Antibiofilm and Enzyme Inhibitory Potentials of Two Annonaceous Food Spices, African Pepper (Xylopia aethiopica) and African Nutmeg (Monodora myristica)

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extraction
2.2. GC-FID (Gas Chromatography Flame Ionization Detector) and GC-MS Analyses
2.3. Determination of MIC (Minimal Inhibitory Concentration)
2.4. Assay of Inhibition of Bacterial Biofilm Formation by Extracts
2.5. Violacein Pigment Inhibition Assay
2.6. Swimming and Swarming Motility Inhibition on Pseudomonas Aeruginosa PA01
2.7. Cholinesterase Inhibition Assay
2.8. Tyrosinase Inhibition Assay
2.9. Urease Inhibition Assay
2.10. Statistical Analysis
3. Results
3.1. GC-MS Chemical Composition
3.2. Minimal Inhibitory Concentrations
3.3. Percentage Biofilm Inhibition
3.4. Violacein and Motility (Swimming and Swarming) Inhibition Percentages
3.5. Enzyme Inhibitory Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Corn, C. Scents of Eden: A History of the Spice Trade; Kindasha: New York, NY, USA, 1999. [Google Scholar]
- Chinenye, M.H. Antibacterial Activity of Piper Guineense, Xylopia Aethiopica and Allium Cepa Against Bacteria Isolated from Spoilt Soup Preparations. Ph.D. Thesis, University of Nigeria, Nsukka, Nigeria, 2014; p. 1. [Google Scholar]
- Enabulele, S.A.; Oboh, F.O.J.; Uwadiae, E. Antimicrobial, nutritional and phytochemical properties of Monodora myristica seeds. IOSR J. Pharm. Biol. Sci. 2014, 9, 1–6. [Google Scholar] [CrossRef]
- Okagu, I.U.; Ogugua, V.N.; Dibor, C.N.I.; Maryann, U.; Nnebe, M.E.; Aniehe, C.C.; Odenigbo, C.J.; Ngwu, U.E. Effects of methanol extraction on some nutritional and antinutrient contents of Xylopia aethiopica fruits from Enugu State, Nigeria. Asian J. Agric. Food Sci. 2018, 6, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Tatsadjieu, L.N.; Essia-Ngang, J.J.; Nyassoum, M.B.; Etoa, F.X. Antibacterial and antifungal activity of Xylopia aethiopica, Monodora myristica, Xanthoxylum xanthoxyloides and Xanthoxylum leprieurii from Cameroun. Fitoterapia 2003, 74, 469–472. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Ozgur, C.; Godloves, C.F.; Mehmet, O.; Mehmet, E.D.; Farzana, S. Antibiofilm, antiquorum sensing and antioxidant activity of secondary metabolites from seeds of Annona senegalensis, Persoon. Mic Pathog 2020, 144, 104191. [Google Scholar] [CrossRef]
- Erhirhie, O.; Moke, G.E. Xylopia Aethiopica: A Review of its ethnomedicinal, chemical and pharmacological properties earnest. Am. J. Pharm. Tech. Res. 2014, 4, 22–37. [Google Scholar]
