Campylobacteriosis Agents in Meat Carcasses Collected from Two District Municipalities in the Eastern Cape Province, South Africa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Clearance
2.2. Study Area
2.3. Sample Collection
2.4. Microbiological Analysis of the Meat Samples
2.5. DNA Extraction
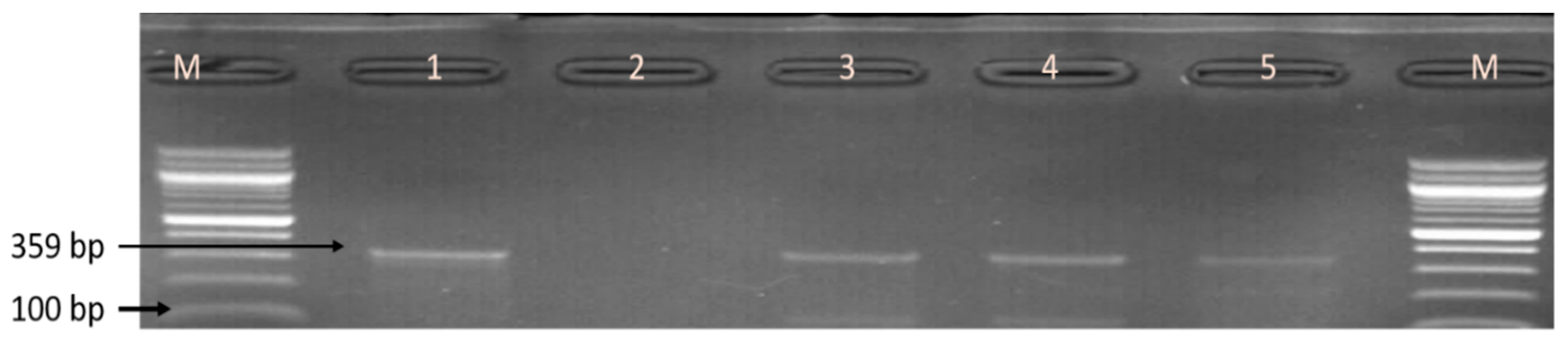
2.6. Molecular Identification of the Genus Campylobacter
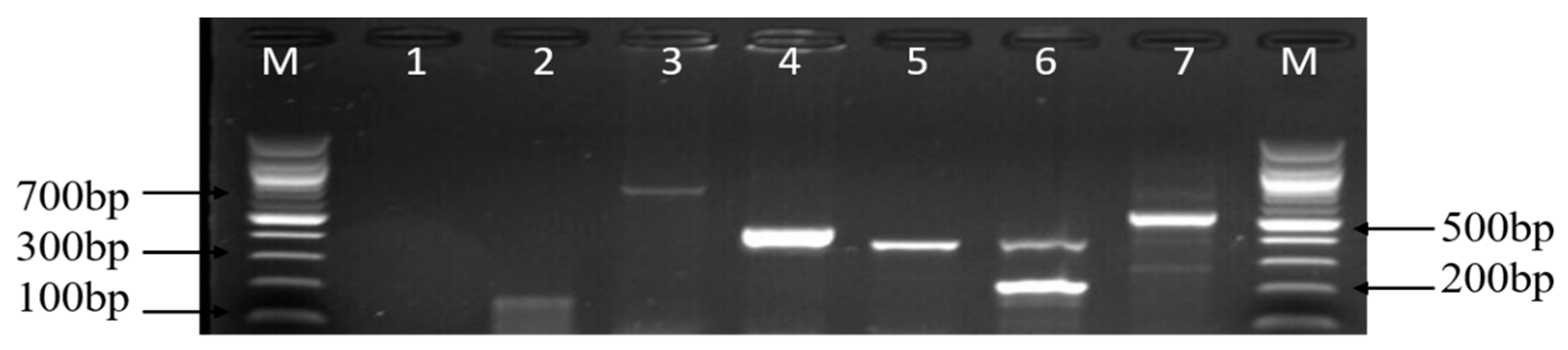
2.7. Molecular Classification of Campylobacter Species
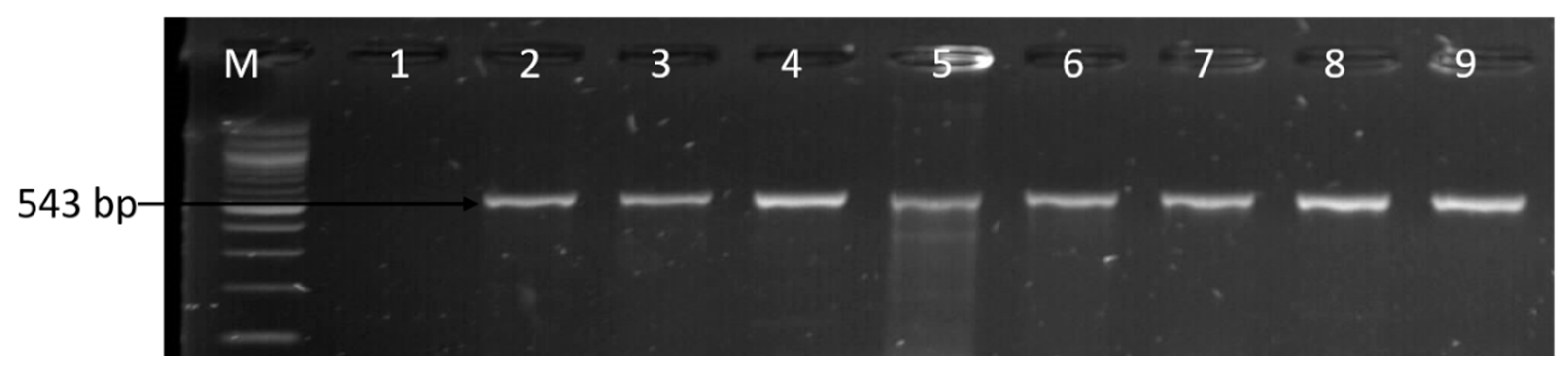
2.8. Molecular Detection of Virulence Genes
2.9. Phenotypic Determination of Antimicrobial Resistance
2.10. Multiple Antibiotic Resistance (MAR) Index
2.11. Genotypic Assessment of Antibiotic Resistance Genes
3. Results
3.1. Molecular confirmation of Campylobacter species
3.2. Molecular Characterisation of Campylobacter Species
3.3. Molecular Detection of Virulence Genes
3.4. Antibiotic Phenotypic Characteristics of the Identified Campylobacter Species
3.5. Assessment of Resistance Determinants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harvala, H.; Rosendal, T.; Lahti, E.; Engvall, E.O.; Brytting, M.; Wallensten, A.; Lindberg, A. Epidemiology of Campylobacter jejuni infections in Sweden, November 2011–October 2012: Is the severity of infection associated with C. jejuni sequence type? Infect. Ecol. Epidemiol. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Torrens, Y.; Miró, E.; Guirado, P.; Llovet, T.; Muñoz, C.; Cerdà-Cuéllar, M.; Madrid, C.; Balsalobre, C.; Navarro, F. Population structure, antimicrobial resistance, and virulence-associated genes in Campylobacter jejuni isolated from three ecological niches: Gastroenteritis patients, broilers, and wild birds. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisar, M.; Mushtaq, M.H.; Shehzad, W.; Hussain, A.; Nasar, M.; Nagaraja, K.V.; Goyal, S.M. Occurrence of Campylobacter in retail meat in Lahore, Pakistan. Acta Trop. 2018, 185, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.-H.; Sulaiman, I.M. Chapter 5—Campylobacteriosis: An Emerging Infectious Foodborne Disease. Foodborne Dis. 2018, 1, 119–155. [Google Scholar]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; De Silva, N.R.; Gargouri, N.; et al. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carron, M.; Chang, Y.M.; Momanyi, K.; Akoko, J.; Kiiru, J.; Bettridge, J.; Chaloner, G.; Rushton, J.; O’Brien, S.; Williams, N.; et al. Campylobacter, a zoonotic pathogen of global importance: Prevalence and risk factors in the fast-evolving chicken meat system of Nairobi, Kenya. PLoS Negl. Trop. Dis. 2018, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.P.; Chattopadhyay, U.K.; Naskar, K. Prevalence of’ Campylobacter species in raw meat samples sold in open markets of Kolkata city. Int. J. Agric. Environ. Biotechnol. 2016, 9, 535–539. [Google Scholar] [CrossRef]
- El-Aziz, A.D.; Abd-Allah, S.M.S. Incidence of Campylobacter species in wholesale chicken carcasses and chicken meat products in Assiut city, Egypt. Int. Food Res. J. 2017, 24, 2660–2665. [Google Scholar]
- Ndahi, M.D.; Kwaga, J.K.P.; Bello, M.; Kabir, J.; Umoh, V.J.; Yakubu, S.E.; Nok, A.J. Prevalence and antimicrobial susceptibility of Listeria monocytogenes and methicillin-resistant Staphylococcus aureus strains from raw meat and meat products in Zaria, Nigeria. Lett. Appl. Microbiol. 2013, 58, 262–269. [Google Scholar] [CrossRef]
- Luo, X.; Xu, X.; Chen, H.; Bai, R.; Zhang, Y.; Hou, X.; Zhang, F.; Zhang, Y.; Sharma, M.; Zeng, H.; et al. Food safety related knowledge, attitudes, and practices (KAP) among the students from nursing, education and medical college in Chongqing, China. Food Control 2019, 95, 181–188. [Google Scholar] [CrossRef]
