Table Olives More than a Fermented Food


 ,
,
Abstract
:1. Introduction
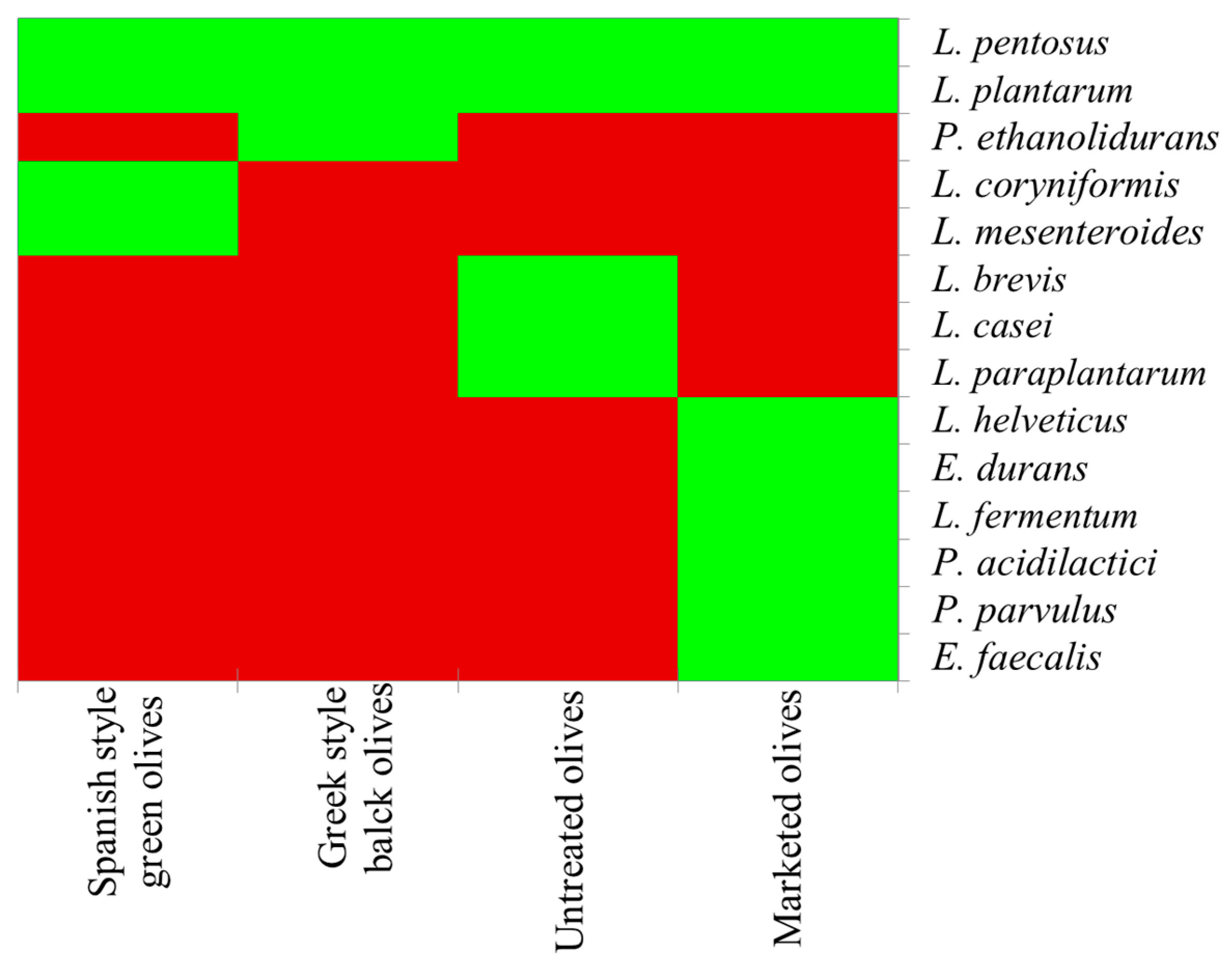
2. Table Olives Associated Microbiota
3. Microbial Spoilage of Table Olives
4. Table Olives’ Starter Cultures
5. New Trend in Olive Production: Probiotic Table Olives
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- IOOC, International Olive Oil Council. Trade Standard Applying to Table Olives; International Olive Oil Council: Madrid, Spain, 2004. [Google Scholar]
- IOOC, International Olive Oil Council. World Table Olive Figures. 2019. Available online: http://www.internationaloliveoil.org/estaticos/view/132-world-table-olive-figures (accessed on 20 January 2019).