- Choumessi, T.A.; Danel, M.; Chassaing, S.; Truchet, I.; Penlap, V.B.; Pieme, A.C.; Asonganyi, T.; Ducommum, B.; Valette, A. Characterization of the anti-proliferative activity of Xylopia aethiopica. Cell Div. 2012, 7, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Adefegha, S.A.; Oboh, G. Effect of diets supplemented with Ethiopian pepper [Xylopia aethiopica (Dun.) A. Rich (Annonaceae)] and Ashanti pepper [Piper guineense Schumach. et Thonn (Piperaceae)] on some biochemical parameters in normal rats. Asian Pac. J. Trop. Biomed. 2012, 16, S558–S566. [Google Scholar] [CrossRef]
- Soh, D.; Nkwengoua, E.; Ngantchou, I.; Nyasse, B.; Denier, C.; Hannaert, V.; Shaker, K.H.; Schneider, B. Xylopioxyde and other bioactive kaurane-diterpenes from Xylopia aethiopica Dunal (Annonaceae). J. Appl. Pharm. Sci. 2013, 3, 013–019. [Google Scholar]
- Boampong, J.N.; Ameyaw, E.O.; Aboagye, B.; Asare, K.; Kyei, S.; Donfack, J.H.; Woode, E. The curative and prophylactic effects of xylopic acid on Plasmodium berghei infection in mice. J. Parasitol. Res. 2013, 2013, 356107. [Google Scholar]
- Etoundi, C.B.; Kuate, D.; Ngondi, J.L.; Oben, J. Anti-amylase, anti-lipase and antioxidant effects of aqueous extracts of some Cameroonian spices. J. Nat. Prod. 2010, 3, 165–171. [Google Scholar]
- Abd-Alg, N.N.; El-Kamali, H.H.; Ramadan, M.M.; Ghanem, K.Z.; Farrag, A.R.H. Xylopia aethiopica volatile compounds protect against panadol-induced hepatic and renal toxicity in male rats. World Appl. Sci. J. 2013, 27, 10–22. [Google Scholar]
- Mohammed, A.; Islam, S.M. Antioxidant activity of xylopia aethiopica fruits acetone fraction in a type 2 diabetes model of rats. Biomed. Pharmacother. 2017, 96, 30–36. [Google Scholar] [CrossRef]
- Weiss, E.A. Spice Crops; CABI Publishing: Oxon, UK, 2002; pp. 102–103. [Google Scholar]
- Bakarnga-Via, I.; Hzounda, J.B.; Fokou, P.V.; Tchokouaha, L.R.; Gary-Bobo, M.; Gallud, A.; Garcia, M.; Walbadet, L.; Secka, Y.; Dongmo, P.M.; et al. Composition and cytotoxic activity of essential oils from Xylopia aethiopica (Dunal) A. Rich, Xylopia parviflora (A. Rich) Benth.) and Monodora myristica (Gaertn) growing in Chad and Cameroon. BMC Comp. Altern. Med. 2014, 14, 125. [Google Scholar] [CrossRef] [Green Version]
- Nwozo, S.O.; Fowokemi, K.T.; Oyinloye, B.E. African nutmeg (Monodora myristica) lowers cholesterol and modulates lipid peroxidation in experimentally induced hypercholesterolemic male wistar rats. Int. J. Biomed. Sci. 2015, 11, 86–92. [Google Scholar]
