Discovery of Unexpected Sphingolipids in Almonds and Pistachios with an Innovative Use of Triple Quadrupole Tandem Mass Spectrometry

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
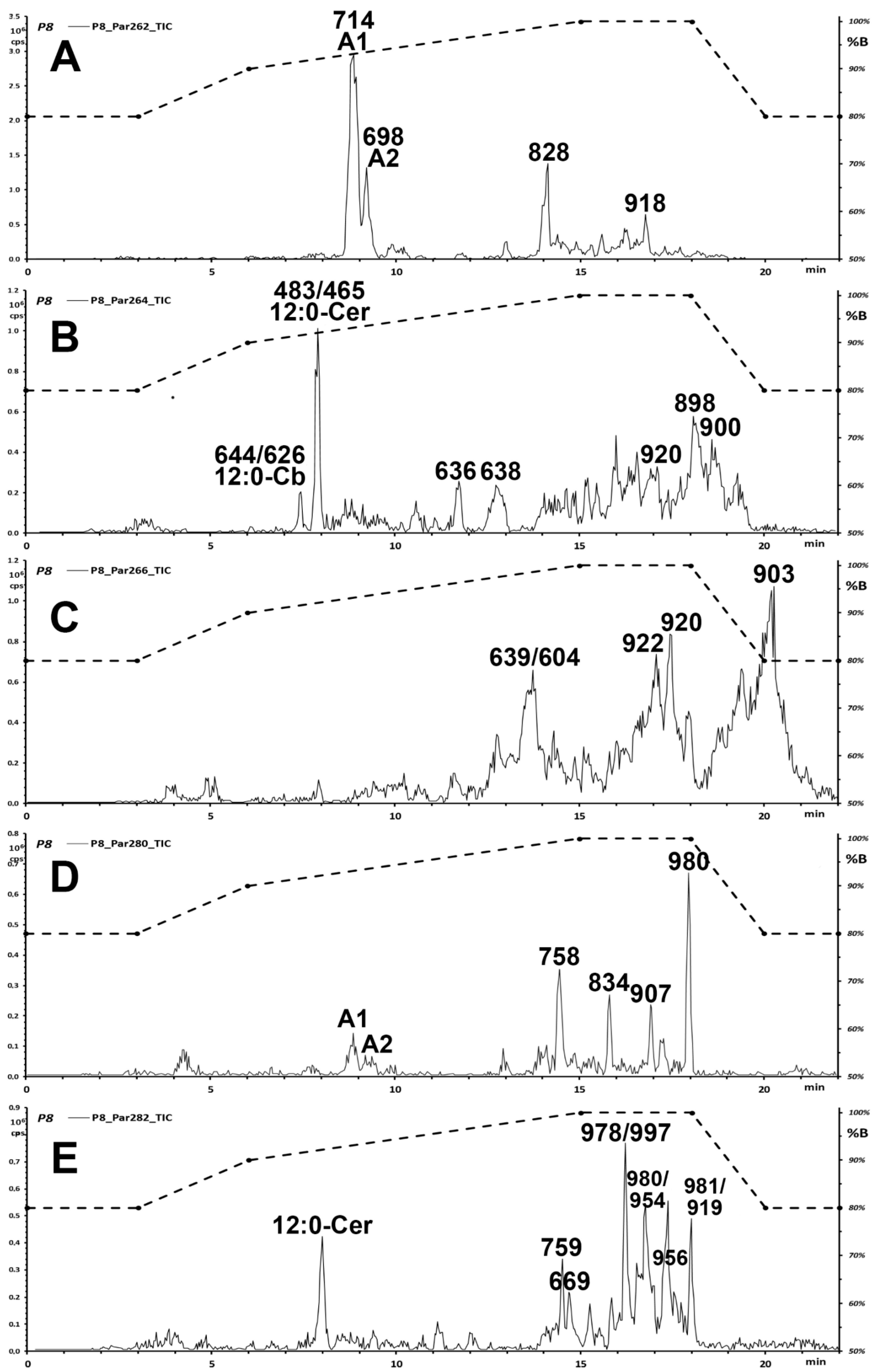
2.1. Systematic Discovery of Sphingolipids in Almonds and Pistachios
- (a)
- The use of full scan Precursor Ion (PI scan) of five O” ceramide reporter fragments (m/z 262, 264, 266, 280, 282; Table 1) to highlight ceramides with modified sphingosines;
- (b)
- The use of a full scan Neutral Loss of C6H12O6 hexose (180.2 Da) to ascertain whether the putatively highlighted ceramides have a hexose attached unit;
- (c)
- The use of a collision energy ramp synchronized to the scan of Q1 in the PI and NL modes (iso-energetic Precursor Ion, i-PI, and iso-energetic Neutral Loss, i-NL) to analyze all ceramides at the same value of effective collision energy (see Appendix A);
- (d)
- An extended isocratic step (in respect to the previously published conditions [9]) at full gradient strength during the UPLC analysis of sphingolipids (total analysis time 22 min) to investigate the possible presence of ceramides with much longer fatty acids.
