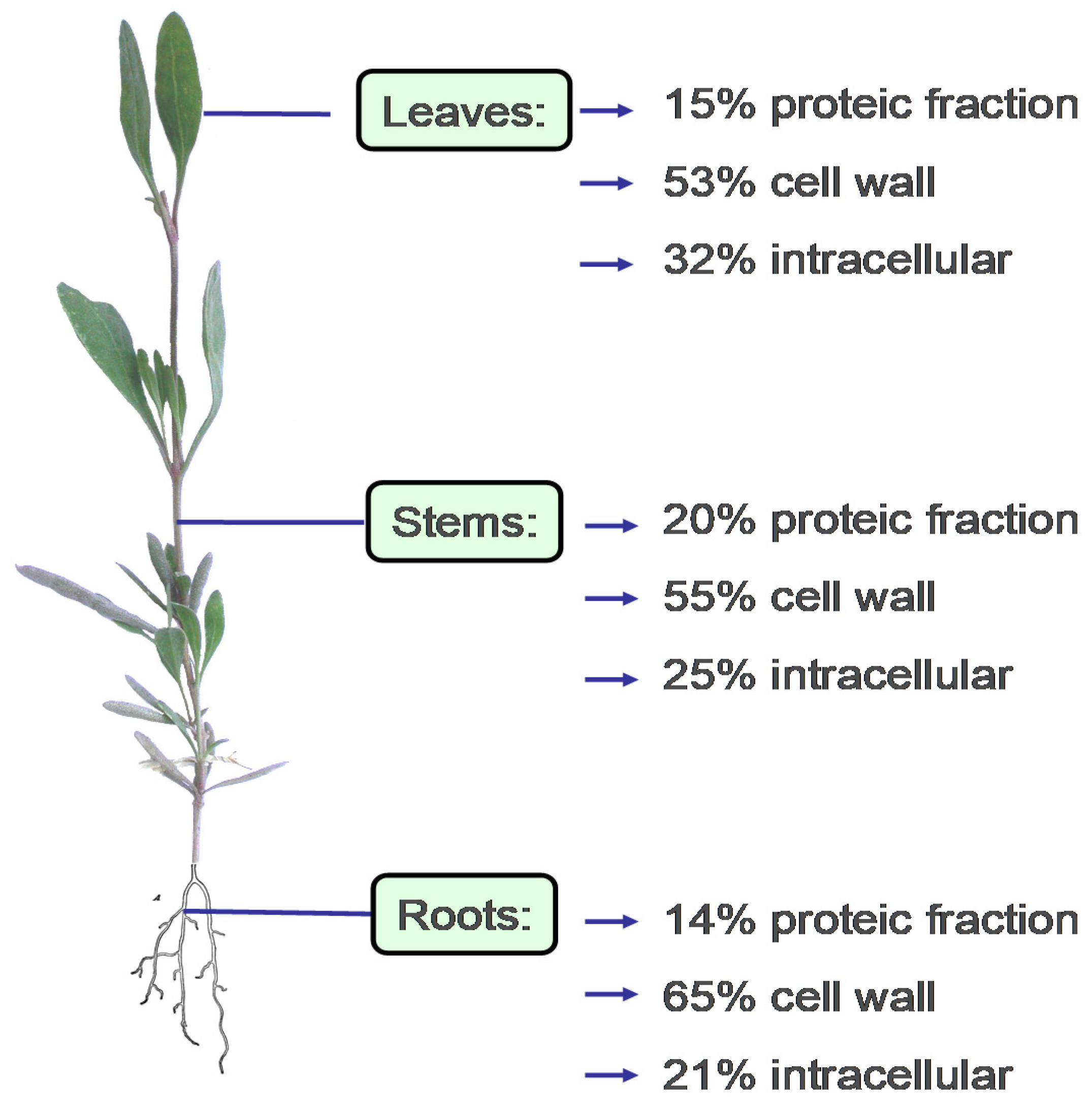{kind=link}
| Leaves | Stems | Shoots (Leaves + Stems) | Roots | Reference | |
|---|---|---|---|---|---|
| Biomass (g/m2) | 212.8 | 1345.6 | 7686.1 | [40] | |
| Nitrogen (% DW) | 1.63 | 1.23 | 0.75 | [40] | |
| Carbon (% DW) | 30.7 | 40.0 | 15.0 | [40] | |
| Moisture (% FW) | 87.5–90.9 * | [31] | |||
| 85.9–86.8 * | 61.5–80.3 * | [41] | |||
| 85.9 | [29] | ||||
| Protein (% DW) | 5.6–9.4 ** | 3.1–4.4 ** | 6.0–7.5 ** | [42] | |
| 10.2% ** | 7.7 ** | 4.7 ** | [40] | ||
| 10–11.2 | 6.6 | 3.5 | [33,43] | ||
| Lipid (% DW) | 2.1 | 1.7 | 3.2 | [44] | |
| Ash (% DW) | 32–40 * | 12–15 * | 12–24 * | [27] | |
| 30–35 * | 10–12 * | [41] | |||
| 27–49 | 9–42 | [25] |
| Lipidic Compounds (mg/kg of Dry Material) | Leaves | Stems | Roots |
|---|---|---|---|
| Fatty acids | 798 | 252 | 374 |
| Saturated | 619 | 193 | 316 |
| Hexadecanoic acid | 99 | 81 | 75 |
| Hexacosanoic acid | 66 | 13 | 18 |
| Octacosanoic acid (montanoic ac.) | 230 | 32 | 70 |
| Triacontanoic acid (melissic ac.) | 147 | 20 | 90 |
| Other saturated fatty acids | 77 | 47 | 63 |
| Unsaturated | 179 | 59 | 58 |
| Octadec-9-enoic acid (oleic ac.) | 115 | 32 | 38 |
| Octadeca-9,12-dienoic acid (linoleic ac.) | 64 | 27 | 20 |
| Long chain aliphatic alcohols | 546 | 82 | 274 |
| 1-Hexacosanol | 132 | 18 | 53 |
| 1-Octacosanol | 252 | 33 | 98 |
| 1-Triacontanol | 122 | 8 | 31 |
| Other long chain aliphatic alcohols | 40 | 23 | 92 |
| Long chain n-alkanes | 106 | 9 | 15 |
| Sterols/triterpenoids | 323 | 609 | 3245 |
| Cholesterol | 66 | 5 | 19 |
| Campesterol | 37 | 4 | 9 |
| Stigmasterol | 20 | 169 | n.d. |
| β-Sitosterol + β-Sitostanol | 148 | n.d. | 291 |
| Spinasterol + β-Sitosterol + β-Sitostanol | n.d. | 122 | n.d. |
| Schottenol | 52 | 30 | 108 |
| Hop-17(21)-en-3-one | n.d. | 169 | 2102 |
| Hop-17(21)-en-3-one (enolic form) | n.d. | 110 | 716 |
| Other identified and unidentified compounds | 273 | 130 | 243 |