- Campus, M.; Degirmencioglu, N.; Comunian, R. Technologies and Trends to Improve Table Olive Quality and Safety. Front. Microbiol. 2018, 9, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Fernández, A.; Fernández-Díez, M.J.; Adams, R.M. Table Olives: Production and Processing, 1st ed.; Chapman and Hall: London, UK, 1997. [Google Scholar]
- Rejano, L.; Montaño, A.; Casado, F.J.; Sánchez, A.H.; de Castro, A. Table olives: Varieties and variations. In Olives and Olive Oil in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Eds.; Oxford Academic Press: Oxford, UK, 2010; pp. 5–15. [Google Scholar]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.J.E. Table olive fermentation using starter cultures with multifunctional potential. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Cabello, A.; Romero-Gil, V.; Rodríguez-Gómez, F.; Garrido-Fernández, A.; Jiménez-Díaz, R.; Arroyo-López, F.N. Evaluation and identification of poly-microbial biofilms on natural green Gordal table olives. Antonie Van Leeuwenhoek 2015, 108, 597–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randazzo, C.L.; Restuccia, C.; Romano, A.D.; Caggia, C. Lactobacillus casei, dominant species in naturally fermented Sicilian green olives. Int. J. Microbiol. 2004, 90, 9–14. [Google Scholar] [CrossRef]
- De Bellis, P.; Valerio, F.; Sisto, A.; Lonigro, S.L.; Lavermicocca, P. Probiotic table olives: Microbial populations adhering on olive surface in fermentation sets inoculated with the probiotic strain Lactobacillus paracasei IMPC2.1 in an industrial plant. Int. J. Food Microbiol. 2010, 140, 6–13. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Altieri, C.; Corbo, M.R.; Sinigaglia, M.; Ouoba, L.I.I. Characterization of lactic acid bacteria isolated from Italian Bella di Cerignola table olives: Selection of potential multifunctional starter cultures. J. Food Sci. 2010, 75, 536–544. [Google Scholar] [CrossRef]
- Franzetti, A.; Gandolfi, I.; Gaspari, E.; Ambrosini, R.; Bestetti, G. Seasonal variability of bacteria in fine and coarse urban air particulate matter. Appl. Microbiol. Biotechnol. 2011, 90, 745–753. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G.; La Croce, F.; Mazzaglia, A.; Farina, V.; Settanni, L. Use of selected autochthonous lactic acid bacteria for Spanish style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Esteve-Zarzoso, B.; Bordons, A.; Rozès, N. Microbial population dynamics during the processing of Aberquina table olives. Food Res. Int. 2008, 41, 738–744. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Hondrodimou, O.; Iliopoulos, V.; Panagou, E.Z. Lactic acid bacteria and yeast heterogeneity during aerobic and modified atmosphere packaging storage of natural black Conservolea olives in polyethylene pouches. Food Control 2012, 26, 49–57. [Google Scholar] [CrossRef]
- Tofalo, R.; Perpetuini, G.; Schirone, M.; Ciarrocchi, A.; Fasoli, G.; Suzzi, G.; Corsetti, A. Lactobacillus pentosus dominates spontaneous fermentation of Italian table olives. LWT Food Sci. Technol. 2014, 57, 710–717. [Google Scholar] [CrossRef]
- Campaniello, D.; Bevilacqua, A.; D’Amato, D.; Corbo, M.R.; Altieri, C.; Sinigaglia, M. Microbial characterization of table olives processed according to Spanish and natural styles. Food Technol. Biotechnol. 2005, 43, 289–294. [Google Scholar]
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of starter cultures to table olive fermentation: An overview on the experimental studies. Front. Microbiol. 2012, 3, 248. [Google Scholar] [CrossRef] [Green Version]
- Caggia, C.; Randazzo, C.L.; Di Salvo, M.; Romeo, F.V.; Giudici, P. Occurrence of Listeria monocytogenes in green table olives. J. Food Prot. 2004, 67, 2189–2194. [Google Scholar] [CrossRef]
- Alves, M.; Gonçalves, T.; Quintas, C. Microbial quality and yeast population dynamics in cracked green table olives’ fermentations. Food Control 2012, 23, 363–368. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Querol, A.; Bautista-Gallego, J.; Garrido-Fernández, A. Role of yeasts in table olive production. Int. J. Food Microbiol. 2008, 128, 189–196. [Google Scholar] [CrossRef]
- Hernández, A.; Martín, A.; Aranda, E.; Pérez-Nevado, F.; Córdoba, M.G. Identification and characterization of yeast isolated from the elaboration of seasoned green table olives. Food Microbiol. 2007, 24, 346–351. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Rodriguez-Gomez, F.; Barrio, E.; Querol, A.; Garrido- Fernandez, A.; Arroyo-López, F.N. Exploring the yeast biodiversity of green table olive industrial fermentations for technological applications. Int. J. Food Microbiol. 2011, 147, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-López, F.N.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; García-García, P.; Querol, A.; Garrido-Fernández, A. Potential benefits of the application of yeast starters in table olive processing. Front. Microbiol. 2012, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Perpetuini, G.; Schirone, M.; Suzzi, G.; Corsetti, A. Yeast biota associated to naturally fermented table olives from different Italian cultivars. Int. J. Food Microbiol. 2013, 161, 203–208. [Google Scholar] [CrossRef]
- Hernández, A.; Martín, A.; Córdoba, M.G.; Benito, M.J.; Aranda, E.; Pérez-Nevado, F. Determination of killer activity in yeasts isolated from the elaboration of seasoned green table olives. Int. J. Food Microbiol. 2008, 121, 178–188. [Google Scholar] [CrossRef]