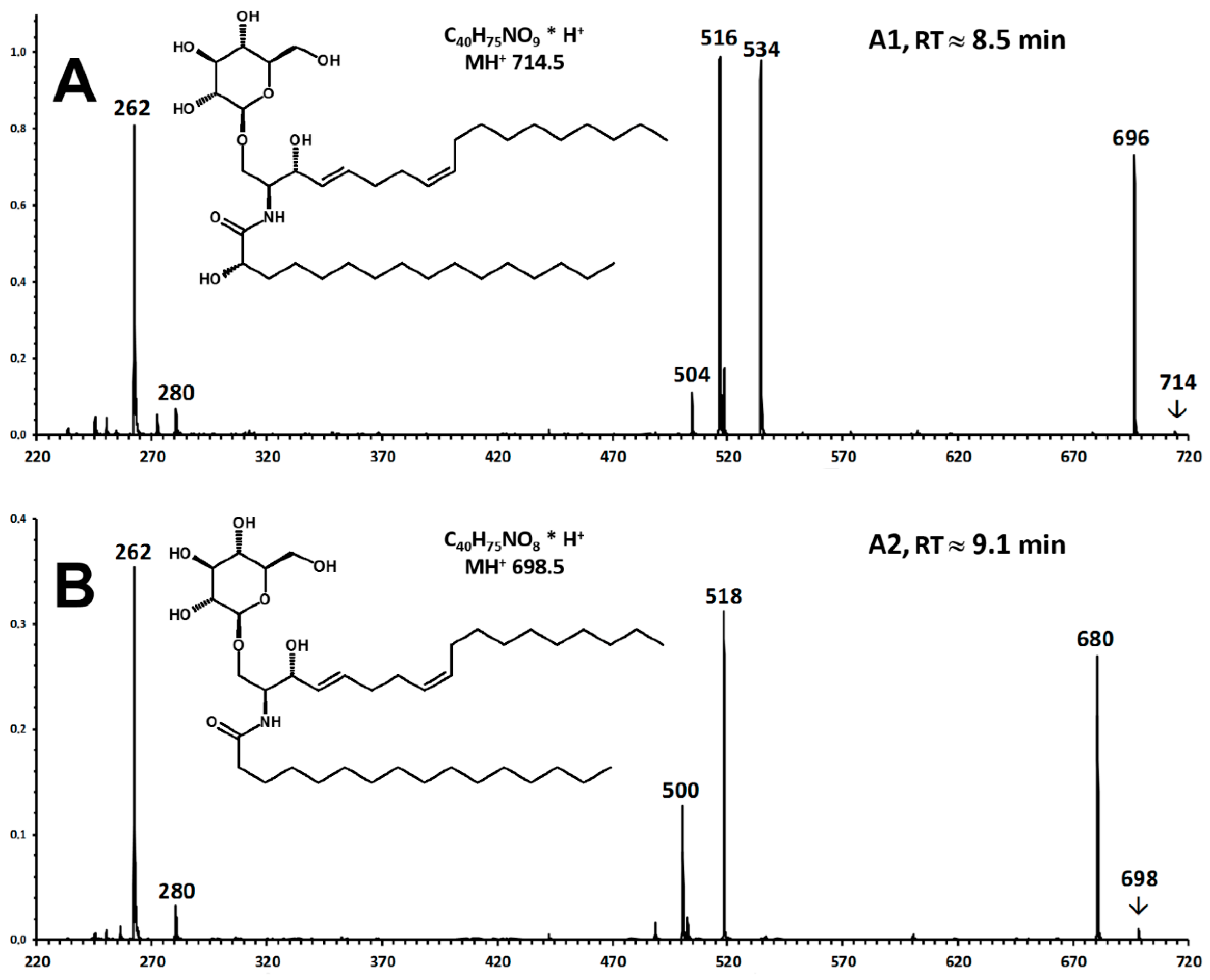
- A modified sphingosine that carries a further unsaturation in the C18 chain (d18:2, fragment ion at m/z 262 Th);
- The C-1 of sphingosine is linked to a hexose (paired losses of 180 Da from MH+ and [MH − H2O]+ ions);
- The 2-amino group of the sphingosine is linked to two saturated C16 fatty acids (mass difference between m/z 262 and the fragments generated by hexose loss), one of which (earlier-eluting compound A1) carries an additional hydroxyl group.
2.2. Levels of the Discovered Sphingolipids in Almonds and Pistachios
2.3. Interference of Triglycerides in the Discovery of Ceramides in Almonds and Pistachios
3. Discussion
4. Materials and Methods
4.1. Reagents, Chemicals, and Standards
4.2. Plant Material
4.3. Sphingolipid Extraction Procedure
4.4. LC-MS/MS Instrumentation
4.5. Separation and Detection of Sphingolipids by LC-MS/MS
4.6. Untargeted Discovery LC-MS/MS Analysis by Iso-Energetic Precursor Ion and Neutral Loss Scan in a Triple Quadrupole
4.7. Relationship of Molecular Structure to Chromatographic Retention
4.8. Isotope Pattern Calculation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
- On the mass of the resting target gas in the collision cell (m TAR) and
- On the mass (m/z) of the impinging (singly-charged) precursor ion (m PAR),
References
- Hawker, J.S.; Buttrose, M.S. Development of the Almond Nut (Prunus dulcis (Mill.) DA Webb). Anatomy and Chemical Composition of Fruit Parts from Anthesis to Maturity. Ann. Bot. 1980, 46, 313–321. [Google Scholar] [CrossRef]
- Summo, C.; Palasciano, M.; De Angelis, D.; Paradiso, V.M.; Caponio, F.; Pasqualone, A. Evaluation of the chemical and nutritional characteristics of almonds (Prunus dulcis (Mill). DA Webb) as influenced by harvest time and cultivar. J. Sci. Food Agric. 2018, 98, 5647–5655. [Google Scholar] [CrossRef] [Green Version]
- Alasalvar, C.; Bolling, B.W. Review of nut phytochemicals, fat-soluble bioactives, antioxidant components and health effects. Br. J. Nutr. 2015, 113, S68–S78. [Google Scholar] [CrossRef]