- Guerra, M.M.M.; de Almeida, A.M.; Willingham, A.L. An overview of food safety and bacterial foodborne zoonoses in food production animals in the Caribbean region. Trop. Anim. Health Prod. 2016, 48, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, J.W.F.; Ab Mutalib, N.S.; Chan, K.G.; Lee, L.H. Rapid methods for the detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2015, 5, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newell, D.G.; Koopmans, M.; Verhoef, L.; Duizer, E.; Aidara-Kane, A.; Sprong, H.; Opsteegh, M.; Langelaar, M.; Threfall, J.; Scheutz, F.; et al. Food-borne diseases-the challenges of 20 years ago still persist while new ones continue to emerge. Int. J. Food Microbiol. 2010, 139, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization estimates of the global and regional disease burden of 11 foodborne parasitic diseases, 2010: A data synthesis. PLoS Med. 2015, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Camino, F.M.M.; Arisseto-Bragotto, A.P.; Block, J.M. Food quality, food-borne diseases, and food safety in the Brazilian food industry. Food Qual. Saf. 2017, 1, 13–27. [Google Scholar] [CrossRef]
- Epps, S.; Harvey, R.; Hume, M.; Phillips, T.; Anderson, R.; Nisbet, D. Foodborne Campylobacter: Infections, metabolism, pathogenesis and reservoirs. Int. J. Environ. Res. Public Health 2013, 10, 6292–6304. [Google Scholar] [CrossRef] [Green Version]
- Gourley, C.R.; Negretti, N.M.; Konkel, M.E. The food-borne pathogen Campylobacter jejuni depends on the AddAB DNA repair system to defend against bile in the intestinal environment. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, S.; He, M.; Zhang, Y.; Fu, Y.; Liang, H.; Jing, H.; Li, Y.; Ma, H.; Zhang, M. Prevalence and molecular characterization of Campylobacter spp. isolated from patients with diarrhea in Shunyi, Beijing. Front. Microbiol. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Wainwright, L.M.; Elvers, K.T.; Park, S.F.; Poole, R.K. A truncated haemoglobin implicated in oxygen metabolism by the microaerophilic food-borne pathogen Campylobacter jejuni. Microbiology 2005, 151, 4079–4091. [Google Scholar] [CrossRef] [Green Version]
- Reich, F.; Valero, A.; Schill, F.; Bungenstock, L.; Klein, G. Characterisation of Campylobacter contamination in broilers and assessment of microbiological criteria for the pathogen in broiler slaughterhouses. Food Control 2018, 87, 60–69. [Google Scholar] [CrossRef]
- Rahimi, E.; Alipoor-Amroabadi, M.; Khamesipour, F. Investigation of prevalence of thermotolerant Campylobacter spp. in livestock faeces. Can. J. Anim. Sci. 2017, 97, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Man, S.M. The clinical importance of emerging Campylobacter species. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Granić, K.; Krčar, D.; Uhitil, S.; Jakšić, S. Determination of Campylobacter spp. in poultry slaughterhouses and poultry meat. Vet. Arhiv. 2009, 79, 79–491. [Google Scholar]
- Modi, S.; Brahmbhatt, M.N.; Chatur, Y.A.; Nayak, J.B. Prevalence of Campylobacter species in milk and milk products, their virulence gene profile and anti-biogram. Vet. World 2015, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Platts-Mills, J.A.; Kosek, M. Update on the burden of Campylobacter in developing countries. Curr. Opin. Infect. Dis. 2014, 27, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Naravaneni, R.; Jamil, K. Rapid detection of food-borne pathogens by using molecular techniques. J. Med. Microbiol. 2005, 54, 51–54. [Google Scholar] [CrossRef]
- Katiyo, W.; de Kock, H.L.; Coorey, R.; Buys, E.M. Assessment of safety risks associated with handling chicken as based on practices and knowledge of a group of South African consumers. Food Control 2019, 101, 104–111. [Google Scholar] [CrossRef]
- NICD. Communicable Diseases Communique. Available online: http://www.nicd.ac.za/wp-content/uploads/2018/08 (accessed on 1 April 2018).
- Jahan, S. Epidemiology of Foodborne Illness. In Scientific, Health and Social Aspects of the Food Industry; Valdez, B., Ed.; IntechOpen: Qassim, Saudi Arabia, 2012; Volume 1, pp. 321–342. [Google Scholar]
- Magana, M.; Chatzipanagiotou, S.; Burriel, A.R.; Ioannidis, A. Inquiring into the Gaps of Campylobacter Surveillance Methods. Vet. Sci. 2017, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Kang, M. In vitro activity of fosfomycin against Campylobacter isolates from poultry and wild birds. PLoS ONE 2018, 13, 1–8. [Google Scholar] [CrossRef]