- Ogunmoyole, T.; Inaboya, S.; Makun, J.O.; Kade, I.J. Differential antioxidant properties of ethanol and water soluble phytochemicals of false nutmeg (Monodora myristica) seeds. Int. J. Biochem. Biotechnol. 2013, 2, 253–262. [Google Scholar]
- Lekana-Douki, J.B.; Bongui, J.B.; Oyegue, L.S.L.; Zang-Edou, S.E.; Zatra, R.; Bisvigoua, U.; Druilhe, P.; Lebibi, J.; Ndouo, F.S.T.; Kombila, M. In vitro antiplasmodial activity and cytotoxicity of nine plants traditionally used in Gabon. J. Ethnopharmacol. 2011, 133, 1103–1108. [Google Scholar] [CrossRef]
- Lacroix, D.; Prado, S.; Kamoga, D.; Kasenene, J.; Namukobe, J.; Krief, S.; Dumontet, V.; Mouray, E.; Bodo, B.; Brunois, F. Antiplasmodial and cytotoxic activities of medicinal plants traditionally used in the village of Kiohima, Uganda. J. Ethnopharmacol. 2011, 133, 850–855. [Google Scholar] [CrossRef]
- Oyinloye, B.E.; Adenowo, A.F.; Osunsanmi, F.O.; Ogunyinka, B.I.; Nwozo, S.O.; Kappo, A.P. Aqueous extract of Monodora myristica ameliorates cadmium-induced hepatotoxicity in male rats. SpringerPlus 2016, 5, 641. [Google Scholar] [CrossRef] [Green Version]
- Akinwunmi, K.F.; Oyedapo, O.O. In vitro anti-inflammatory evaluation of African nutmeg (Monodora myristica) seeds. Eur. J. Med. Plants. 2015, 8, 167–174. [Google Scholar] [CrossRef]
- Ishola, I.O.; Ikumawoyi, V.O.; AfolayaN, G.O.; Olorife, O.J. Antinociceptive and anti-inflammatory properties of hydroethanolic seed extract of Monodora myristica (Annonaceae) in rodents. West Afr. J. Pharm. 2016, 27, 22–32. [Google Scholar]
- Nwankwo, P.O. Comparative study of the antioxidant activities of Monodora myristica and A. sceptrum on protein and lipid levels of diabetic-induced rats. IOSR J. Biotechnol. Biochem. 2015, 1, 63–71. [Google Scholar]
- Agiriga, A.; Siwela, M. Monodora myristica (Gaertn.) Dunal: A plant with multiple food, health and medicinal applications: A review. Am. J. Food Technol. 2017, 12, 271–284. [Google Scholar] [CrossRef]
- Talla, E.; Tamfu, A.N.; Gade, I.S.; Yanda, L.; Mbafor, J.T.; Sophie, L.; Vander-Elst, L.; Popova, M.; Bankova, V. New mono-ether of glycerol and triterpenes with DPPH radical scavenging activity from Cameroonian propolis. Nat. Prod. Res. 2017, 31, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Tamfu, N.A.; Jabeen, A.; Tagatsing, F.M.; Tariq, A.B.; Farzana, S. Organic and mineral composition of seeds of Afrostyrax lepidophyllus Mildbr. and evaluation of ROS inhibition and cytotoxicity of isolated compounds. Chem. Afr. 2019, 2, 615–624. [Google Scholar]