- Wang, D.D.; Toledo, E.; Hruby, A.; Rosner, B.A.; Willett, W.C.; Sun, Q.; Razquin, C.; Zheng, Y.; Ruiz-Canela, M.; Guasch-Ferré, M.; et al. Plasma ceramides, mediterranean diet, and incident cardiovascular disease in the PREDIMED trial (prevención con dieta mediterránea). Circulation 2017, 135, 2028–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeland, I.J.; Singh, S.; McGuire, D.K.; Vega, G.L.; Roddy, T.; Reilly, D.F.; Castro-Perez, J.; Kozlitina, J.; Scherer, P.E. Relation of plasma ceramides to visceral adiposity, insulin resistance and the development of type 2 diabetes mellitus: The Dallas Heart Study. Diabetologia 2018, 61, 2570–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, F.; Ho, C.T.; Sang, S.; Rosen, R.T. Determination of sphingolipids in nuts and seeds by a single quadrupole liquid chromatography-mass spectrometry method. J. Food Lipids 2005, 12, 327–343. [Google Scholar] [CrossRef]
- Michaelson, L.V.; Napier, J.A.; Molino, D.; Faure, J.D. Plant sphingolipids: Their importance in cellular organization and adaption. Biochim. Biophys. Acta 2016, 1861, 1329–1335. [Google Scholar]
- Miraliakbari, H.; Shahidi, F. Antioxidant activity of minor components of tree nut oils. Food Chem. 2008, 111, 421–427. [Google Scholar] [CrossRef]
- Paroni, R.; Dei Cas, M.; Rizzo, J.; Ghidoni, R.; Montagna, M.T.; Rubino, F.M.; Iriti, M. Bioactive phytochemicals of tree nuts. Determination of the melatonin and sphingolipid content in almonds and pistachios. J. Food Compos. Anal. 2019, 82, 103227. [Google Scholar] [CrossRef]
- Sang, S.; Kikuzaki, H.; Lapsley, K.; Rosen, R.T.; Nakatani, N.; Ho, C.T. Sphingolipid and other constituents from almond nuts (Prunus amygdalus Batsch). J. Agric. Food Chem. 2002, 50, 4709–4712. [Google Scholar] [CrossRef]
- Rubino, F.M.; Zecca, L.; Sonnino, S. Characterization of a Complex Mixture of Ceramides by Fast-Atom-Bombardment and Precursor and Fragment Analysis Tandem Mass-Spectrometry. Biol. Mass Spectrom. 1994, 23, 82–90. [Google Scholar] [CrossRef]