- Post, A.; Martiny, D.; van Waterschoot, N.; Hallin, M.; Maniewski, U.; Bottieau, E.; Van Esbroeck, M.; Vlieghe, E.; Ombelet, S.; Vandenberg, O.; et al. Antibiotic susceptibility profiles among Campylobacter isolates obtained from international travelers between 2007 and 2014. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2101–2107. [Google Scholar] [CrossRef] [Green Version]
- Zollner-Schwetz, I.; Krause, R. Therapy of acute gastroenteritis: Role of antibiotics. Clin. Microbiol. Infect. 2015, 21, 744–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepanska, B.; Andrzejewska, M.; Spica, D.; Klawe, J.J. Prevalence and antimicrobial resistance of Campylobacter jejuni and Campylobacter coli isolated from children and environmental sources in urban and suburban areas. BMC Microbiol. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashoma, I.P.; Mkomba, F.D.; Bunyaga, A.S. Prevalence and antimicrobial resistance in Campylobacter from different stages of the chicken meat supply chain in Morogoro, Tanzania. Microb. Drug Res. 2016, 22, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giannatale, E.; Di Serafino, G.; Zilli, K.; Alessiani, A.; Sacchini, L.; Garofolo, G.; Aprea, G.; Marotta, F. Characterization of antimicrobial resistance patterns and detection of virulence genes in Campylobacter isolates in Italy. Sensors 2014, 14, 3308–3322. [Google Scholar] [CrossRef] [Green Version]
- Habib, I.; Berkvens, D.; De Zutter, L.; Dierick, K.; Van Huffel, X.; Speybroeck, N.; Geeraerd, A.H.; Uyttendaele, M. Campylobacter contamination in broiler carcasses and correlation with slaughterhouses operational hygiene inspection. Food Microbiol. 2012, 29, 105–112. [Google Scholar] [CrossRef]
- ISO. Part 1: Detection method. UNI EN ISO Norm 10272-1:2006. In Microbiology of Food and Animal Feeding Stuffs-Horizontal Method for Detection and Enumeration of campylobacter spp.; International Organization for Standardization: Geneva, Switzerland, 2006. [Google Scholar]
- Sierra-Arguello, Y.M.; Furian, T.Q.; Perdoncini, G.; Moraes, H.L.; Salle, C.T.; Rodrigues, L.B.; dos Santos, L.R.; Gomes, M.J.P.; do Nascimento, V.P. Fluoroquinolone resistance in Campylobacter jejuni and Campylobacter coli from poultry and human samples assessed by PCR-restriction fragment length polymorphism assay. PLoS ONE 2018, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Moreno, Y.; Botella, S.; Luis Alonso, J.; Ferru’s, M.A.; Herna’ndez, M.; Herna’ndez, J. Specific detection of Arcobacter and Campylobacter strains in Water and Sewage by PCR and Fluorescent in Situ Hybridization. Appl. Environ. Microbiol. 2003, 69, 1181–1186. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki-Matsune, W.; Taguchi, M.; Seto, K.; Kawahara, R.; Kawatsu, K.; Kumeda, Y.; Kitazato, M.; Nukina, M.; Misawa, N.; Tsukamoto, T. Development of a multiplex PCR assay for identification of Campylobacter coli, Campylobacter fetus, Campylobacter hyointestinalis subsp. hyointestinalis, Campylobacter jejuni, Campylobacter lari and Campylobacter upsaliensis. J. Med. Microbiol. 2007, 56, 1467–1473. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.C.; Ruiz-Palacios, G.M.; Ramos-Cervantes, P.; Cervantes, L.E.; Jiang, X.; Pickering, L.K. Molecular characterization of invasive and noninvasive Campylobacter jejuni and Campylobacter coli isolates. J. Clin. Microbiol. 2001, 39, 1353–1359. [Google Scholar] [CrossRef] [Green Version]
- Casabonne, C.; Gonzalez, A.; Aquili, V.; Subils, T.; Balague, C. Prevalence of seven virulence genes of Campylobacter jejuni isolated from patients with diarrhea in Rosario, Argentina. Intern. J. Infect. 2016, 3, 1–6. [Google Scholar] [CrossRef]
