Effects of Genotype, Storage Temperature and Time on Quality and Compositional Traits of Cherry Tomato

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Material
2.2. Growth Conditions, Fruit Sampling and Storage
2.3. Carpometric Determinations
2.4. Cherry Tomato Quality Variables
2.5. Fruit Chromatic Coordinates
2.6. Total Polyphenols Content
2.7. Carotenoids Extraction and HPLC Analysis
2.8. Statistical Procedures
3. Results
3.1. Carpometric Traits
3.2. Cherry Tomato Quality Variables
3.3. Chromatic Variables
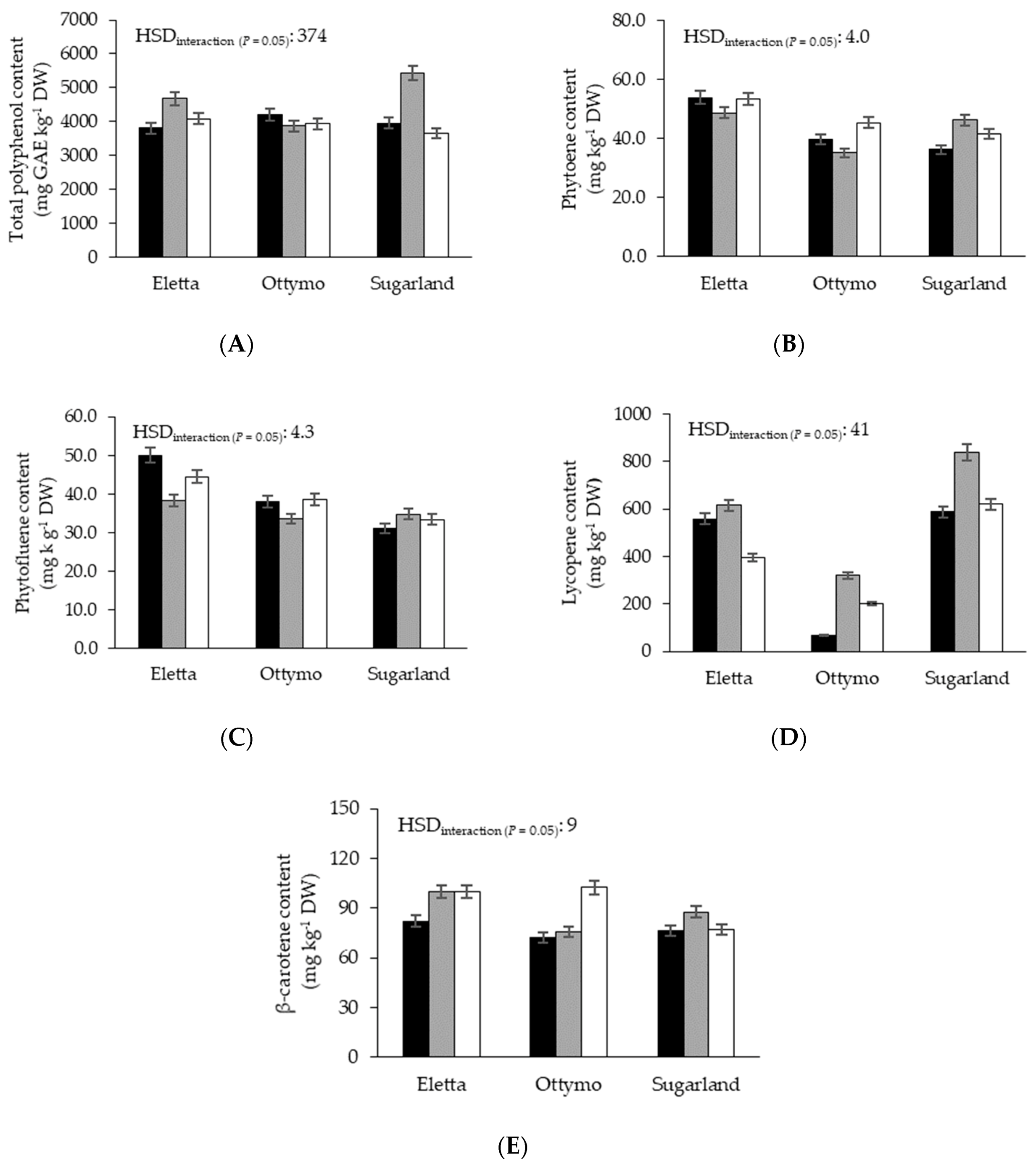
3.4. Total Polyphenols Content
3.5. Carotenoids Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#home (accessed on 26 September 2020).
- Mauro, R.P.; Lo Monaco, A.; Lombardo, S.; Restuccia, A.; Mauromicale, G. Eradication of Orobanche/Phelipanche spp. seedbank by soil solarization and organic supplementation. Sci. Hortic. 2015, 193, 62–68. [Google Scholar] [CrossRef]
- Capurso, C.; Vendemiale, G. The Mediterranean diet reduces the risk and mortality of the prostate cancer: A narrative review. Front. Nutr. 2017, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cainelli, N.; Ruperti, B. Biochemistry and molecular biology in fruits during cold storage. Annu. Plant Rev. Online 2019, 2, 659–688. [Google Scholar]
- Mauro, R.P.; Agnello, M.; Rizzo, V.; Graziani, G.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Recovery of eggplant field waste as a source of phytochemicals. Sci. Hortic. 2020, 261, 109023. [Google Scholar] [CrossRef]
- Petric, T.; Kiferle, C.; Perata, P.; Gonzali, S. Optimizing shelf life conditions for anthocyanin-rich tomatoes. PLoS ONE 2018, 13, e0205650. [Google Scholar] [CrossRef]
- Affandi, F.Y.; Verdonk, J.C.; Ouzounis, T.; Ji, Y.; Woltering, E.J.; Schouten, R.E. Far-red light during cultivation induces postharvest cold tolerance in tomato fruit. Postharvest Biol. Technol. 2020, 159, 111019. [Google Scholar] [CrossRef]
- Tadesse, T.N.; Ibrahim, A.M.; Abtew, W.G. Degradation and formation of fruit color in tomato (Solanum lycopersicum L.) in response to storage temperature. Am. J. Food Technol. 2015, 10, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Farzaneh, V.; Gominho, J.; Pereira, H.; Carvalho, I.S. Screening of the antioxidant and enzyme inhibition ootentials of portuguese Pimpinella anisum L. seeds by GC-MS. Food Anal. Methods 2018, 11, 2645–2656. [Google Scholar] [CrossRef]
- Park, M.H.; Sangwanangkul, P.; Baek, D.R. Changes in carotenoid and chlorophyll content of black tomatoes (Lycopersicon esculentum L.) during storage at various temperatures. Saudi J. Biol. Sci. 2018, 25, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Martí, R.; Roselló, S.; Cebolla-Cornejo, J. Tomato as a source of carotenoids and polyphenols targeted to cancer prevention. Cancers 2016, 8, 58. [Google Scholar] [CrossRef]