- Psani, M.; Kotzekidou, P. Technological characteristics of yeaststrains and their potential as starter adjuncts in Greek-style black olive fermentation. World J. Microbiol. Biotechnol. 2006, 22, 1329–1336. [Google Scholar] [CrossRef]
- Lucena-Padrós, H.; Ruiz-Barba, J.L. Microbial biogeography of Spanish-style green olive fermentations in the province of Seville, Spain. Food Microbiol. 2019, 82, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Lanza, B. Abnormal fermentations in table-olive processing: Microbial origin and sensory evaluation. Front. Microbiol. 2013, 4, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golomb, B.L.; Morales, V.; Jung, A.; Yau, B.; Boundy-Mills, K.L.; Marco, M.L. Effects of pectinolytic yeast on the microbial composition and spoilage of olive fermentations. Food Microbiol. 2013, 33, 97–106. [Google Scholar] [CrossRef]
- Kailis, S.; Harris, D. Producing Table Olives; Landlinks Press: Collingwood, VIC, Australia, 2007; p. 328. [Google Scholar]
- Montano, A.; de Castro, A.; Rejano, L.; Brenes, M. 4- hydroxycyclohexanecarboxylic acid as a substrate for cyclohexane carboxylic acid production during the “Zapatera” spoilage of Spanish-style green table olives. J. Food Prot. 1996, 59, 657–662. [Google Scholar] [CrossRef]
- Garcia, P.G.; Barranco, C.R.; Quintana, M.C.; Fernandez, A.G. Biogenic amine formation and “zapatera” spoilage of fermented green olives: Effect of storage temperature and debittering process. J. Food Prot. 2004, 67, 117–123. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Campanella, D.; Cosmai, L.; Summo, C.; Rizzello, C.G.; Caponio, F. Microbiota and metabolome of un-started and started Greek-type fermentation of Bella di Cerignola table olives. Food Microbiol. 2015, 52, 18–30. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Todaro, A.; Pino, A.; Corona, O.; Caggia, C. Microbiota and metabolome during controlled and spontaneous fermentation of Nocellara Etnea table olives. Food Microbiol. 2017, 65, 136–148. [Google Scholar] [CrossRef]
- Perpetuini, G.; Caruso, G.; Urbani, S.; Schirone, M.; Esposto, S.; Ciarrocchi, A.; Prete, R.; Garcia-Gonzalez, N.; Battistelli, N.; Gucci, R.; et al. Changes in polyphenolic concentrations of table olives (cv. Itrana) produced under different irrigation regimes during spontaneous or inoculated fermentation. Front. Microbiol. 2018, 9, 1287. [Google Scholar] [CrossRef]
- Martorana, A.; Alfonzo, A.; Settanni, L.; Corona, O.; La Croce, F.; Caruso, T.; Moschetti, G.; Francesca, N. An innovative method to produce green table olives based on “pied de cuve” technology. Food Microbiol. 2015, 50, 126–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marugg, J.D. Bacteriocins, their role in developing natural products. Food Biotechnol. 1991, 5, 305–312. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Comunian, R.; Ferrocino, I.; Paba, A.; Daga, E.; Campus, M.; Di Salvo, R.; Cauli, E.; Piras, F.; Zurru, E.; Cocolin, L. Evolution of microbiota during spontaneous and inoculated Tonda di Cagliari table olives fermentation and impact on sensory characteristics. LWT Food Sci. Technol. 2017, 84, 64–72. [Google Scholar] [CrossRef]
- Ramírez, E.; Medina, E.; García, P.; Brenes, M.; Romero, C. Optimization of the natural debittering of table olives. LWT Food Sci. Technol. 2017, 77, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Chranioti, C.; Kotzekidou, P.; Gerasopoulos, D. Effect of starter cultures on fermentation of naturally and alkali-treated cv. Conservolea green olives. LWT Food Sci. Technol. 2018, 89, 403–408. [Google Scholar] [CrossRef]
- Sherahi, M.H.A.; Shahidi, F.; Yazdi, F.T.; Hashemi, S.M.B. Effect of Lactobacillus plantarum on olive and olive oil quality during fermentation process. LWT Food Sci. Technol. 2018, 89, 572–580. [Google Scholar] [CrossRef]
- Pino, A.; Vaccalluzzo, A.; Solieri, L.; Romeo, F.V.; Todaro, A.; Caggia, C.; Arroyo-López, F.N.; Bautista-Gallego, J.; Randazzo, C.L. Effect of Sequential Inoculum of Beta-Glucosidase Positive and Probiotic Strains on Brine Fermentation to Obtain Low Salt Sicilian Table Olives. Front. Microbiol. 2019, 10, 174. [Google Scholar] [CrossRef]
- Caponio, F.; Difonzo, G.; Calasso, M.; Cosmai, L.; De Angelis, M. Effects of olive leaf extract addition on fermentative and oxidative processes of table olives and their nutritional properties. Food Res. Int. 2019, 116, 1306–1317. [Google Scholar] [CrossRef]
- Servili, M.; Settanni, L.; Veneziani, G.; Esposto, S.; Massitti, O.; Taticchi, A.; Urbani, S.; Montedoro, G.F.; Corsetti, A. The use of Lactobacillus pentosus 1MO to shorten the debittering process time of black table olives (Cv. Itrana and Leccino): A pilot-scale application. J. Agric. Food Chem. 2006, 54, 3869–3875. [Google Scholar] [CrossRef]
- Tassou, C.C.; Panagou, E.Z.; Katsaboxakis, K.Z. Microbiological and physicochemical changes of naturally black olives fermented at different temperatures and NaCl levels in the brines. Food Microbiol. 2002, 19, 605–615. [Google Scholar] [CrossRef]