- Wilson, D.L.; Rathinam, V.A.; Qi, W.; Wick, L.M.; Landgraf, J.; Bell, J.A.; Plovanich-Jones, A.; Parrish, J.; Finley, R.L.; Mansfield, L.S.; et al. Genetic diversity in Campylobacter jejuni is associated with differential colonization of broiler chickens and C57BL/6J IL10-deficient mice. Microbiology 2010, 156, 2046–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Method for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; CLSI: Wayne, NJ, USA, 2015. [Google Scholar]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes 2001, 15, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.S.; Fox, M.L.; Holland, S.M.; Stock, F.; Gill, V.J.; Fedorko, D.P. Resistance to multiple fluoroquinolones in a clinical isolate of Streptococcus pyogenes: Identification of gyrA and parC and specification of point mutations associated with resistance. Antimicrob. Agents Chemother. 2000, 44, 196–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osode, A.N.; Okoh, A.I. Impact of discharged wastewater final effluent on the physicochemical qualities of a receiving watershed in a suburban community of the Eastern Cape Province. Clean Soil Air Water 2009, 37, 938–944. [Google Scholar] [CrossRef]
- Maynard, C.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Lariviere, S.; Harel, J. Heterogeneity among virulence and antimicrobial resistance gene profiles of extraintestinal Escherichia coli isolates of animal and human origin. J. Clin. Microbiol. 2004, 42, 5444–5452. [Google Scholar] [CrossRef] [Green Version]
- Velusamy, S.; Barbara, E.G.; Mark, J.L.; Lien, T.N.; Susan, I.H.; Ynte, H.S.; Stephen, P.O. Phenotypic and genotypic antimicrobial resistance patterns of Escherichia coli isolated from dairy cows with mastitis. Vet. Microbiol. 2007, 124, 319–328. [Google Scholar]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Stella, S.; Soncini, G.; Ziino, G.; Panebianco, A.; Pedonese, F.; Nuvoloni, R.; Di Giannatale, E.; Colavita, G.; Alberghini, L.; Giaccone, V. Prevalence and quantification of thermophilic Campylobacter spp. in Italian retail poultry meat: Analysis of influencing factors. Food Microbiol. 2007, 62, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Sinulingga, T.S.; Aziz, S.A.; Bitrus, A.A.; Zunita, Z.; Abu, J. Occurrence of Campylobacter species from broiler chickens and chicken meat in Malaysia. Trop. Anim. Health Prod. 2019, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dong, Y.; Deng, F.; Liu, D.; Yao, H.; Zhang, Q.; Shen, J.; Liu, Z.; Gao, Y.; Wu, C.; et al. Species shift and multidrug resistance of Campylobacter from chicken and swine, China, 2008–2014. J. Antimicrob. Chemother. 2015, 71, 666–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyard-Nicodème, M.; Rivoal, K.; Houard, E.; Rose, V.; Quesne, S.; Mourand, G.; Rouxel, S.; Kempf, I.; Guillier, L.; Gauchard, F.; et al. Prevalence and characterization of Campylobacter jejuni from chicken meat sold in French retail outlets. Int. J. Food Microbiol. 2015, 203, 8–14. [Google Scholar] [CrossRef]
- Chon, J.W.; Lee, S.K.; Yoon, Y.; Yoon, K.S.; Kwak, H.S.; Joo, I.S.; Seo, K.H. Quantitative prevalence and characterization of Campylobacter from chicken and duck carcasses from poultry slaughterhouses in South Korea. Poult. Sci. 2018, 97, 2909–2916. [Google Scholar] [CrossRef]
- García-Sánchez, L.; Melero, B.; Diez, A.M.; Jaime, I.; Rovira, J. Characterization of Campylobacter species in Spanish retail from different fresh chicken products and their antimicrobial resistance. Food Microbiol. 2018, 76, 457–465. [Google Scholar] [CrossRef]
- Zou, C.; Zhang, H.; Yang, X.; Tang, H.; Xu, X.; Ren, F.; Huang, J. Qualitative and quantitative epidemiological analysis of Campylobacter in chicken sold in Yangzhou. J. Food Saf. Qual. 2018, 9, 5040–5043. [Google Scholar]
- Andrzejewska, M.; Klawe, J.; Szczepańska, B.; Śpica, D. Occurrence of virulence genes among Campylobacter jejuni and Campylobacter coli isolates from domestic animals and children. Pol. J. Vet. Sci. 2011, 4, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Seliwiorstow, T.; Baré, J.; Berkvens, D.; Van Damme, I.; Uyttendaele, M.; De Zutter, L. Identification of risk factors for Campylobacter contamination levels on broiler carcasses during the slaughter process. Int. J. Food Microbiol. 2016, 226, 26–32. [Google Scholar] [CrossRef]