| Total detected compounds | 2066 | 1084 | 4151 |
| Phenolic Content (mg/kg of Dry Material) | Leaves | Stems | Roots |
| Isorhamnetin-glucosyl-rhamnosyl-glucuronide isomer | 457 | – | – |
| Isorhamnetin-sulphate-pentoside isomer | 67 | – | – |
| Isorhamnetin-sulphate-glucosyl-rhamnoside | 1190 | – | – |
| Diosmetin-sulphate-glucuronide | 422 | 19 | – |
| Isorhamnetin-sulphate-pentoside isomer | 384 | 18 | – |
| Isorhamnetin-glucosyl-rhamnoside isomer | 107 | 38 | – |
| Isorhamnetin-sulphate-glucosyl-rhamnosyl-glucuronide | 782 | – | – |
| Diosmetin-hexoside | 77 | – | – |
| Isorhamnetin-glucosyl-rhamnoside isomer | 514 | – | – |
| Isorhamnetin-pentoside | 89 | – | – |
| Isorhamnetin-glucosyl-rhamnosyl-glucuronide isomer | 185 | – | – |
| Diosmetin-sulphate | 243 | – | – |
| Diosmetin | 61 | – | – |
| Total detected compounds | 4578 | 76 | – |
| Productivity DW (t/ha) | Ash % | Protein % | Lipid % | |
|---|---|---|---|---|
| Atriplex portulacoides | 10 a | 27 c | 10–11.2 e | 2.1 f |
| Medicago sativa hay | 11 b | 10.3 d | 17 d | 2.1 g |
| Forage grasses (Graminaceae) | 9.9 b | 8.8 b | 1.6 b | |
| Polyphitic meadows | 9.7 b | 8.9 b | 1.6 b | |
| Wheat | 7.7 b | 8.9 b | 2.1 b |
| Sheep | Cattle | Atriplex portulacoides | Corn-Silage | Polyphitic Meadows | Alfalfa | |||
|---|---|---|---|---|---|---|---|---|
| Mineral | Maintenance Requirement (g/kg DW) | Growing, Pregnant, or Lactating Animals (g/kg DW) | Maintenance Requirement (g/kg DW) | Growing, Pregnant, or Lactating Animals (g/kg DW) | (g/kg DW) | (g/kg DW) | (g/kg DW) | (g/kg DW) |
| Calcium | 1.4 | 7.0 | 2.0 | 11.0 | 7–9 a | 2.7 h | 6.8 f | 15.3 f |
| Phosphorus | 0.9 | 3.0 | 1.0 | 3.8 | 1.1–1.4 a | 2.3 h | 2.6 f | 2.6 f |
| Chlorine | 0.3 | 1.0 | 0.7 | 2.4 | 21–31 d | - | - | - |
| Magnesium | 0.9 | 1.2 | 1.3 | 2.2 | 7–10 a | 1.8 h | 1.7 f | 2.4 f |
| Potassium | 5.0 | 5.0 | 5.0 | 5.0 | 43–37 d | 10.7 h | 23.3 f | 26.2 f |