- CLSI (Clinical Laboratory Standards Institute). Quality Control Minimal Inhibitory Concentration (MIC) Limits for Broth Dilution and MIC Interpretative Breakpoints (M27-s2); Clinical Laboratory of Standards Institute: Wayne, PA, USA, 2006. [Google Scholar]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and Analyzing Static Biofilms. Current Protocols in Microbiol; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005. [Google Scholar]
- Packiavathy, I.A.S.V.; Agilandeswari, P.; Musthafa, K.S.; Pandian, S.K.; Ravi, A.V. Antibiofilm and quorum sensing inhibitory potential of Cuminum cyminum and its secondary metabolite methyl eugenol against Gram negative bacterial pathogens. Food Res. Int. 2012, 45, 85–92. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Khattak, S.; Saeed-Ur-Rehman, S.; Khan, T.; Ahmad, M. In vitro enzyme inhibition activities of crude ethanolic extracts derived from medicinal plants of Pakistan. Nat. Prod. Res. 2005, 19, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Olusimbo, O.A.; Abimbola, R.E.; Chinonye, L.A. Antimicrobial activities of some Nigerian spices on some pathogens. Agric. Biol. J. N. Am. 2011, 2, 1187–1193. [Google Scholar]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vasconcelos, A.T.R.; De Almeida, D.F.; Hungria, M.; Guimarães, C.T.; Antonio, R.V.; Almeida, F.C.; De Almeida, L.G.; De Almeida, R.; Alves-Gomes, J.A.; Andrade, E.M.; et al. The complete genome sequence of Chromobacterium violaceum reveals remarkable and exploitable bacterial adaptability. Proc. Natl. Acad. Sci. USA 2003, 100, 11660–11665. [Google Scholar]
- Kothari, V.; Sharma, S.; Padia, D. Recent research advances on Chromobacterium violaceum. Asian Pac. J. Trop. Med. 2017, 10, 744–752. [Google Scholar] [CrossRef] [PubMed]
- O’May, C.; Tufenkji, N. The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Environ. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K. Cholinesterase inhibitors as Alzheimer’s therapeutics (Review). Mol. Med. Reports. 2019, 20, 1479–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svane, S.; Sigurdarson, J.J.; Finkenwirth, F.; Eitinger, T.; Karring, H. Inhibition of urease activity by different compounds provides insight into the modulation and association of bacterial nickel import and ureolysis. Sci. Rep. 2020, 10, 8503. [Google Scholar] [CrossRef]