- Maktabi, S.; Ghorbanpoor, M.; Hossaini, M.; Motavalibashi, A. Detection of multi-antibiotic resistant Campylobacter coli and Campylobacter jejuni in beef, mutton, chicken and water buffalo meat in Ahvaz, Iran. Vet. Res. Forum 2019, 10, 37–42. [Google Scholar]
- Kashoma, I.P.; Kassem, I.I.; John, J.; Kessy, B.M.; Gebreyes, W.; Kazwala, R.R.; Rajashekara, G. Prevalence and antimicrobial resistance of Campylobacter isolated from dressed beef carcasses and raw milk in Tanzania. Microb. Drug Resist. 2016, 2, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premarathne, J.M.; Anuar, A.S.; Thung, T.Y.; Satharasinghe, D.A.; Jambari, N.N.; Abdul-Mutalib, N.A.; Huat, J.T.Y.; Basri, D.F.; Rukayadi, Y.; Nakaguchi, Y.; et al. Prevalence and antibiotic resistance against tetracycline in Campylobacter jejuni and C. coli in cattle and beef meat from Selangor, Malaysia. Front. Microbiol. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Noormohamed, A.; Fakhr, M.K. Prevalence and antimicrobial susceptibility of Campylobacter species in Oklahoma conventional and organic retail poultry. Open Microbiol. J. 2014, 8, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szosland-Fałtyn, A.N.N.A.; Bartodziejska, B.; Krolasik, J.; Paziak-Domańska, B.E.A.T.A.; Korsak, D.; Chmiela, M. The Prevalence of Campylobacter spp. Pol. J. Microbiol. 2018, 67, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Hodges, L.M.; Carrillo, C.D.; Upham, J.P.; Borza, A.; Eisebraun, M.; Kenwell, R.; Mutschall, S.K.; Haldane, D.; Schleihauf, E.; Taboada, E.N. A strain comparison of Campylobacter isolated from retail poultry and human clinical cases in Atlantic Canada. PLoS ONE 2019, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ocejo, M.; Oporto, B.; Hurtado, A. Occurrence of Campylobacter jejuni and Campylobacter coli in Cattle and Sheep in Northern Spain and Changes in Antimicrobial Resistance in Two Studies 10-years Apart. Pathogens 2019, 8, 98. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, I.M.; Hsieh, Y.H.; Simpson, S. Species identification of Campylobacter jejuni and Campylobacter coli isolates from raw poultry products by MALDI–TOF MS and rRNA sequence analysis. J. AOAC Int. 2019, 103, 197–204. [Google Scholar] [CrossRef]
- Vinueza-Burgos, C.; Wautier, M.; Martiny, D.; Cisneros, M.; Van Damme, I.; De Zutter, L. Prevalence, antimicrobial resistance and genetic diversity of Campylobacter coli and Campylobacter jejuni in Ecuadorian broilers at slaughter age. Poult. Sci. 2017, 96, 2366–2374. [Google Scholar] [CrossRef]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a foodborne pathogen: A review. Front. Microbiol. 2011, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chukwu, M.O.; Luther King Abia, A.; Ubomba-Jaswa, E.; Obi, L.; Dewar, J.B. Characterization and Phylogenetic Analysis of Campylobacter Species Isolated from Paediatric Stool and Water Samples in the Northwest Province, South Africa. Int. J. Environ. Res. Public Health 2019, 16, 2205. [Google Scholar] [CrossRef] [Green Version]
- Abu-Madi, M.; Behnke, J.M.; Sharma, A.; Bearden, R.; Al-Banna, N. Prevalence of virulence/stress genes in Campylobacter jejuni from chicken meat sold in Qatari retail outlets. PLoS ONE 2016, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Redondo, N.; Carroll, A.; McNamara, E. Molecular characterization of Campylobacter causing human clinical infection using whole-genome sequencing: Virulence, antimicrobial resistance and phylogeny in Ireland. PLoS ONE 2019, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrzejewska, M.; Szczepańska, B.; Śpica, D.; Klawe, J.J. Trends in the occurrence and characteristics of Campylobacter jejuni and Campylobacter coli isolates from poultry meat in Northern Poland. Food Control 2015, 51, 190–194. [Google Scholar] [CrossRef]