- Reisberg, M.; Arnold, N.; Porzel, A.; Neubert, R.H.H.; Dräger, B. Production of Rare Phyto-Ceramides from Abundant Food Plant Residues. J. Agric. Food Chem. 2017, 65, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Sesana, S.; Ghidoni, R.; Monti, M. Content of the different lipid classes in psoriatic scale. Arch. Dermatol. Res. 1995, 287, 691–694. [Google Scholar] [CrossRef]
- Gaudin, K.; Chaminade, P.; Baillet, A. Structure-retention diagrams of ceramides established for their identification. J. Chromatogr. A 2002, 973, 69–83. [Google Scholar] [CrossRef]
- Murphy, R.C.; James, P.F.; McAnoy, A.M.; Krank, J.; Duchoslav, E.; Barkley, R.M. Detection of the abundance of diacylglycerol and triacylglycerol molecular species in cells using neutral loss mass spectrometry. Anal. Biochem. 2007, 366, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Goodacre, R.; Vaidyanathan, S.; Bianchi, G.; Kell, D.B. Metabolic profiling using direct infusion electrospray ionisation mass spectrometry for the characterisation of olive oils. Analyst 2002, 127, 1457–1462. [Google Scholar] [CrossRef] [PubMed]
- Gorrepati, K.; Balasubramanian, S.; Chandra, P. Plant based butters. J. Food Sci. Technol. 2015, 52, 3965–3976. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, A.N.; Akinci-Yildirim, F.; Şan, B.; Sesli, Y. Total Oil Content and Fatty Acid Profile of some Almond (Amygdalus communis L.) Cultivars. Pol. J. Food Nutr. Sci. 2016, 66, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Moayedi, A.; Rezaei, K.; Moini, S.; Keshavarz, B. Chemical compositions of oils from several wild almond species. J. Am. Oil Chem. Soc. 2011, 88, 503–508. [Google Scholar] [CrossRef]
- Fernandes, G.D.; Gómez-Coca, R.B.; Pérez-Camino, M.D.C.; Moreda, W.; Barrera-Arellano, D. Chemical Characterization of Major and Minor Compounds of Nut Oils: Almond, Hazelnut, and Pecan Nut. J. Chem. 2017, 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Evoli, L.D.; Lucarini, M.; Gabrielli, P.; Aguzzi, A.; Lombardi-boccia, G. Nutritional Value of Italian Pistachios from Bronte (Pistacia vera, L.), Their Nutrients, Bioactive Compounds and Antioxidant Activity. Food Nutr. Sci. 2015, 6, 1267. [Google Scholar]