- Altameme, H.J.; Hameed, I.H.; Kareem, M.A. Analysis of alkaloid phytochemical compounds in the ethanolic extract of Datura stramonium and evaluation of antimicrobial activity. Afr. J. Biotechnol. 2015, 14, 1668–1674. [Google Scholar]
- Mohammed, A.B.; Hafiz, A.M.; Hassan, A.A.; Sohier, M.S.; Ashraf, N.A.; Husham, E.H.; Shahnaz, S.; Waquar, A.; Asaad, K. Phytochemical, Cytotoxic, and Antimicrobial Evaluation of the Fruits of Miswak Plant, Salvadora persica L. J. Chem. 2020, 2020, 4521951. [Google Scholar]
- Selwyn, S.; Durodie, J. The antimicrobial activity of sugar against pathogens of wounds and other infections of man. In Properties of Water in Foods; Simatos, D., Multon, J.L., Eds.; NATO ASI Series (Series E: Applied Sciences); Springer: Dordrecht, The Netherlands, 1985; Volume 90, pp. 293–308. [Google Scholar]
- Singh, B.R. Antibacterial Activity of Glycerol, Lactose, Maltose, Mannitol, Raffinose and Xylose. Med. Noto-Are 2014, 2014, 17223318. [Google Scholar]
- Bisignano, G.; Sanogo, R.; Marino, A.; Aquinol, R.; D’Angelo, V.; Germano, M.P.; De-Pasquale, R.; Pizza, C. Antimicrobial activity of Mitracarpus scaber extract and isolated constituents. Lett. Appl. Microbiol. 2000, 30, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Katsure, S.V.; Suvarna, A.K.; Dhiraj, M.; Rupali, W.; Mahalaxmi, M.; Sanjay, B.K. Antioxidant and Antiparkinson Activity of Gallic Acid Derivatives. Pharmacologyonline 2009, 1, 385–395. [Google Scholar]
- Vennila, V.; Udayakumar, R. GC-MS Analysis of Leaf, Fruits and Latex of Croton bonplandianum Baill. Int. J. Biochem. Res. Rev. 2015, 5, 187–197. [Google Scholar] [CrossRef]
- Hameed, I.H.; Altameme, H.J.; Idan, S.A. Artemisia annua: Biochemical products analysis of methanolic aerial parts extract and anti-microbial capacity. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 1843–1868. [Google Scholar]
- Wang, J.; Gu, X.-Z.; He, L.-M.; Li, C.-C.; Qiu, W.-W. Synthesis of ursodeoxycholic acid from plant-source (20S)-21-hydroxy-20-methylpregn-4-en-3-one. Steroids 2020, 157, 108600. [Google Scholar] [CrossRef]
- Zhe, F.; Su, Y.J.; Hyun, A.J.; Jae, S.C.; Byung, S.M.; Mi, H.W. Anticholinesterase and Antioxidant Constituents from Gloiopeltis furcate. Chem. Pharm. Bull. 2010, 58, 1236–1239. [Google Scholar]
- Mustapha, N.A.; Runner, R.T.M. GC-MS Analysis and Preliminary Antimicrobial Activity of Albizia adianthifolia (Schumach) and Pterocarpus angolensis (DC). Medicines 2016, 3, 3. [Google Scholar]