- Ripabelli, G.; Tamburro, M.; Minelli, F.; Leone, A.; Sammarco, M.L. Prevalence of virulence-associated genes and cytolethal distending toxin production in Campylobacter spp. isolated in Italy. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.T.; Nalevaiko, P.C.; Mendonça, E.P.; Borges, L.W.; Fonseca, B.B.; Beletti, M.E.; Rossi, D.A. Campylobacter jejuni strains isolated from chicken meat harbour several virulence factors and represent a potential risk to humans. Food Control 2013, 33, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Melo, R.T.; Grazziotin, A.L.; Júnior, E.C.V.; Prado, R.R.; Mendonça, E.P.; Monteiro, G.P.; Peres, P.A.; Rossi, D.A. Evolution of Campylobacter jejuni of poultry origin in Brazil. Food Microbiol. 2019, 82, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Wu, Q.; Zhang, J.; Shen, S. Prevalence, genetic diversity and antimicrobial susceptibility of Campylobacter jejuni isolated from retail food in China. Food Control 2016, 62, 10–15. [Google Scholar] [CrossRef]
- Wieczorek, K.; Denis, E.; Lynch, O.; Osek, J. Molecular characterization and antibiotic resistance profiling of Campylobacter isolated from cattle in Polish slaughterhouses. Food Microbiol. 2013, 34, 130–136. [Google Scholar] [CrossRef]
- Reddy, S.; Zishiri, O.T. Genetic characterisation of virulence genes associated with adherence, invasion and cytotoxicity in Campylobacter spp. isolated from commercial chickens and human clinical cases. Onderstepoort. J. Vet. Res. 2018, 85, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.K.; Moon, D.C.; Chae, M.H.; Kim, H.J.; Nam, H.M.; Kim, S.R.; Jang, G.C.; Lee, K.; Jung, S.C.; Lee, H.S. Macrolide resistance mechanisms and virulence factors in erythromycin-resistant Campylobacter species isolated from chicken and swine feces and carcasses. J. Vet. Med. Sci. 2016, 72, 1791–1795. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Luo, Q.; Chen, Y.; Li, T.; Wen, G.; Zhang, R.; Luo, L.; Lu, Q.; Ai, D.; Wang, H.; et al. Molecular epidemiology, virulence determinants and antimicrobial resistance of Campylobacter spreading in retail chicken meat in Central China. Gut Pathogens 2016, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Guan, X.; Zeng, H.; Li, J.; Huang, X.; Wen, Y.; Zhao, Q.; Huang, X.; Yan, Q.; Huang, Y.; et al. Prevalence, antimicrobial resistance profiles and virulence-associated genes of thermophilic Campylobacter spp. isolated from ducks in a Chinese slaughterhouse. Food Control 2019, 104, 157–166. [Google Scholar] [CrossRef]
- Miranda, K.L.; Lage, A.P. Antimicrobial susceptibility of Campylobacter sp strains isolated from calves with and without diarrhea in Minas Gerais state, Brazil. Braz. J. Microbiol. 2007, 38, 357–362. [Google Scholar] [CrossRef]
- Kuana, S.L.; Santos, L.R.; Rodrigues, L.B.; Borsoi, A.; Moraes, H.L.S.; Salle, C.T.P.; Nascimento, V.P. Occurrence and characterization of Campylobacter in the Brazilian production and processing of broilers. Avian Dis. 2008, 52, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, M. A Cosmopolitan One Health Issue: Campylobacteriosis. Clin. Microbiol. 2018, 7, 1–5. [Google Scholar] [CrossRef]
- Davis, R.; Brown, P.D. Multiple antibiotic resistance index, fitness and virulence potential in respiratory Pseudomonas aeruginosa from Jamaica. J. Med. Microbiol. 2016, 65, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Wołkowicz, T.; Osek, J. Antimicrobial resistance and virulence-associated traits of Campylobacter jejuni isolated from poultry food chain and humans with diarrhea. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Nguyen, T.N.M.; Hotzel, H.; Njeru, J.; Mwituria, J.; El-Adawy, H.; Tomaso, H.; Neubauer, H.; Hafez, H.M. Antimicrobial resistance of Campylobacter isolates from small scale and backyard chicken in Kenya. Gut Pathogens 2016, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Ye, Y.; Liu, Y.; Wang, A.; Li, Y.; Zhou, X.; Pan, H.; Zhang, J.; Xu, X. Molecular identification of multidrug-resistant Campylobacter species from diarrheal patients and poultry meat in Shanghai, China. Front. Microbiol. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Rahimi, E.; Ameri, M.; Kazemeini, H.R. Prevalence and antimicrobial resistance of Campylobacter species isolated from raw camel, beef, lamb, and goat meat in Iran. Foodborne Path. Dis. 2010, 7, 443–447. [Google Scholar] [CrossRef]