- Alasalvar, C.; Shahidi, F. Tree Nuts: Composition, Phytochemicals, and Health Effects: An Overview. In Tree Nuts: Composition, Phytochemicals, and Health Effects; CRC Press: Boca Raton, FL, USA, 2008; ISBN 978-1-4200-1939-1. [Google Scholar]
- Valsecchi, M.; Mauri, L.; Casellato, R.; Ciampa, M.G.; Rizza, L.; Bonina, A.; Bonina, F.; Sonnino, S. Ceramides as possible nutraceutical compounds: Characterization of the ceramides of the moro blood orange (Citrus sinensis). J. Agric. Food Chem. 2012, 60, 10103–10110. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Wu, X.; Zhu, M.; Zhao, W.; Li, F.; Zou, Y.; Yang, L. Chemical composition, nutritional value, and antioxidant activities of eight mulberry cultivars from China. Pharmacogn. Mag. 2012, 8, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteki, M.; Ahmadi, P.; Vander Heyden, Y.; Simal-Gandara, J. Fatty acids-based quality index to differentiate worldwide commercial pistachio cultivars. Molecules 2019, 24, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, N.; Benetti, G.; Campolongo, G.; Ferrari, G.; Palermo, R.; Vranic, B. Preliminary Assessment of Vegetable Oil Adulteration of Pistachio Paste by near Infrared Spectroscopy. NIR News 2014, 25, 20–21. [Google Scholar] [CrossRef]
- Eksi-Kocak, H.; Mentes-Yilmaz, O.; Boyaci, I.H. Detection of green pea adulteration in pistachio nut granules by using Raman hyperspectral imaging. Eur. Food Res. Technol. 2016, 242, 271–277. [Google Scholar] [CrossRef]
- Sezer, B.; Apaydin, H.; Bilge, G.; Boyaci, I.H. Detection of Pistacia vera adulteration by using laser induced breakdown spectroscopy. J. Sci. Food Agric. 2018, 99, 2236–2242. [Google Scholar] [CrossRef]
- Çavuş, F.; Us, M.F.; Güzelsoy, N.A. Assesing Pistachio Nut (Pistacia vera L.) Adulteration with Green Pea (Pisum sativum L.) by Untargeted Liquid Chromatography-(Quadrupole-Time of Flight)-Mass Spectrometry Method and Chemometrics. Available online: https://dergipark.org.tr/en/pub/bursagida/issue/40169/477784 (accessed on 20 January 2020).