- Story, E.N.; Kopec, R.E.; Schwartz, S.J.; Keith Harris, G. An update on the health effects of tomato lycopene. Annu. Rev. Food Sci. Technol. 2010, 1, 189–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, U.; Leitzmann, M.F.; Chatterjee, N.; Wang, Y.; Albanes, D.; Gelmann, E.P.; Friesen, M.D.; Riboli, E.; Hayes, R.B. Serum lycopene, other carotenoids, and prostate cancer risk: A nested case-control study in the prostate, lung, colorectal, and ovarian cancer screening trial. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 962–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Srivastava, A.K. Lycopene; chemistry, biosynthesis, metabolism and degradation under various abiotic parameters. J. Food Sci. Technol. 2015, 52, 41–53. [Google Scholar] [CrossRef]
- Liu, L.; Shao, Z.; Zhang, M.; Wang, Q. Regulation of carotenoid metabolism in tomato. Mol. Plant 2015, 8, 28–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gul, K.; Tak, A.; Singh, A.K.; Singh, P.; Yousuf, B.; Wani, A.A. Chemistry, encapsulation, and health benefits of beta-carotene—A review. Cogent Food Agric. 2015, 1. [Google Scholar] [CrossRef]
- Basu, A.; Imrhan, V. Tomatoes versus lycopene in oxidative stress and carcinogenesis: Conclusions from clinical trials. Eur. J. Clin. Nutr. 2007, 61, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, N.J.; Clinton, S.K.; Erdman, J.W.J. Nutritional aspects of phytoene and phytofluene, carotenoid precursors of lycopene. Adv. Nutr. 2011, 2, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Passam, H.; Karapanos, I.; Bebeli, P.; Savvas, D. A review of recent research on tomato nutrition, breeding and post-harvest technology with reference to fruit quality. Eur. J. Plant Sci. Biotechnol. 2007, 1, 1–21. [Google Scholar]
- Raiola, A.; Rigano, M.M.; Calafiore, R.; Frusciante, L.; Barone, A. Enhancing the health-promoting effects of tomato fruit for biofortified food. Mediators Inflamm. 2014, 2014, 1–16. [Google Scholar] [CrossRef]
- Kavitha, P.; Shivashankara, K.S.; Rao, V.K.; Sadashiva, A.T.; Ravishankar, K.V.; Sathish, G.J. Genotypic variability for antioxidant and quality parameters among tomato cultivars, hybrids, cherry tomatoes and wild species. J. Sci. Food Agric. 2014, 94, 993–999. [Google Scholar] [CrossRef]
- Alenazi, M.M.; Shafiq, M.; Alsadon, A.A.; Alhelal, I.M.; Alhamdan, A.M.; Solieman, T.H.I.; Ibrahim, A.A.; Shady, M.R.; Al-Selwey, W.A. Improved functional and nutritional properties of tomato fruit during cold storage. Saudi J. Biol. Sci. 2020, 27, 1467–1474. [Google Scholar] [CrossRef]