- Ciafardini, G.; Marsilio, A.; Lanza, B.; Pozzi, N. Hydrolysis of oleuropein by Lactobacillus plantarum strains associated with olive fermentation. Appl. Environ. Microbiol. 1994, 60, 4142–4147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega Leal-Sánchez, M.; Ruiz-Barba, J.L.; Sánchez Gómez, A.H.; Rejano, L.; Jiménez-Dıíaz, R.; Garrido Fernández, A. Fermentation profile and optimization of green olive fermentation using Lactobacillus plantarum LPCO10 as a starter culture. Food Microbiol. 2003, 20, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Nisiotou, A.A.; Chorianopoulos, N.; Nychas, G.J.E.; Panagou, E.Z. Yeast heterogeneity during spontaneous fermentation of black Conservolea olives in different brine solutions. J. Appl. Microbiol. 2010, 108, 396–405. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Beneduce, L.; Sinigaglia, M.; Corbo, M.R. Selection of yeasts as starter cultures for table olives. J. Food Sci. 2013, 78, 742–751. [Google Scholar] [CrossRef]
- Bevilacqua, A.; De Stefano, F.; Augello, S.; Pignatiello, S.; Sinigaglia, M.; Corbo, M.R. Biotechnological innovations for table olives. Int. J. Food Sci. Nutr. 2015, 66, 127–131. [Google Scholar] [CrossRef]
- Grounta, A.; Doulgeraki, A.I.; Nychas, G.J.E.; Panagou, E.Z. Biofilm formation on Conservolea natural Black olives during single and combined inoculation with a functional Lactobacillus pentosus starter culture. Food Microbiol. 2016, 56, 35–44. [Google Scholar] [CrossRef]
- Tufariello, M.; Durante, M.; Ramires, F.A.; Grieco, F.; Tommasi, L.; Perbellini, E.; Falco, V.; Tasioula-Margari, M.; Logrieco, A.F.; Mita, G.; et al. New process for production of fermented black table olives using selected autochthonous microbial resources. Front. Microbiol. 2015, 6, 1007. [Google Scholar] [CrossRef] [Green Version]
- Bonatsou, S.; Benítez, A.; Rodríguez-Gómez, F.; Panagou, E.Z.; Arroyo-López, F.N. Selection of yeasts with multifunctional features for application as starters in natural black table olive processing. Food Microbiol. 2015, 46, 66–73. [Google Scholar] [CrossRef]
- Domínguez-Manzano, J.; Olmo-Ruiz, C.; Bautista-Gallego, J.; Arroyo-López, F.N.; Garrido Fernández, A.; Jiménez-Díaz, R. Biofilm formation on abiotic and biotic surfaces during Spanish style green table olive fermentation. Int. J. Food Microbiol. 2012, 157, 230–238. [Google Scholar] [CrossRef]
- Grounta, A.; Panagou, E.Z. Mono and dual species biofilm formation between Lactobacillus pentosus and Pichia membranifaciens on the surface of black olives under different sterile brine conditions. Ann. Microbiol. 2014, 64, 1757–1767. [Google Scholar] [CrossRef]
- Grounta, A.; Doulgeraki, A.I.; Panagou, E.Z. Quantification and characterization of microbial biofilm community attached on the surface of fermentation vessels used in green table olive processing. Int. J. Food Microbiol. 2015, 203, 41–48. [Google Scholar] [CrossRef] [PubMed]
- León-Romero, Á.; Domínguez-Manzano, J.; Garrido-Fernández, A.; Arroyo-López, F.N.; Jiménez Díaz, R. Formation of in vitro mixed-species biofilms by Lactobacillus pentosus and yeasts isolated from Spanish-style green table olive fermentations. Appl. Environ. Microbiol. 2016, 82, 689–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porru, C.; Rodríguez-Gómez, F.; Benítez-Cabello, A.; Jiménez-Díaz, R.; Zara, G.; Budroni, M.; Mannazzu, I.; Arroyo-Lopez, F.N. Genotyping, identification and multifunctional features of yeasts associated to Bosana naturally black table olive fermentations. Food Microbiol. 2018, 69, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Medina, E.; Ruiz-Bellido, M.A.; Romero-Gil, V.; Rodríguez-Gómez, F.; Montes-Borrego, M.; Landa, B.B.; Arroyo-López, F.N. Assessment of the bacterial community in directly brined Aloreña de Málaga table olive fermentations by metagenetic analysis. Int. J. Food Microbiol. 2016, 236, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Campus, M.; Sedda, P.; Cauli, E.; Piras, F.; Comunian, R.; Paba, A.; Daga, E.; Schirru, S.; Angioni, A.; Zurru, R.; et al. Evaluation of a single strain starter culture, a selected inoculum enrichment, and natural microflora in the processing of Tonda di Cagliari natural table olives: Impact on chemical, microbiological, sensory and texture quality. LWT Food Sci. Technol. 2015, 64, 671–677. [Google Scholar] [CrossRef]
- Tataridou, M.; Kotzekidou, P. Fermentation of table olives by oleuropeinolytic starter culture in reduced salt brines and inactivation of Escherichia coli O157:H7 and Listeria monocytogenes. Int. J. Food Microbiol. 2015, 208, 122–130. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Use of Lactobacillus plantarum and glucose to control the fermentation of “Bella di Cerignola” Table Olives, a traditional variety of Apulian region (Southern Italy). J. Food Sci. 2010, 75, 430–436. [Google Scholar] [CrossRef]
- Perricone, M.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Use of starter cultures in olives: A not-correct use could cause a delay of performances. Food Nutr. Sci. 2013, 4, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Cosmai, L.; Campanella, D.; De Angelis, M.; Summo, C.; Paradiso, V.M.; Pasqualone, A.; Caponio, F. Use of starter cultures for table olives fermentation as possibility to improve the quality of thermally stabilized olive-based paste. LWT Food Sci. Technol. 2018, 90, 381–388. [Google Scholar] [CrossRef]
- Benincasa, C.; Muccilli, S.; Amenta, M.; Perri, E.; Romeo, F.V. Phenolic trend and hygienic quality of green table olives fermented with Lactobacillus plantarum starter culture. Food Chem. 2015, 186, 271–276. [Google Scholar] [CrossRef]