| Sodium | 0.7 | 1.0 | 0.8 | 1.2 | 34–41 d | 0.05 h | 0.14 h | - |
| Sulfur | 2.0 | 2.0 | 1.5 | 1.5 | 2 e | 1.4 h | 2.0 h | - |
| Cobalt | 0.00008 | 0.00015 | 0.00007 | 0.00015 | 0.008–0.010 c | - | - | - |
| Copper | 0.004 | 0.014 | 0.004 | 0.014 | 0.012–0.015 c | 0.0081 h | 0.0129 h | 0.011 i |
| Iodine | 0.0005 | 0.0005 | 0.0005 | 0.0005 | - | - | - | - |
| Iron | 0.040 | 0.040 | 0.040 | 0.040 | 0.194–0.715 g | 0.200 h | 0.184 h | 0.382 i |
| Manganese | 0.020 | 0.025 | 0.020 | 0.025 | 0–0.02 a | 0.0381 h | 0.0764 h | - |
| Selenium | 0.00005 | 0.00005 | 0.00004 | 0.00004 | - | - | - | - |
| Zinc | 0.009 | 0.020 | 0.009 | 0.020 | 0.045–0.108 b | 0.0308 h | 0.0276 h | 0.029 i |
| Age (Years) | Ca (g) | P (g) | Mg (mg) | Na (g) | K (g) | Cl (g) | Fe (mg) | Zn (mg) | Cu (mg) | Se (μg) | I (mg) | Mn (mg) | Mo (μg) | Cr (μg) | F (mg) | |
| Males | 30–59 | 1 | 0.7 | 0.24 | 1.5 | 3.9 | 2.3 | 10 | 12 | 0.9 | 55 | 0.15 | 2.7 | 65 | 35 | 3.5 |
| Females | 30–59 | 1 | 0.7 | 0.24 | 1.5 | 3.9 | 2.3 | 18/10 | 9 | 0.9 | 55 | 0.15 | 2.3 | 65 | 25 | 3.0 |
| FW (g) | Ca (g) | P (g) | Mg (mg) | Na (g) | K (g) | Cl (g) | Fe (mg) | Zn (mg) | Cu (mg) | Se (μg) | I (mg) | Mn (mg) | Mo (μg) | Cr (μg) | F (mg) | |
| A. portulac. | 100 | 0.1 | 0.016 | 0.11 | 0.47 | 0.5 | 0.33 | 5.7 | 1.0 | 0.17 | - | - | 0.13 | - | - | - |
| Chemical Hazard | ||||||
| Code | Element | Limit (mg/kg FW) | Law Reference | |||
| 3.1.11 | Lead | 0.30 | Commission Regulation (EC) n. 1881/2006 of 19 December 2006 | |||
| 3.2.17 | Cadmium | 0.20 | Commission Regulation (EC) n. 629/2008 of 2 July 2008 | |||
| Biological Hazard | ||||||
| Food Category | Microorganism | Limit (cfu/g) | Analytical Reference Method | Stage Where the Criterion Applies | Law Ref. | |
| 1.2. Ready-to-eat foods able to support the growth of L. monocytogenes, other than those intended for infants and for special medical purposes | Listeria monocytogenes | n = 5 c = 0 absent in 25 g or < 100 cfu/g | EN/ISO 11290-2 | Products placed on the market during their shelf-life | (A) | |