- Mauro, R.P.; Agnello, M.; Distefano, M.; Sabatino, L.; San Bautista Primo, A.; Leonardi, C.; Giuffrida, F. Chlorophyll fluorescence, photosynthesis and growth of tomato plants as affected by long-term oxygen root zone deprivation and grafting. Agronomy 2020, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Giuffrida, F.; Agnello, M.; Mauro, R.P.; Ferrante, A.; Leonardi, C. Cultivation under salt stress conditions influences postharvest quality and glucosinolates content of fresh-cut cauliflower. Sci. Hortic. 2018, 236, 166–174. [Google Scholar] [CrossRef]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How does tomato quality (sugar, acid, and nutritional quality) vary with ripening stage, temperature, and irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Khairi, A.N.; Falah, M.A.F.; Pamungkas, A.P.; Takahashi, N. Optimization of storage temperatures to maintain Lycopene content of tomato from moderate water stress irrigated greenhouse. IOP Conf. Ser. Mater. Sci. Eng. 2018, 403, 011001. [Google Scholar] [CrossRef] [Green Version]
- Carli, P.; Barone, A.; Fogliano, V.; Frusciante, L.; Ercolano, M.R. Dissection of genetic and environmental factors involved in tomato organoleptic quality. BMC Plant Biol. 2011, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- McGuire, R.G. Reporting of objective color measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Atanasova, A.H.; Denev, P.N.; Tringovska, I.I.; Grozeva, S.Y.; Ganeva, D.G.; Kratchanova, M.G.; Panchev, I.N. Optimization of the key parameters for extraction of polyphenol compounds from tomato fruits (Solanum lycopersicum L.). Kinetics of the process. Bulg. Chem. Commun. 2014, 46, 65–70. [Google Scholar]
- Gahler, S.; Otto, K.; Böhm, V. Alterations of vitamin C, total phenolics, and antioxidant capacity as affected by processing tomatoes to different products. J. Agric. Food Chem. 2003, 51, 7962–7968. [Google Scholar] [CrossRef]
- Siracusa, L.; Patanè, C.; Rizzo, V.; Cosentino, S.L.; Ruberto, G. Targeted secondary metabolic and physico-chemical traits analysis to assess genetic variability within a germplasm collection of “long storage” tomatoes. Food Chem. 2018, 244, 275–283. [Google Scholar] [CrossRef]
- Batu, A. Determination of acceptable firmness and colour values of tomatoes. J. Food Eng. 2004, 61, 471–475. [Google Scholar] [CrossRef]
- Bui, H.T.; Makhlouf, J.; Ratti, C. Postharvest ripening characterization of greenhouse tomatoes. Int. J. Food Prop. 2010, 13, 830–846. [Google Scholar] [CrossRef]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Tadesse, T.N.; Abtew, W.G. Effect of hot water treatment on reduction of chilling injury and keeping quality in tomato (Solanum lycopersicum L.) fruits. J. Stored Prod. Postharvest Res. 2016, 7, 61–68. [Google Scholar]
- Leonardi, C.; Baille, A.; Guichard, S. Effects of fruit characteristics and climatic conditions on tomato transpiration in a greenhouse. J. Hortic. Sci. Biotechnol. 1999, 74, 748–756. [Google Scholar] [CrossRef]
- Saladié, M.; Matas, A.J.; Isaacson, T.; Jenks, M.A.; Goodwin, S.M.; Niklas, K.J.; Xiaolin, R.; Labavitch, J.M.; Shackel, K.A.; Fernie, A.R.; et al. A reevaluation of the key factors that influence tomato fruit softening and integrity. Plant Physiol. 2007, 144, 1012–1028. [Google Scholar] [CrossRef] [Green Version]