- Argyri, A.A.; Nisiotou, A.A.; Mallouchos, A.; Panagou, E.Z.; Tassou, C.C. Performance of two potential probiotic Lactobacillus strains from the olive microbiota as starters in the fermentation of heat shocked green olives. Int. J. Food Microbiol. 2014, 171, 68–76. [Google Scholar] [CrossRef]
- Blana, V.A.; Grounta, A.; Tassou, C.C.; Nychas, G.J.E.; Panagou, E.Z. Inoculated fermentation of green olives with potential probiotic Lactobacillus pentosus and Lactobacillus plantarum starter cultures isolated from industrially fermented olives. Food Microbiol. 2014, 38, 208–218. [Google Scholar] [CrossRef]
- Blana, V.A.; Polymeneas, N.; Tassou, C.C.; Panagou, E.Z. Survival of potential probiotic lactic acid bacteria on fermented green table olives during packaging in polyethylene pouches at 4 and 20 °C. Food Microbiol. 2016, 53, 71–75. [Google Scholar] [CrossRef]
- Ruiz-Barba, J.L.; Caballero-Guerrero, B.; Maldonado-Barragán, A.; Jiménez-Díaz, R. Coculture with specific bacteria enhances survival of Lactobacillus plantarum NC8, an autoinducer-regulated bacteriocin producer, in olive fermentations. Food Microbiol. 2010, 27, 413–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltsa, A.; Papaliaga, D.; Papaioannou, E.; Kotzekidou, P. Characteristics of oleuropeinolytic strains of Lactobacillus plantarum group and influence on phenolic compounds in table olives elaborated under reduced salt conditions. Food Microbiol. 2015, 48, 58–62. [Google Scholar] [CrossRef]
- Lamzira, Z.; Asehraou, A.; Brito, D.; Oliveira, M.; Faid, M.; Peres, C. Bloater spoilage of greenolives. Food Technol. Biotechnol. 2005, 43, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Evaluation of a single and combined inoculation of a Lactobacillus pentosus starter for processing cv. Arbequina natural green olives. Food Microbiol. 2010, 27, 731–740. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Romero-Gil, V.; Rodríguez-Gómez, F.; Garcia-Garcia, P.; Garrido-Fernández, A. Chloride salt mixtures affect Gordal cv. green Spanish-style table olive fermentation. Food Microbiol. 2011, 28, 1316–1325. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Bautista-Gallego, J.; Arroyo-López, F.N.; Romero-Gil, V.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Garcia-Garcia, P. Table olive fermentation with multifunctional Lactobacillus pentosus strains. Food Control 2013, 34, 96–105. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Bautista-Gallego, J.; García-García, P.; Garrido-Fernández, A.; Arroyo-López, F.N. Production of potential probiotic Spanish-style green table olives at pilot plant scale using multifunctional starters. Food Microbiol. 2014, 44, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Arroyo-López, F.N.; Roldán-Reyes, J.C.; Torres-Gallardo, R.; Bautista-Gallego, J.; Garcia-Garcia, P.; Garrido-Fernandez, A. Assessing the challenges in the application of potential probiotic lactic acid bacteria in the large-scale fermentation of Spanish-style table olives. Front. Microbiol. 2017, 8, 915. [Google Scholar] [CrossRef] [Green Version]
- López-López, A.; Moreno-Baquero, J.M.; Rodríguez-Gómez, F.; García-García, P.; Garrido-Fernández, A. Sensory Assessment by Consumers of Traditional and Potentially Probiotic Green Spanish-Style Table Olives. Front. Nutr. 2018, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- de Castro, A.; Sánchez, A.H.; Cortés-Delgado, A.; López-López, A.; Montaño, A. Effect of Spanish-style processing steps and inoculation with Lactobacillus pentosus starter culture on the volatile composition of cv. Manzanilla green olives. Food Chem. 2019, 271, 543–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorana, A.; Alfonzo, A.; Gaglio, R.; Settanni, L.; Corona, O.; La Croce, F.; Vagnoli, P.; Caruso, T.; Moschetti, N.; Francesca, N. Evaluation of different conditions to enhance the performances of Lactobacillus pentosus OM13 during industrial production of Spanish-style table olives. Food Microbiol. 2017, 61, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Campus, M.; Cauli, E.; Scano, E.; Piras, F.; Comunian, R.; Paba, A.; Daga, E.; Di Salvo, R.; Sedda, P.; Angioni, A.; et al. Towards controlled fermentation of table olives: Lab starter driven process in an automatic pilot processing plant. Food Bioprocess Technol. 2017, 10, 1063–1073. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Todaro, A.; Pino, A.; Pitino, I.; Corona, O.; Mazzaglia, A.; Caggia, C. Giarraffa and Grossa di Spagna naturally fermented table olives: effect of starter and probiotic cultures on chemical, microbiological and sensory traits. Food Res. Int. 2014, 62, 1154–1164. [Google Scholar] [CrossRef]
- Ciafardini, G.; Zullo, B.A. Use of selected yeast starter cultures in industrial-scale processing of brined Taggiasca black table olives. Food Microbiol. 2019, 84, 103250. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Fava, G.; Tomaselli, F.; Romeo, F.V.; Pennino, G.; Vitello, E.; Caggia, C. Effect of kaolin and copper based products and of starter cultures on green table olive fermentation. Food Microbiol. 2011, 28, 910–919. [Google Scholar] [CrossRef]
- Pino, A.; De Angelis, M.; Todaro, A.; Van Hoorde, K.; Randazzo, C.L.; Caggia, C. Fermentation of Nocellara Etnea Table Olives by Functional Starter Cultures at Different Low Salt Concentrations. Front. Microbiol. 2018, 9, 1125. [Google Scholar] [CrossRef] [Green Version]