- Krishnan, K.; James, R.F.; Mohan, A. Isolation and characterization of n-hexadecanoic acid from Canthium parviflorum leaves. J. Chem. Pharm. Res. 2016, 8, 614–617. [Google Scholar]
- Mahalingam, G.; Krishnan, K. Antidiabetic activity of 2-hydroxy 4-methoxy benzoic acid isolated from the roots of Hemidesmus indicus on streptozotocin-induced diabetic rats. Int. J. Diabetes Metabol. 2009, 17, 53–57. [Google Scholar]
- Shao, C.; Guo, Z.; Peng, H.; Peng, G.; Huang, Z.; She, Z.; Lin, Y. A new isoprenyl phenyl ether compound from mangrove fungus. Chem. Nat. Compd. 2007, 43, 377–380. [Google Scholar] [CrossRef]
- Ramnath, M.G.; Thirugnanasampandan, R.; Sadasivam, M.; Mohan, P.S. Antioxidant, antibacterial and antiacetylcholinesterase activities of abietic acid from Isodon wightii (Bentham) H. Hara. Free Rad. Antiox. 2015, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Elaissi, A.; Rouis, Z.; Salem, N.A.; Mabrouk, S.; Ben-Salem, Y.; Salah, K.B.; Aouni, M.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F.; et al. Chemical composition of 8 eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2012, 12, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavijo-Romero, A.; Quintanilla-Carvajal, M.X.; Ruiz, Y. Stability and antimicrobial activity of eucalyptus essential oil emulsions. Food Sci. Technol. Int. 2018, 25, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Huizhen, S.; Haiming, C.; Xiaolong, W.; Yueying, H.; Yonghuan, Y.; Qiuping, Z.; Weijun, C.; Wenxue, C. Antimicrobial Activity and Proposed Action Mechanism of 3-Carene against Brochothrix thermosphacta and Pseudomonas fluorescens. Molecules 2019, 24, 3246. [Google Scholar]



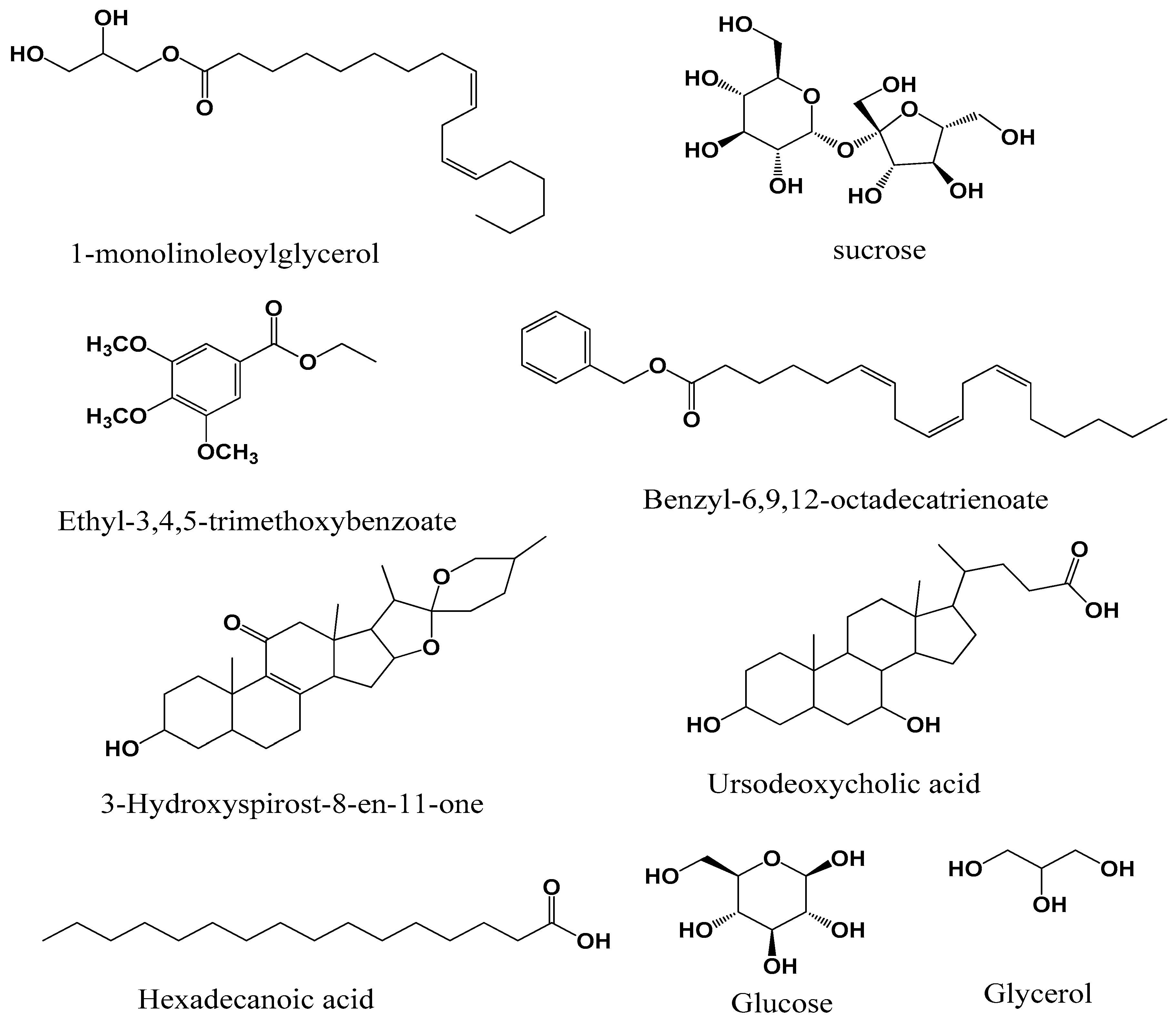{kind=link}
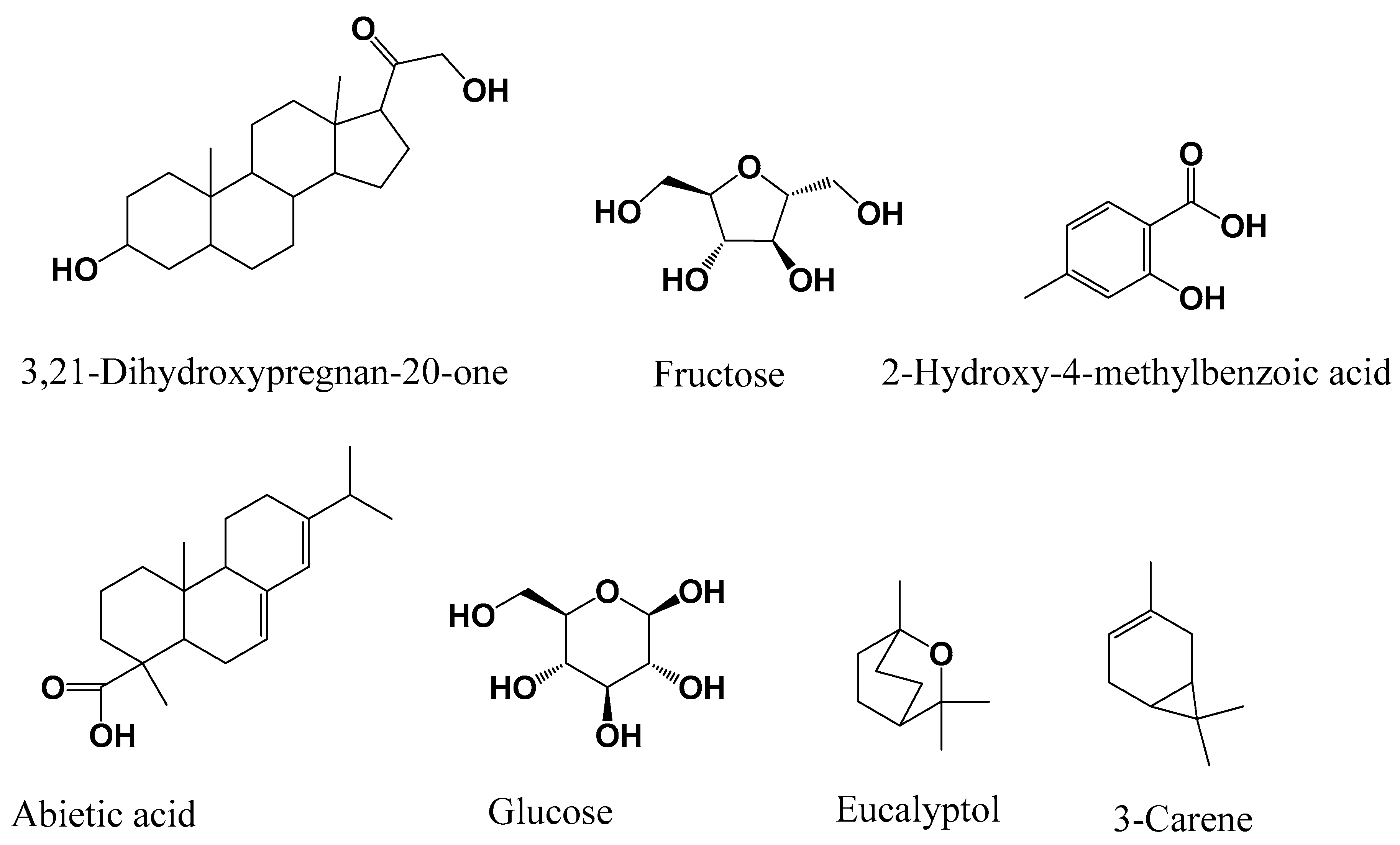{kind=link}
| Microorganisms | Extracts | |
|---|---|---|
| X. aethiopica | M. myristica | |
| C. albicans ATCC 10239 | 0.625 | 0.625 |
| S. aureus ATCC 25923 | 0.625 | 0.3125 |
| E. coli ATCC 25922 | 2.5 | 2.5 |
| C. violaceum CV12472 | 2.5 | 1.25 |
| P. aeruginosa PA01 | 0.625 | 1.25 |
| Strains | Conc. | X. aethiopica | M. myristica |
|---|---|---|---|
| % of Biofilm Inhibition | |||
| S. aureus ATCC 25923 | MIC | 45.3 ± 1.5 * | 52.6 ± 3.3 |
| MIC/2 | 30.0 ± 3.4 | 39.0 ± 1.4 | |
| MIC/4 | 16.7 ± 0.4 | 23.8 ± 0.6 | |
| MIC/8 | 5.2 ± 1.0 | 11.3 ± 1.5 | |
| MIC/16 | - | - | |
| E. coli ATCC 25922 | MIC | 21.0 ± 2.5 | 18.6 ± 0.5 |
| MIC/2 | 10.5 ± 0.5 | 8.3 ± 2.0 | |
| MIC/4 | - | - | |