- Anton, D.; Bender, I.; Kaart, T.; Roasto, M.; Heinonen, M.; Luik, A.; Püssa, T. Changes in polyphenols contents and antioxidant capacities of organically and conventionally cultivated tomato (Solanum lycopersicum L.) fruits during ripening. Int. J. Anal. Chem. 2017, 2017, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Mauro, R.P.; Rizzo, V.; Leonardi, C.; Mazzaglia, A.; Muratore, G.; Distefano, M.; Sabatino, L.; Giuffrida, F. Influence of harvest stage and rootstock genotype on compositional and sensory profile of the elongated tomato cv. Sir Elyan. Agriculture 2020, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Ganje, M.; Jafari, S.M.; Farzaneh, V.; Malekjani, N. Kinetics modelling of color deterioration during thermal processing of tomato paste with the use of response surface methodology. Heat Mass Transf. 2018, 54, 3663–3671. [Google Scholar] [CrossRef]
- Clément, A.; Dorais, M.; Vernon, M. Multivariate approach to the measurement of tomato maturity and gustatory attributes and their rapid assessment by vis-NIR spectroscopy. J. Agric. Food Chem. 2008, 56, 1538–1544. [Google Scholar] [CrossRef]
- Goisser, S.; Wittmann, S.; Fernandes, M.; Mempel, H.; Ulrichs, C. Comparison of colorimeter and different portable food-scanners for non-destructive prediction of lycopene content in tomato fruit. Postharvest Biol. Technol. 2020, 167, 111232. [Google Scholar] [CrossRef]
- Licciardello, F.; Lombardo, S.; Rizzo, V.; Pitino, I.; Pandino, G.; Strano, M.G.; Muratore, G.; Restuccia, C.; Mauromicale, G. Integrated agronomical and technological approach for the quality maintenance of ready-to-fry potato sticks during refrigerated storage. Postharvest Biol. Technol. 2018, 136, 23–30. [Google Scholar] [CrossRef]
- Dattner, M.; Bohn, D. Characterization of print quality in terms of colorimetric aspects. In Printing on Polymers: Fundamentals and Applications; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 329–345. ISBN 9780323374682. [Google Scholar]
- Fernandes, I.; Leça, J.M.; Aguiar, R.; Fernandes, T.; Marques, J.C.; Cordeiro, N. Influence of crop system fruit quality, carotenoids, fatty acids and phenolic compounds in cherry tomatoes. Agric. Res. 2020. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Stinco, C.M.; Meléndez-Martínez, A.J. Comparative study of the bioaccessibility of the colorless carotenoids phytoene and phytofluene in powders and pulps of tomato: Microstructural analysis and effect of addition of sunflower oil. Food Funct. 2018, 9, 5016–5023. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Holst, B.; Williamson, G. Nutrients and phytochemicals: From bioavailability to bioefficacy beyond antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef]