- Durante, M.; Tufariello, M.; Tommasi, L.; Lenucci, M.S.; Bleve, G.; Mita, G. Evaluation of bioactive compounds in black table olives fermented with selected microbial starters. J. Sci. Food Agric. 2017, 98, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; González-Ortiz, G.; Benítez-Cabello, A.; Calero-Delgado, B.; Jiménez-Díaz, R.; Martín-Orúe, S.M. The use of starter cultures in the table olive fermentation can modulate antiadhesive properties of brine exopolysaccharides against enterotoxigenic Escherichia coli. Food Funct. 2019, 10, 3738–3747. [Google Scholar] [CrossRef]
- Shah, P.N. Functional cultures and health benefits. Int. Dairy J. 2007, 17, 1262–1277. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animal. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [PubMed]
- Fuller, R.; Gibson, G.R. Probiotics and prebiotics: microflora management for improved gut health. Clin. Microbiol. Infect. 1998, 4, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Gorbach, S.L. Probiotics in the third millennium. Dig. Liver Dis. 2002, 34, S2–S7. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Schillinger, U. Introduction to pre- and probiotics. Food Res. Int. 2002, 35, 109–116. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2002, 334, 1–15. [Google Scholar] [CrossRef] [Green Version]
- FAO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO: Rome, Italy, 2006; p. 85. [Google Scholar]
- Haller, D.; Colbus, H.; Ganzle, M.G.; Scherenbacher, P.; Bode, C.; Hammes, W.P. Metabolic and functional properties of lactic acid bacteria in the gastro-intestinal ecosystem: A comparative in vitro study between bacteria of intestinal and fermented food origin. Syst. Appl. Microbiol. 2001, 24, 218–226. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Varma, Y.; Fischbach, M.A.; Biddinger, S.B.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, A.M.; O’Regan, P.; Fanning, A.; O’Mahony, C.; Macsharry, J.; Lyons, A.; Bienenstock, J.; O’Mahony, L.; Shanahan, F. Functional modulation of human intestinal epithelial cell responses by Bifidobacterium infantis and Lactobacillus salivarius. Immunology 2006, 118, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Botta, C.; Langerholc, T.; Cencic, A.; Cocolin, L. In vitro selection and characterization of new probiotic candidates from table olive microbiota. PLoS ONE 2014, 9, e94457. [Google Scholar] [CrossRef] [PubMed]
- Pérez Montoro, B.; Benomar, N.; Lavilla Lerma, L.; Castillo Gutiérrez, S.; Gálvez, A.; Abriouel, H. Fermented Aloreña table olives as a source of potential probiotic Lactobacillus pentosus strains. Front. Microbiol. 2016, 7, 1583. [Google Scholar] [CrossRef] [Green Version]
- Prete, R.; Tofalo, R.; Federici, E.; Ciarrocchi, A.; Cenci, G.; Corsetti, A. Food-associated Lactobacillus plantarum and yeasts inhibit the genotoxic effect of 4-Nitroquinoline-1-Oxide. Front. Microbiol. 2017, 8, 2349. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gonzalez, N.; Prete, R.; Battista, N.; Corsetti, A. Adhesion Properties of Food-Associated Lactobacillus plantarum Strains on Human Intestinal Epithelial Cells and Modulation of IL-8 Release. Front. Microbiol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Prete, R.; Long, S.L.; Gallardo, A.L.; Gahan, C.G.; Corsetti, A.; Joyce, S.A. Beneficial bile acid metabolism from Lactobacillus plantarum of food origin. Sci. Rep. 2020, 10, 1165. [Google Scholar] [CrossRef] [Green Version]
- Delgado, A.; Brito, D.; Peres, C.; Noe-Arroyo, F.; Garrido-Fernández, A. Bacteriocin production by Lactobacillus pentosus B96 can be expressed as a function of temperature and NaCl concentration. Food Microbiol. 2005, 22, 521–528. [Google Scholar] [CrossRef]
- Kacem, M.; Karam, N.E. Microbiological study of naturally fermented Algerian green olives: Isolation and identification of lactic acid bacteria and yeasts along with the effects of brine solutions obtained at the end of olive fermentation on Lactobacillus plantarum growth. Grasas y Aceites 2006, 57, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Mokhbi, A.; Kaid-Harche, M.; Lamri, K.; Rezki, M.; Kacem, M. Selection of Lactobacillus plantarum strains for their use as starter culturesin Algerian olive fermentations. Grasas y Aceites 2009, 60, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Guantario, B.; Zinno, P.; Schifano, E.; Roselli, M.; Perozzi, G.; Palleschi, C.; Uccelletti, D.; Devirgiliis, C. In vitro and in vivo selection of potentially probiotic lactobacilli from Nocellara del Belice table olives. Front. Microbiol. 2018, 9, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Cabello, A.; Torres-Maravilla, E.; Bermúdez-Humarán, L.; Langella, P.; Martín, R.; Jiménez-Díaz, R.; Arroyo-López, F.N. Probiotic properties of Lactobacillus strains isolated from table olive biofilms. Probiotics Antimicrob. Proteins 2019. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; Lonigro, S.L.; de Angelis, M.; Morelli, L.; Callegari, M.L. Study of adhesion and survival of lactobacilli and bifidobacteria on table olives with the aim of formulating a new probiotic food. Appl. Environ. Microbiol. 2005, 71, 4233–4240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valerio, F.; de Candia, S.; Lonigro, S.L.; Russo, F.; Riezzo, G.; Orlando, A.; De Bellis, P.; Sisto, A.; Lavermicocca, P. Role of the probiotic strain Lactobacillus paracasei LMGP22043 carried by artichokes in influencing faecal bacteria and biochemical parameters in human subjects. J. Appl. Microbiol. 2011, 111, 155–164. [Google Scholar] [CrossRef]