| MIC/8 | - | - | |
| MIC/16 | - | - | |
| C. albicans ATCC 10239 | MIC | 31.7 ± 1.5 | 29.6 ± 3.3 |
| MIC/2 | 22.4 ± 1.3 | 15.3 ± 0.1 | |
| MIC/4 | 09.2 ± 0.5 | 8.9 ± 2.0 | |
| MIC/8 | - | - | |
| MIC/16 | - | - | |
| X. aethiopica | M. myristica | |||
|---|---|---|---|---|
| Conc. | % of Violacein Inhibition (on C. violaceum CV12472) | |||
| MIC | 80.5 ± 3.0 * | 75.1 ± 2.5 | ||
| MIC/2 | 37.3 ± 0.7 | 35.8 ± 0.6 | ||
| MIC/4 | 19.4 ± 1.0 | 15.5 ± 1.1 | ||
| MIC/8 | 5.6 ± 0.2 | - | ||
| MIC/16 | - | - | ||
| % motility inhibition (on P. aeruginosa PA01) | ||||
| Conc. (μg/mL) | Swimming inh. | Swarming inh. | Swimming inh. | Swarming inh. |
| 100 | 25.1 ± 0.5 | 28.9 ± 2.0 | 29.2 ± 0.0 | 31.7 ± 1.2 |
| 75 | 13.6 ± 0.5 | 12.0 ± 0.5 | 16.5 ± 0.8 | 22.3 ± 1.0 |
| 50 | - | - | - | 11.6 ± 0.5 |
| Sample | AChE | BChE | Tyrosinase | Urease |
|---|---|---|---|---|
| % Inhibition | ||||
| X. aethiopica | 7.80 ± 0.36 | 15.48 ± 0.57 | 30.45 ± 0.84 | 5.69 ± 0.33 |
| M. myristica | 9.65 ± 0.11 | 13.64 ± 0.88 | 23.44 ± 1.27 | 15.18 ± 0.86 |
| Galantamine | 80.76 ± 0.52 | 74.43 ± 0.30 | NT | NT |
| Kojic acid | NT | NT | NT | 67.58 ± 0.23 |
| Thiourea | NT | NT | 75.17 ± 0.18 | NT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamfu, A.N.; Ceylan, O.; Kucukaydin, S.; Ozturk, M.; Duru, M.E.; Dinica, R.M. Antibiofilm and Enzyme Inhibitory Potentials of Two Annonaceous Food Spices, African Pepper (Xylopia aethiopica) and African Nutmeg (Monodora myristica). Foods 2020, 9, 1768. https://doi.org/10.3390/foods9121768
Tamfu AN, Ceylan O, Kucukaydin S, Ozturk M, Duru ME, Dinica RM. Antibiofilm and Enzyme Inhibitory Potentials of Two Annonaceous Food Spices, African Pepper (Xylopia aethiopica) and African Nutmeg (Monodora myristica). Foods. 2020; 9(12):1768. https://doi.org/10.3390/foods9121768
Chicago/Turabian StyleTamfu, Alfred Ngenge, Ozgur Ceylan, Selcuk Kucukaydin, Mehmet Ozturk, Mehmet Emin Duru, and Rodica Mihaela Dinica. 2020. "Antibiofilm and Enzyme Inhibitory Potentials of Two Annonaceous Food Spices, African Pepper (Xylopia aethiopica) and African Nutmeg (Monodora myristica)" Foods 9, no. 12: 1768. https://doi.org/10.3390/foods9121768
APA StyleTamfu, A. N., Ceylan, O., Kucukaydin, S., Ozturk, M., Duru, M. E., & Dinica, R. M. (2020). Antibiofilm and Enzyme Inhibitory Potentials of Two Annonaceous Food Spices, African Pepper (Xylopia aethiopica) and African Nutmeg (Monodora myristica). Foods, 9(12), 1768. https://doi.org/10.3390/foods9121768