- Mirdehghan, S.H.; Valero, D. Bioactive compounds in tomato fruit and its antioxidant activity as affected by incorporation of Aloe, eugenol, and thymol in fruit package during storage. Int. J. Food Prop. 2017, 20, 1798–1806. [Google Scholar]
- Brashlyanova, B.P.; Pevicharova, G.T. Effects of cold storage and ripening on antioxidant components in tomatoes. Acta Hortic. 2009, 830, 349–353. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Nature and distribution of carotenoids in foods. In Shelf Life Studies of Foods and Beverages Chemical, Biological, Physical and Nutritional Aspects; Charalambous, G., Ed.; Elsevier Science: Amsterdam, The Netherlands, 1993; ISBN 0-444-89459-4. [Google Scholar]
- Gruszecki, W.I.; Strzałka, K. Carotenoids as modulators of lipid membrane physical properties. Biochim. Biophys. Acta Mol. Basis Dis. 2005, 1740, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Dumas, Y.; Dadomo, M.; Di Lucca, G.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidant content of tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Benítez-González, A.; Stinco, C.M. A comprehensive review on the colorless carotenoids phytoene and phytofluene. Arch. Biochem. Biophys. 2015, 572, 188–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapelli-Brahm, P.; Corte-Real, J.; Meléndez-Martínez, A.J.; Bohn, T. Bioaccessibility of phytoene and phytofluene is superior to other carotenoids from selected fruit and vegetable juices. Food Chem. 2017, 229, 304–311. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ‘Eletta’ | ‘Ottymo’ | ‘Sugarland’ | |
|---|---|---|---|
| Seed company | TSI Italia srl, Foggia (FG), Italy | Vilmorin Italia srl, Funo (BO), Italy | Rijk Zwaan Italia srl, Bologna (BO), Italy |
| Fruit color | Deep red | Red | Deep red |
| Average fruit diameter (mm) | 15 ± 2 | 18 ± 2 | 12 ± 1 |
| Average fruit weight (g) | 15.0 ± 1.5 | 20.5 ± 2.5 | 12.0 ± 1.0 |
| Variable | Source of Variation | |||||
|---|---|---|---|---|---|---|
| Storage Temperature | Genotype | Storage Time | (T) × (G) | (T) × (S) | (G) × (S) | |
| Average fruit weight | 13.6 *** | 140.8 *** | 65.6 *** | 3.8 * | NS | 8.1 *** |
| Fruit dry matter | 10.6 ** | 88.4 *** | 29.7 *** | 3.5 * | NS | 8.7 *** |
| Fruit firmness | 8.7 ** | 29.4 *** | 11.5 *** | 3.9 * | NS | NS |
| Reducing sugars content | NS | 50.2 *** | 10.2 *** | NS | NS | 3.8 * |
| SSC/TA | NS | 265.2 *** | 4.7 * | NS | NS | 12.4 *** |
| Fruit pH | NS | 19.6 *** | NS | NS | NS | NS |
| (a*/b*)2 | NS | 38.0 *** | 6.3 ** | NS | NS | NS |
| Chroma | 39.4 *** | 544.7 *** | 30.9 *** | NS | 13.2 *** | NS |
| Tomato color index | NS | 39.9 *** | 4.9 * | NS | NS | NS |
| ΔE*ab | 8.1 ** | 15.1 *** | 5.7 * | 6.0 ** | NS | 5.0 * |
| Total polyphenols content | 20.9 *** | 9.5 *** | 56.4 *** | 17.2 *** | 4.7* | 32.5 *** |