- Wieczorek, K.; Osek, J. Antimicrobial resistance mechanisms among Campylobacter. BioMed Res. Int. 2013, 1–12. [Google Scholar]
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A One Health perspective. In Antimicrobial Resistance in Bacteria from Livestock and Companion Animals; ASM Press: Washington, DC, USA, 2018; pp. 521–547. [Google Scholar]
- Garcia, S.N.; Osburn, B.I.; Cullor, J.S. A one health perspective on dairy production and dairy food safety. One Health 2019, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Hughes, J.M. Critical importance of a one health approach to antimicrobial resistance. EcoHealth 2019, 16, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meat Types | No. of Samples | No. of Presumptive Campylobacter Isolates | No. of Isolates Identified as Genus Campylobacter |
|---|---|---|---|
| Turkey | 11 | 16 | 5 (31.25%) |
| Pork | 35 | 131 | 33 (25.19%) |
| Mutton | 22 | 9 | 4 (44.44%) |
| Mutton offals (heart) | 2 | 6 | 0 |
| Beef | 27 | 89 | 30 (33.71%) |
| Beef offals (intestine, kidney, and liver) | 31 | 126 | 39 (30.95%) |
| Chicken | 68 | 300 | 81 (27%) |
| Chicken offals (liver, gizzard, and heart) | 52 | 165 | 48 (29.09%) |
| Meat Typologies | C. jejuni | C. coli | C. lari | C. fetus | No. of Campylobacter Species Detected in the Meat Types |
|---|---|---|---|---|---|
| Pork | 4 | 9 | 0 | 3 | 16 |
| Beef | 3 | 0 | 0 | 2 | 5 |
| Beef offals | 10 | 15 | 0 | 1 | 26 |
| Chicken | 9 | 16 | 0 | 2 | 27 |
| Chicken offals | 14 | 5 | 0 | 1 | 20 |
| Mutton | 0 | 0 | 0 | 0 | 0 |
| Mutton offals | 0 | 3 | 0 | 0 | 3 |
| Turkey | 0 | 5 | 0 | 0 | 5 |
| Total | 40 | 53 | 0 | 9 | 102 |
| Virulence Genes Screened | Campylobacter Species | ||
|---|---|---|---|
| C. jejuni (%) | C. coli (%) | C. fetus (%) | |
| iam | 7 (6.86) | 35 (34.31) | 2 (1.96) |
| cadF | 4 (3.92) | 34 (33.33) | 0 |
| flgR | 11 (10.78) | 8 (7.84) | 0 |
| cdtB | 6 (5.88) | 17 (16.67) | 1(0.98) |
| flaA | 0 | 2 (1.96) | 0 |
| ciaB | 0 | 0 | 0 |
| Pattern of Multiple Virulence Genes | Number of Campylobacter Species | Total Number | ||||
|---|---|---|---|---|---|---|
| C. coli | C. fetus | C. jejuni | - | |||
| 1 | iam, cadF | 16 | - | 1 | - | 17 |
| 2 | iam, flgR | 2 | - | - | - | 2 |
| 3 | iam, cdtB | 1 | - | - | - | 1 |
| 4 | cadF, flgR | - | - | 1 | - | 1 |
| 5 | cadF, cdtB | 4 | - | - | - | 4 |
| 6 | flgR, cdtB | 1 | - | - | - | 1 |
| 7 | iam, cadF, flaA | 1 | - | - | - | 1 |
| 8 | iam, cadF, cdtB | 7 | - | - | - | 7 |
| 9 | cadF, cdtB, flgR | 1 | - | - | - | 1 |
| 10 | iam, cadF, flgR, cdtB | 2 | - | 1 | - | 3 |
| No | Multiple Antimicrobial Resistance Profile | No of Isolates | Total | MAR Index | ||
|---|---|---|---|---|---|---|
| C. coli | C. jejuni | C. fetus | ||||
| 1 | C-CD-AP | 1 | - | - | 1 | 0.25 |
| 2 | C-E-ATH-CD-AP | 1 | - | - | 1 | 0.42 |
| 3 | CRO-C-E-CD-AP | - | - | 1 | 1 | 0.42 |
| 4 | E-ATH-CD-T-DXT-AP | - | 1 | - | 1 | 0.5 |
| 5 | LEV-CRO-CIP-ATH-CD-AP | - | - | 1 | 1 | 0.5 |
| 6 | CRO-C-CIP-E-ATH-CD-AP | - | 1 | - | 1 | 0.58 |
| 7 | CRO-E-ATH-CD-T-DXT-AP | - | 1 | - | 1 | 0.58 |
| 8 | CRO-C-CIP-E-ATH-CD-AP | - | 1 | - | 1 | 0.58 |
| 9 | C-E-ATH-CD-T-DXT-AP | 1 | - | - | 1 | 0.58 |
| 10 | LEV-CRO-C-CIP-E-ATH-CD-AP | 1 | - | - | 1 | 0.67 |
| 11 | CRO-C-E-ATH-T-GM-DXT-AP | 1 | - | - | 1 | 0.67 |
| 12 | CRO-C-E-ATH-CD-T-DXT-AP | 1 | 1 | - | 2 | 0.67 |
| 13 | CRO-CIP-E-ATH-CD-T-DXT-AP | 1 | 1 | - | 2 | 0.67 |
| 14 | CRO-E-ATH-CD-T-GM-DXT-AP | - | 1 | - | 1 | 0.75 |
| 15 | CRO-C-CIP-E-ATH-CD-T-DXT-AP | - | - | 1 | 1 | 0.75 |
| 16 | CRO-CIP-E-ATH-IMI-CD-T-DXT-AP | - | - | 1 | 1 | 0.75 |
| 17 | LEV-CRO-CIP-E-ATH-CD-T-DXT-AP | - | 1 | - | 1 | 0.75 |
| 18 | CRO-C-CIP-ATH-CD-T-GM-DXT-AP | 1 | - | - | 1 | 0.75 |
| 19 | LEV-CRO-C-CIP-E-ATH-CD-T-DXT | 1 | - | - | 1 | 0.75 |
| 20 | CRO-C-E-ATH-CD-T-GM-DXT-AP | 2 | 1 | 3 | 0.75 | |
| 21 | LEV-CRO-C-CIP-E-ATH-CD-T-AP | - | - | 1 | 1 | 0.75 |
| 41 | LEV-CRO-C-CIP-E-ATH-CD-GM-DXT-AP | - | 1 | - | 1 | 0.83 |
| 22 | LEV-CRO-C-CIP-E-ATH-CD-T-DXT-AP | 6 | 8 | - | 14 | 0.83 |
| 23 | LEV-CRO-CIP-E-ATH-CD-T-GM-DXT-AP | 1 | - | - | 1 | 0.83 |
| 24 | CRO-C-CIP-E-ATH-CD-T-GM-DXT-AP | - | 1 | - | 1 | 0.83 |