- Argyri, A.A.; Nisiotou, A.A.; Pramateftaki, P.; Doulgeraki, A.I.; Panagou, E.Z.; Tassou, C.C. Preservation of green table olives fermented with lactic acid bacteria with probiotic potential under modified atmosphere packaging. LWT Food Sci. Technol. 2015, 62, 783–790. [Google Scholar] [CrossRef]
- Perpetuini, G.; Pham-Hoang, B.N.; Scornec, H.; Tofalo, R.; Schirone, M.; Suzzi, G.; Cavin, J.F.; Waché, Y.; Corsetti, A.; Licandro-Seraut, H. In Lactobacillus pentosus, the olive brine adaptation genes are required for biofilm formation. Int. J. Food Microbiol. 2016, 216, 104–109. [Google Scholar] [CrossRef]
- Pérez Montoro, B.; Benomar, N.; Caballero Gómez, N.; Ennahar, S.; Horvatovich, P.; Knapp, C.W.; Alonso, E.; Gálvez, A.; Abriouel, H. Proteomic analysis of Lactobacillus pentosus for the identification of potential markers of adhesion and other probiotic features. Food Res. Int. 2018, 111, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abriouel, H.; Pérez Montoro, B.; Casimiro-Soriguer, C.S.; Pérez Pulido, A.J.; Knapp, C.W.; Caballero Gómez, N.; Castillo-Gutiérrez, S.; Estudillo-Martínez, M.D.; Gálvez, A.; Benomar, N. Insight into potential probiotic markers predicted in Lactobacillus pentosus MP-10 genome sequence. Front. Microbiol. 2017, 8, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calero-Delgado, B.; Pérez-Pulido, A.J.; Benítez-Cabello, A.; Martín-Platero, A.M.; Casimiro-Soriguer, C.S.; Martínez-Bueno, M.; Arroyo-López, F.N.; Jiménez Díaz, R. Multiple genome sequences of Lactobacillus pentosus strains isolated from biofilms on the skin of fermented green table olives. Microbiol. Resour. Announc. 2019, 8, e01546-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, E.M.F.; Ramos, A.M.; Vanzela, E.S.L.; Stringheta, P.C.; Pinto, C.L.O.; Martins, J.M. Products of vegetable origin: A new alternative for the consumption of probiotic bacteria. Food Res. Int. 2013, 51, 764–770. [Google Scholar] [CrossRef]
- Sazawal, S.; Hiremath, G.; Dhingra, U.; Malik, P.; Deb, S.; Black, R.E. Efficacy of probiotics in prevention of acute diarrhoea: A meta-analysis of masked, randomised, placebo-controlled trials. Lancet Infect. Dis. 2006, 6, 374–382. [Google Scholar] [CrossRef]
- Pennacchia, C.; Blaiotta, G.; Pepe, O.; Villani, F. Isolation of Saccharomyces cerevisiae strains from different food matrices and their preliminary selection for a potential use as probiotics. J. Appl. Microbiol. 2008, 105, 1919–1928. [Google Scholar] [CrossRef]
- Kourelis, A.; Kotzamanidis, C.; Litopoulou-Tzanetaki, E.; Scouras, Z.G.; Tzanetakis, N.; Yiangou, M. Preliminary probiotic selection of dairy and human yeast strains. J. Biol. Res. 2010, 13, 93–104. [Google Scholar]
- Moslehi-Jenabian, S.; Lindegaard Pedersen, L.; Jespersen, L. Beneficial effects of probiotic and food borne yeasts on human health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef]
- Etienne-Mesmin, L.; Livrelli, V.; Privat, M.; Denis, S.; Cardot, J.M.; Alric, M.; Blanquet-Diot, S. Effect of a new probiotic Saccharomyces cerevisiae strain on survival of Escherichia coli O157:H7 in a dynamic gastrointestinal model. Appl. Environ. Microbiol. 2011, 77, 1127–1131. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.; Reto, M.; Sol, M.; Peito, A.; Peres, C.M.; Peres, C.; Malcata, F.X. Characterization of yeasts from Portuguese brined olives, with a focus on their potentially probiotic behaviour. LWT Food Sci. Technol. 2011, 44, 1349–1354. [Google Scholar] [CrossRef]
- Granato, D.; Branco, G.F.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional foods and non-dairy probiotic food development: Trends, concepts, and products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 292–302. [Google Scholar] [CrossRef]
- Vijaya Kumar, B.; Vijayendra, S.V.N.; Reddy, O.V.S. Trends in dairy and non-dairy probiotic products—A review. J. Food Sci. Technol. 2015, 52, 6112–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranadheera, S.; Baines, S.K.; Adams, M.C. Importance of food in probiotic efficacy. Food Res. Int. 2010, 43, 1–7. [Google Scholar] [CrossRef]
- Lamsal, B.P.; Faubion, J. The beneficial use of cereal and cereal components in probiotic foods. Food Rev. Int. 2009, 25, 103–114. [Google Scholar] [CrossRef]


{kind=link}
| Preparation Method | Process |
|---|---|
| Treated olives | It is applied to green olives, olives turning color, or black olives. Olive debittering is achieved through an alkaline treatment (lye 2.5%–3% w/v). Olives are then placed in brine (NaCl 10%–11% w/v) where the fermentation takes place and lasts 3–7 months. Fermentation is driven by lactic acid bacteria. |
| Natural olives | It is applied to green olives, olives turning color. or black olives. Olives are placed directly in brine. With a salt concentration of about 6%–10% (w/v). Oleuropein is removed through the enzymatic activities (mainly β-glucosidase and esterase) of indigenous microorganisms. The fermentation process can last 8–12 months and it is mainly driven by yeasts and lactic acid bacteria. |