| Phytoene content | NS | 82.8 *** | 7.5 ** | NS | 3.6 * | 15.1 *** |
| Phytofluene content | NS | 44.3 *** | 6.5 ** | 6.3 ** | 3.9 * | 6.8 *** |
| Lycopene content | 33.8 *** | 1462.3 *** | 138.5 *** | 3.6 * | 9.8 *** | 121.4 *** |
| β-carotene content | NS | 17.4 *** | 23.1 *** | NS | NS | 11.7 *** |
| Variable | Genotype | Storage Time | Storage Temperature Mean | |||||
|---|---|---|---|---|---|---|---|---|
| ‘Eletta’ | ‘Ottymo’ | ‘Sugarland’ | S0 | S7 | S14 | |||
| Average fruit weight (g) | T10 | 13.3 | 17.2 | 12.0 | 15.3 | 14.1 | 12.9 | 14.1 a |
| T20 | 13.0 | 15.5 | 11.1 | 15.5 | 13.6 | 12.0 | 13.2 b | |
| Mean | 13.2 b | 16.4 a | 11.6 c | 15.4 a | 13.9 b | 12.5 c | ||
| HSDinteraction | 0.8 | 0.8 | ||||||
| Fruit dry matter content (%) | T10 | 8.2 | 12.1 | 9.1 | 9.1 | 9.4 | 10.9 | 9.8 b |
| T20 | 8.2 | 13.9 | 10.2 | 9.2 | 10.3 | 12.9 | 10.8 a | |
| Mean | 8.2 c | 13.0 a | 9.7 b | 9.2 b | 9.8 b | 11.9 a | ||
| HSDinteraction | 1.1 | NS | ||||||
| Fruit firmness (N) | T10 | 14.1 | 19.7 | 12.8 | 16.5 | 16.1 | 14.1 | 15.6 a |
| T20 | 13.6 | 16.0 | 11.6 | 16.6 | 12.9 | 11.7 | 13.7 b | |
| Mean | 13.8 b | 17.9 a | 12.2 c | 16.6 a | 14.5 b | 12.9 c | ||
| HSDinteraction | 2.2 | NS | ||||||
| Variable | Genotype | Storage Time | Storage Temperature Mean | |||||
|---|---|---|---|---|---|---|---|---|
| ‘Eletta’ | ‘Ottymo’ | ‘Sugarland’ | S0 | S7 | S14 | |||
| Reducing sugars content (g kg−1 DW) | T10 | 434 | 595 | 552 | 557 | 555 | 463 | 527 a |
| T20 | 420 | 597 | 579 | 565 | 536 | 499 | 529 a | |
| Mean | 427 b | 596 a | 565 a | 561 a | 545 a | 481 b | ||
| HSDinteraction | NS | NS | ||||||
| SSC/TA (adimensional) | T10 | 2.15 | 1.65 | 1.52 | 1.80 | 1.77 | 1.71 | 1.77 a |
| T20 | 2.18 | 1.66 | 1.54 | 1.86 | 1.74 | 1.80 | 1.79 a | |
| Mean | 2.16 a | 1.65 b | 1.53 c | 1.83 a | 1.76 b | 1.76 b | ||
| HSDinteraction | NS | NS | ||||||
| Fruit pH | T10 | 4.59 | 4.21 | 4.33 | 4.37 | 4.35 | 4.42 | 4.38 a |
| T20 | 4.61 | 4.22 | 4.38 | 4.38 | 4.42 | 4.42 | 4.40 a | |
| Mean | 4.60 a | 4.21 b | 4.36 ab | 4.37 a | 4.38 a | 4.42 a | ||
| HSDinteraction | NS | NS | ||||||
| Variable | Genotype | Storage Time | Storage Temperature Mean | |||||
|---|---|---|---|---|---|---|---|---|
| ‘Eletta’ | ‘Ottymo’ | ‘Sugarland’ | S0 | S7 | S14 | |||
| (a*/b*)2 | T10 | 0.79 | 0.74 | 0.59 | 0.72 | 0.67 | 0.73 | 0.71 a |
| T20 | 0.84 | 0.71 | 0.63 | 0.72 | 0.67 | 0.78 | 0.73 a | |
| Mean | 0.81 a | 0.73 b | 0.61 c | 0.72 a | 0.67 b | 0.75 a | ||
| HSDinteraction | NS | NS | ||||||
| Chroma | T10 | 26.8 | 26.5 | 21.1 | 24.5 | 24.1 | 26.0 | 24.8 a |
| T20 | 26.1 | 25.2 | 20.3 | 24.3 | 23.1 | 24.1 | 23.8 b | |
| Mean | 26.4 a | 25.9 b | 20.7 c | 24.4 b | 23.6 c | 25.1 a | ||
| HSDinteraction | NS | 0.7 | ||||||
| TCI | T10 | 33.9 | 34.9 | 31.1 | 33.1 | 32.9 | 33.8 | 33.3 a |
| T20 | 34.1 | 34.2 | 31.6 | 33.1 | 32.6 | 34.0 | 33.3 a | |