| 25 | LEV-CRO-CIP-E-ATH-CD-T-GM-DXT-AP | - | 1 | - | 1 | 0.83 |
| 26 | LEV-CRO-C-CIP-E-ATH-CD-T-GM-DXT-AP | 8 | 8 | - | 16 | 0.92 |
| 27 | CRO-C-CIP-E-ATH-IMI-CD-T-GM-DXT-AP | - | 1 | - | 1 | 0.92 |
| 28 | LEV-CRO-C-CIP-E-ATH-IMI-CD-GM-DXT-AP | - | 1 | - | 1 | 0.92 |
| 29 | LEV-CRO-C-CIP-E-ATH-IMI-CD-T-GM-DXT-AP | 14 | 3 | - | 17 | 1 |
| No | Distribution Pattern of Antibiotic Resistance Determinants | No of Isolates | Total | ||
|---|---|---|---|---|---|
| C. coli | C. jejuni | C. fetus | |||
| 1 | tetA, catII | 1 | - | 3 | 4 |
| 2 | tetM, catII | 2 | - | - | 2 |
| 3 | ampC, catII | 1 | - | - | 1 |
| 4 | catII, ermB | 1 | - | - | 1 |
| 5 | tetA, ampC | - | - | 1 | 1 |
| 6 | tetA, catII, gyra | - | - | 1 | 1 |
| 7 | tetA, tetB, ampC | - | 1 | - | 1 |
| 8 | tetA, ampC, catII | 2 | 2 | - | 4 |
| 9 | tetA, catII, ermB | 1 | - | - | 1 |
| 10 | tetM, ampC, catII | 1 | - | - | 1 |
| 11 | tetA, tetK, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 12 | ampC, catII, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 13 | tetM, catII, aac(3)-IIa (aacC2)a | 2 | - | - | 2 |
| 14 | tetA, ampC, catII, gyrA | 2 | 1 | 1 | 4 |
| 15 | tetA, ampC, catII, ermB | - | 2 | - | 2 |
| 16 | tetA, tetM, catII, ermB | 2 | - | - | 2 |
| 17 | tetA, tetB, ampC, gyrA | - | 1 | - | 1 |
| 18 | tetA, tetB, ampC, catII | 1 | 1 | - | 2 |
| 19 | tetA, tetM, tetK, catII | - | 1 | - | 1 |
| 20 | tetA, catII, ermB, gyrA | - | 1 | - | 1 |
| 21 | tetA, tetB, catII, ermB | 1 | - | - | 1 |
| 22 | tetA, tetM, ampC, catII | 1 | - | - | 1 |
| 23 | tetA, tetM, ampC, gyrA | 1 | - | - | 1 |
| 24 | tetM, ampC, catII, gyrA | 1 | - | - | 1 |
| 25 | tetM, catII, gyrA, aac(3)-IIa (aacC2)a | 1 | - | - | 1 |
| 26 | tetM, ampC, gyrA, aac(3)-IIa (aacC2)a | 1 | - | - | 1 |
| 27 | tetA, ampC, catII, aac(3)-IIa (aacC2)a | 1 | 3 | - | 4 |
| 28 | tetA, tetM, ampC, catII, gyrA | 3 | 1 | - | 4 |
| 29 | tetA, tetB, tetM, catII, gyrA | 1 | - | - | 1 |
| 30 | tetA, tetB, ampC, catII, gyrA | - | 3 | - | 3 |
| 31 | tetA, ampC, catII, ermB, gyrA | - | 2 | - | 2 |
| 32 | tetA, tetM, ampC, catII, aac(3)-IIa (aacC2)a | 1 | - | - | 1 |
| 33 | tetA, tetM, catII, gyrA, aac(3)-IIa (aacC2)a | 1 | - | - | 1 |
| 34 | tetM, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | 1 | - | - | 1 |
| 35 | tetA, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | 1 | 2 | - | 3 |
| 36 | tetA, tetM, ampC, catII, ermB, gyrA | 2 | 1 | - | 3 |
| 37 | tetA, tetB, tetM, ampC, catII, ermB | 1 | - | - | 1 |
| 38 | tetA, tetM, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 39 | tetA, tetB, tetK, ampC, ermB, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 40 | tetA, tetB, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 41 | tetA, tetM, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | 2 | - | - | 2 |
| 42 | tetA, tetM, ampC, ermB, gyrA, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 42 | tetA, tetM, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | 1 | 1 | - | 2 |
| 43 | tetA, tetM, ampC, catII, ermB, gyrA, aac(3)-IIa (aacC2)a | - | 1 | - | 1 |
| 44 | tetA, tetB, tetM, ampC, catII, gyrA, aac(3)-IIa (aacC2)a | 1 | - | - | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Igwaran, A.; I. Okoh, A. Campylobacteriosis Agents in Meat Carcasses Collected from Two District Municipalities in the Eastern Cape Province, South Africa. Foods 2020, 9, 203. https://doi.org/10.3390/foods9020203
Igwaran A, I. Okoh A. Campylobacteriosis Agents in Meat Carcasses Collected from Two District Municipalities in the Eastern Cape Province, South Africa. Foods. 2020; 9(2):203. https://doi.org/10.3390/foods9020203
Chicago/Turabian StyleIgwaran, Aboi, and Anthony I. Okoh. 2020. "Campylobacteriosis Agents in Meat Carcasses Collected from Two District Municipalities in the Eastern Cape Province, South Africa" Foods 9, no. 2: 203. https://doi.org/10.3390/foods9020203
APA StyleIgwaran, A., & I. Okoh, A. (2020). Campylobacteriosis Agents in Meat Carcasses Collected from Two District Municipalities in the Eastern Cape Province, South Africa. Foods, 9(2), 203. https://doi.org/10.3390/foods9020203