- Canela, N.; Herrero, P.; Mariné, S.; Nadal, P.; Ras, M.R.; Rodríguez, M.Á.; Arola, L. Analytical methods in sphingolipidomics: Quantitative and profiling approaches in food analysis. J. Chromatogr. A 2016, 1428, 16–38. [Google Scholar] [CrossRef]
- Markham, J.E.; Jaworski, J.G. Rapid measurement of sphingolipids from Arabidopsis thaliana by reversed-phase high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 1304–1314. [Google Scholar] [CrossRef]
- Dalmau, N.; Jaumot, J.; Tauler, R.; Bedia, C. Epithelial-to-mesenchymal transition involves triacylglycerol accumulation in DU145 prostate cancer cells. Mol. Biosyst. 2015, 11, 3397–3406. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Small samples of some almond and pistachio specimens are available from the authors upon reasonable request. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Core Structure of Long Chain Base (LCB) | LCB (MW) and O” (m/z) |
|---|---|---|
| A |  | d18:2-4,8 C18H35NO2 MW 297.49 O” C18H32N]+ m/z 262.22 |
| B |  | d18:1-4 C18H37NO2 MW 299.50 O” C18H34N]+ m/z 264.22 |
| C |  | d18:0 C18H39NO2 MW 301.52 O” C18H36N]+ m/z 266.22 |
| D |  | t18:1-8 C18H37NO3 MW 315.50 O” C18H34NO]+ m/z 280.22 |
| E |  | t18:0 C18H39NO3 MW 317.52 O” C18H36NO]+ m/z 282.22 |
| R = H (ceramide); R = Hex (cerebroside); FA = fatty acid residue Fragment O” = [LCB − 2 H2O] × H+ |
| Sample ID | A1 | A2 |
|---|---|---|
| M1 | 13.65 | n.d. |
| M2 | 22.48 | n.d. |
| M3 | 36.47 | n.d. |
| P1 | 39.97 | 13.64 |
| P2 | 17.37 | 2.87 |
| P3 | 14.69 | 4.53 |
| P4 | 14.35 | 3.63 |
| P5 | 15.95 | 4.04 |
| P6 | 17.26 | 4.09 |
| P7 | 19.40 | 3.74 |
| P8 | 22.24 | 7.13 |
| P9 | 12.52 | 2.85 |
| ID | Sample | Nut Product | Origin | Characteristics |
| Pistachio (Pistacia vera L.) | ||||
| P1 | P7 | Shelled | Bronte DOP (Sicily, Italy) | Not roasted, not salted |
| P2 | P8 | Shell | USA | Roasted, salted |
| P3 | P9 | Shell | Non EU | Organic, roasted, not salted |
| P4 | P10 | Shell | Iran | Not roasted, not salted |
| P5 | P11 | Shell | USA | Roasted, not salted |
| P6 | P12 | Shelled | Noberasco | Not roasted, not salted |
| P7 | P13 | Flour | Iran | |
| P8 | P14 | Flour | Italy | |
| P9 | P15 | Flour | Italy | |
| ID | Sample | Nut Product | Origin | Characteristics |
| Almond (Prunus dulcis Mill.) | ||||
| M1 | A6 | Shelled | California (USA) | Died |
| M2 | A7 | Shelled | California (USA) | Dried |
| M3 | A8 | Shelled | California (USA) | Dried |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubino, F.M.; Dei Cas, M.; Bignotto, M.; Ghidoni, R.; Iriti, M.; Paroni, R. Discovery of Unexpected Sphingolipids in Almonds and Pistachios with an Innovative Use of Triple Quadrupole Tandem Mass Spectrometry. Foods 2020, 9, 110. https://doi.org/10.3390/foods9020110
Rubino FM, Dei Cas M, Bignotto M, Ghidoni R, Iriti M, Paroni R. Discovery of Unexpected Sphingolipids in Almonds and Pistachios with an Innovative Use of Triple Quadrupole Tandem Mass Spectrometry. Foods. 2020; 9(2):110. https://doi.org/10.3390/foods9020110
Chicago/Turabian StyleRubino, Federico Maria, Michele Dei Cas, Monica Bignotto, Riccardo Ghidoni, Marcello Iriti, and Rita Paroni. 2020. "Discovery of Unexpected Sphingolipids in Almonds and Pistachios with an Innovative Use of Triple Quadrupole Tandem Mass Spectrometry" Foods 9, no. 2: 110. https://doi.org/10.3390/foods9020110
APA StyleRubino, F. M., Dei Cas, M., Bignotto, M., Ghidoni, R., Iriti, M., & Paroni, R. (2020). Discovery of Unexpected Sphingolipids in Almonds and Pistachios with an Innovative Use of Triple Quadrupole Tandem Mass Spectrometry. Foods, 9(2), 110. https://doi.org/10.3390/foods9020110