| Mean | 34.0 a | 34.5 a | 31.3 b | 33.1 ab | 32.7 b | 33.9 a | ||
| HSDinteraction | NS | NS | ||||||
| ΔE*ab | T10 | 1.37 | 2.65 | 3.11 | - | 2.18 | 2.57 | 2.38 a |
| T20 | 1.49 | 2.36 | 1.77 | - | 1.64 | 2.10 | 1.87 b | |
| Mean | 1.43 b | 2.50 a | 2.44 a | - | 1.91 b | 2.34 a | ||
| HSDinteraction | 0.6 | NS | ||||||
| Variable | Genotype | Storage Time | Storage Temperature Mean | |||||
|---|---|---|---|---|---|---|---|---|
| ‘Eletta’ | ‘Ottymo’ | ‘Sugarland’ | S0 | S7 | S14 | |||
| TPC (mg GAE kg−1 FW) | T10 | 4087 | 4531 | 4364 | 3997 | 4869 | 4116 | 4327 a |
| T20 | 4287 | 4167 | 3647 | 3982 | 4452 | 3668 | 4034 b | |
| Mean | 4187 b | 4349 a | 4006 c | 3989 b | 4660 a | 3892 b | ||
| HSDinteraction | 303 | 303 | ||||||
| Phytoene content (mg kg−1 FW) | T10 | 50.4 | 41.5 | 41.2 | 43.3 | 45.5 | 45.1 | 44.3 a |
| T20 | 53.5 | 38.5 | 41.3 | 43.1 | 41.2 | 48.3 | 44.5 a | |
| Mean | 51.9 a | 40.0 b | 41.2 b | 43.2 b | 43.3 b | 46.7 a | ||
| HSDinteraction | NS | 3.2 | ||||||
| Phytofluene content (mg kg−1 FW) | T10 | 42.1 | 38.7 | 32.3 | 39.6 | 37.7 | 35.6 | 37.7 a |
| T20 | 46.5 | 34.6 | 33.9 | 39.8 | 33.4 | 41.9 | 38.3 a | |
| Mean | 44.3 a | 36.6 b | 33.1 c | 39.7 a | 35.5 c | 38.8 b | ||
| HSDinteraction | 3.5 | 3.5 | ||||||
| Lycopene content (mg kg−1 FW) | T10 | 488 | 185 | 662 | 392 | 526 | 404 | 445 b |
| T20 | 556 | 207 | 701 | 416 | 577 | 484 | 488 a | |
| Mean | 552 b | 196 c | 682 a | 404 c | 552 a | 444 b | ||
| HSDinteraction | 35 | 35 | ||||||
| β-carotene content (mg kg−1 DW) | T10 | 92.0 | 85.2 | 83.4 | 77.0 | 89.7 | 94.0 | 86.9 a |
| T20 | 96.0 | 81.7 | 77.1 | 76.7 | 85.9 | 92.3 | 84.9 a | |
| Mean | 94.0 a | 83.5 b | 80.3 b | 76.8 c | 87.8 b | 93.1 a | ||
| HSDinteraction | NS | NS | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Distefano, M.; Arena, E.; Mauro, R.P.; Brighina, S.; Leonardi, C.; Fallico, B.; Giuffrida, F. Effects of Genotype, Storage Temperature and Time on Quality and Compositional Traits of Cherry Tomato. Foods 2020, 9, 1729. https://doi.org/10.3390/foods9121729
Distefano M, Arena E, Mauro RP, Brighina S, Leonardi C, Fallico B, Giuffrida F. Effects of Genotype, Storage Temperature and Time on Quality and Compositional Traits of Cherry Tomato. Foods. 2020; 9(12):1729. https://doi.org/10.3390/foods9121729
Chicago/Turabian StyleDistefano, Miriam, Elena Arena, Rosario Paolo Mauro, Selina Brighina, Cherubino Leonardi, Biagio Fallico, and Francesco Giuffrida. 2020. "Effects of Genotype, Storage Temperature and Time on Quality and Compositional Traits of Cherry Tomato" Foods 9, no. 12: 1729. https://doi.org/10.3390/foods9121729
APA StyleDistefano, M., Arena, E., Mauro, R. P., Brighina, S., Leonardi, C., Fallico, B., & Giuffrida, F. (2020). Effects of Genotype, Storage Temperature and Time on Quality and Compositional Traits of Cherry Tomato. Foods, 9(12), 1729. https://doi.org/10.3390/foods9121729