| 1.19. Pre-cut fruit and vegetables (ready-to-eat) | Salmonella | n = 5 c = 0 absent in 25 g | EN/ISO 6579 | Products placed on the market during their shelf-life | (A) | |
| 2.5.1. Pre-cut fruit and vegetables (ready-to-eat) | E. coli | n = 5 c = 2 m = 100 M = 1000 | ISO 16649-1 or 16649-2 | Manufacturing process | (A) | |
| 1.29. Sprouts | Shiga toxin producing E. coli (STEC) | n = 5 c = 0 absent in 25 g | CEN/ISO TS 13136 | Products placed on the market during their shelf-life | (B) | |
| other spices and herbs | Enterobacter | n = 5 c = 1 m = 10 M = 100 | ISO 5552 | Import or production/packaging or wholesale/establishment/retail | (C) | |
| other spices and herbs | Clostridium perfrigens | n = 5 c = 1 m = 100 M = 1000 | ISO 7937 | Import or production/packaging or wholesale/establishment/retail | (C) | |
| other spices and herbs | Bacillus cereus | n = 5 c = 1 m = 1000 M = 10,000 | ISO 7932 | Import or production/packaging or wholesale/establishment/retail | (C) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanella, L.; Vianello, F. Functional Food from Endangered Ecosystems: Atriplex portulacoides as a Case Study. Foods 2020, 9, 1533. https://doi.org/10.3390/foods9111533
Zanella L, Vianello F. Functional Food from Endangered Ecosystems: Atriplex portulacoides as a Case Study. Foods. 2020; 9(11):1533. https://doi.org/10.3390/foods9111533
Chicago/Turabian StyleZanella, Lorenzo, and Fabio Vianello. 2020. "Functional Food from Endangered Ecosystems: Atriplex portulacoides as a Case Study" Foods 9, no. 11: 1533. https://doi.org/10.3390/foods9111533
APA StyleZanella, L., & Vianello, F. (2020). Functional Food from Endangered Ecosystems: Atriplex portulacoides as a Case Study. Foods, 9(11), 1533. https://doi.org/10.3390/foods9111533