| Dehydrated and/or shriveled olives | It is applied to green olives, olives turning color, or black olives. Olives are subjected or not to a mild alkaline treatment, preserved in brine, or partially dehydrated in dry salt and/or by heating. |
| Olives darkened by oxidation | It is applied to green olives or olives turning color. Olives are preserved in brine, fermented or not, and darkened by oxidation in an alkaline medium. They are stored in hermetically sealed containers and subjected to heat sterilization. |
| Specialties | Olives prepared in a different way than those above following traditional recipes. |
| Properties | Characteristics | Advantages |
|---|---|---|
| Safety | Safe and stable activity Standardized activity Easy to manage and reproduce | Reproducibility Controlled and stable fermentation Continuous monitoring of fermentation |
| Technological | Ability to colonize olives surface (i.e., biofilm formation) Low demand for nutrients | Rapid and predominant growth High adaptation ability |
| Growth at different pH (high/low) Salt tolerance Ability to survive/growth at low temperatures | Dominance during the fermentation | |
| Biodegradation of phenolic compounds Debittering activity (i.e., oleuropeinolytic activity) High acidification activity | Reduction of fermentation time Avoided use of chemicals (microbial biotransformation) | |
| Functional | Antimicrobial activity vs. pathogens (i.e., bacteriocins production, competitive action on nutrients) Biocontrol agents vs. spoilage microorganisms (i.e., production of killer factors) | Protection from undesirable and/or pathogenic microorganisms Improvement of final product stability and shelf-life extension |
| Enzymatic activities (i.e., lipase, alkaline/acid phosphatase, β-glucosidase) Vitamins production Production of aromatic compounds | Enhancement of organoleptic, nutritional and sensory profile of the final product | |
| Probiotic | Survival under gastrointestinal conditions (i.e., low pH, gastric and pancreatic digestion, bile salts) Ability to adhere and persist in the intestinal mucosa Modulation of host immune system Antimicrobial activity against pathogens | Ensuring product safety Quality enhancement of the final product Production of a health-promoting functional food |
| Bacterial Starter Cultures | Cultivar | References |
|---|---|---|
| L. plantarum | Alorena | [40] |
| Bella di Cerignola | [33,63,64,65] | |
| Carolea/Cassanese | [66] | |
| Conservolea | [41] | |
| Gordal | [40] | |
| Halkidiki | [67,68,69] | |
| Hojiblanca | [40,70] | |
| Kalamata/Chalkidikis | [62,71] | |
| Manzanilla | [40] | |
| Mele | [28] | |
| Nocellara del Belice/Nocellara Messinese | [66] | |
| Nocellara Etnea | [34] | |
| Picholine | [72] | |
| Pishomi | [42] | |
| Tonda di Cagliari | [39,61] | |
| Leccino | [44] | |
| L. pentosus | Arbequina | [73] |
| Conservolea | [41,52] | |
| Gordal | [55,74] | |
| Halkidiki | [67,68,69] | |
| Itrana | [15,35] | |
| Manzanilla | [40,75,76,77,78,79] | |
| Nocellara del Belice | [36,80] | |
| Nocellara Etnea | [34] | |
| Tonda di Cagliari | [39,61,81] | |
| L. paracasei | Bella di Cerignola | [9] |
| L. rhamnosus | Giaraffa e Grossa di Spagna | [82] |
| Yeast starter cultures | Cultivar | References |
| N. molendini-olei/C. matritensis/C. adriatica/C. diddensiae/W. anomalus/S. cerevisiae | Taggiasca | [83] |
| Multi-starter Cultures | Cultivar | References |
|---|---|---|
| L. plantarum/L. pentosus | Bella di Cerignola | [65] |
| Halkidiki | [67,68] | |
| Nocellara Etnea | [34] | |
| L. plantarum/L. casei | Nocellara Etnea | [84] |
| L. plantarum/L. paracasei | Giaraffa e Grossa di Spagna | [82] |
| L. plantarum/L. paracasei | Nocellara Etnea | [43] |
| Nocellara Etnea | [34,85] | |
| L. plantarum/P. pentosaceus | Green olives | [70] |
| L. plantarum/E. faecieum | Green olives | [70] |
| L. paracasei/L. pentosus | Nocellara Etnea | [34] |
| L. pentosus/L. coryniformis | Nocellara del Belice | [12] |
| L. plantarum/L. paracasei/L. rhamnosus | Giaraffa e Grossa di Spagna | [82] |
| L. plantarum/L. paracasei/L. pentosus | Nocellara Etnea | [34] |
| L. plantarum/D. hansenii | Conservolea | [53,86] |
| L. plantarum/C. famata/C. guilliermondii | Bella di Cerignola | [64] |
| L. plantarum/S. cerevisiae | Leccino | [53,86] |
| L. plantarum/W. anomalus | Cellina di Nardò | [53,86] |
| L. plantarum/W. anomalus | Bella di Cerignola | [33,65] |
| L. plantarum/W. anomalus/L. pentosus | Bella di Cerignola | [33] |
| L. pentosus/P. membranifaciens | Conservolea | [52,53] |
| L. pentosus/C. boidinii | Manzanilla | [87] |
| L. mesenteroides/S. cerevisiae | Kalamata | [53,86] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table Olives More than a Fermented Food. Foods 2020, 9, 178. https://doi.org/10.3390/foods9020178
Perpetuini G, Prete R, Garcia-Gonzalez N, Khairul Alam M, Corsetti A. Table Olives More than a Fermented Food. Foods. 2020; 9(2):178. https://doi.org/10.3390/foods9020178
Chicago/Turabian StylePerpetuini, Giorgia, Roberta Prete, Natalia Garcia-Gonzalez, Mohammad Khairul Alam, and Aldo Corsetti. 2020. "Table Olives More than a Fermented Food" Foods 9, no. 2: 178. https://doi.org/10.3390/foods9020178
APA StylePerpetuini, G., Prete, R., Garcia-Gonzalez, N., Khairul Alam, M., & Corsetti, A. (2020). Table Olives More than a Fermented Food. Foods, 9(2), 178. https://doi.org/10.3390/foods9020178