Exogenous Spermidine Promotes γ-Aminobutyric Acid Accumulation and Alleviates the Negative Effect of NaCl Stress in Germinating Soybean (Glycine max L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Material Treatment and Experimental Design
2.2.1. Effects of Exogenous Spd on Physiology and Biochemistry of Germinating Soybeans under NaCl Stress
2.2.2. Effect of Exogenous Spd on GABA Metabolism of Germinating Soybeans under NaCl Stress
2.3. Measurement of Main Physiological and Biochemical Indexes
2.4. Measurement of Antioxidant Activities
2.5. Measurements of GABA Contents
2.6. GAD, DAO, PAO, and AMADH Activity Assay
2.7. GABA Synthase Gene Expression (Quantitative Real-Time PCR).
2.8. Statistical Analyses
3. Results
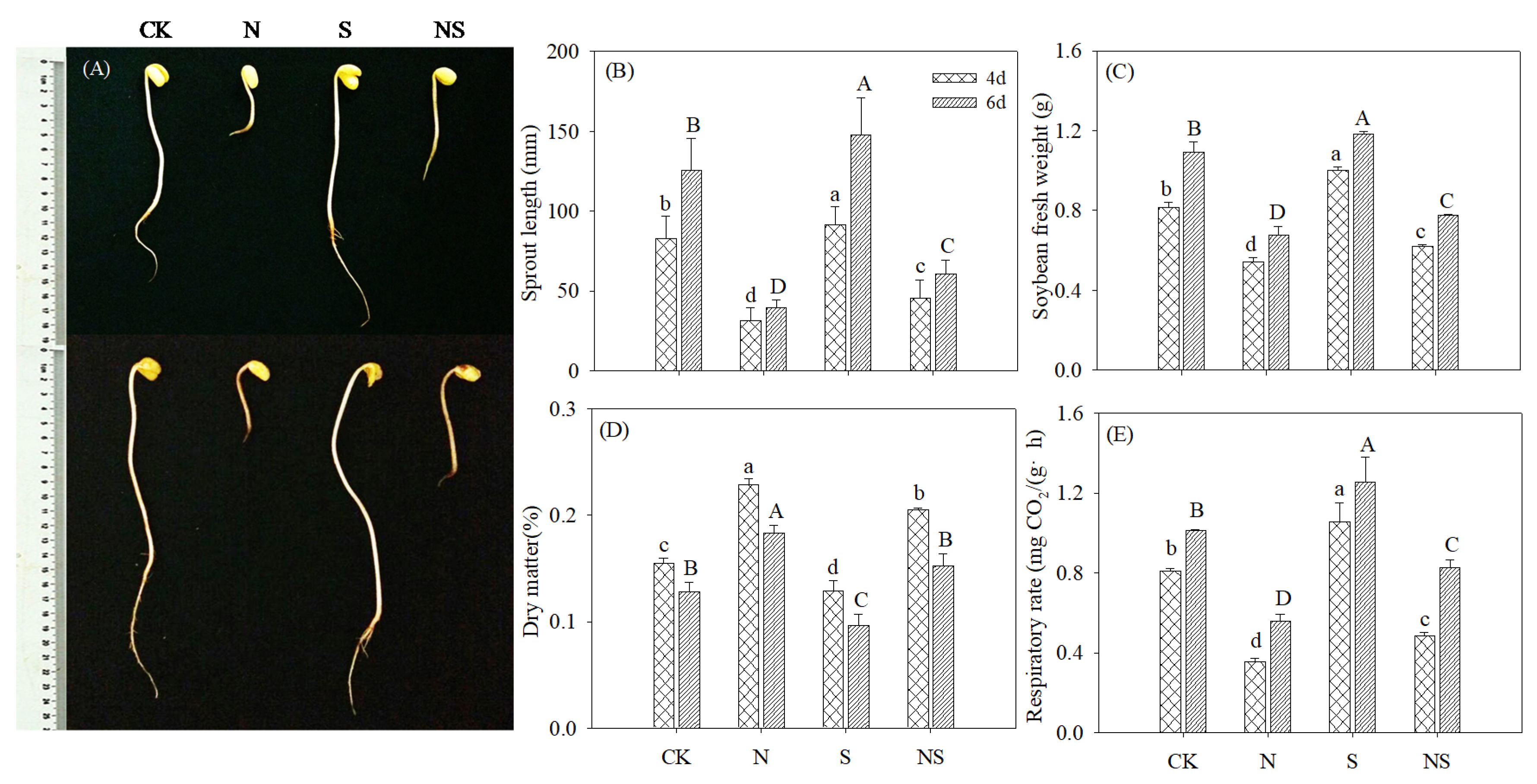
3.1. Growth Characteristics of Germinating Soybeans
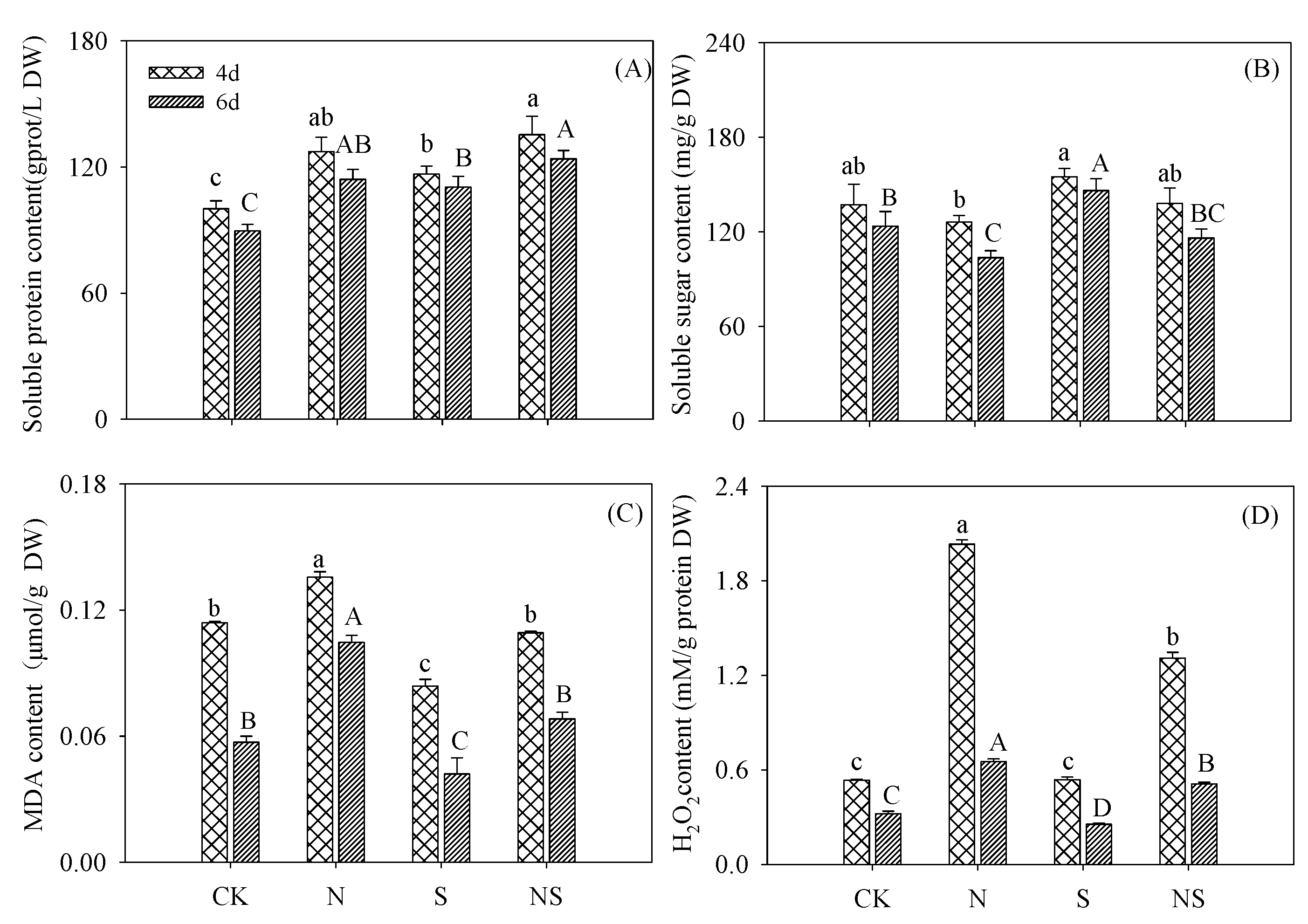
3.2. Soluble Protein, Soluble Sugar, MDA, and H2O2 Contents in Germinating Soybeans
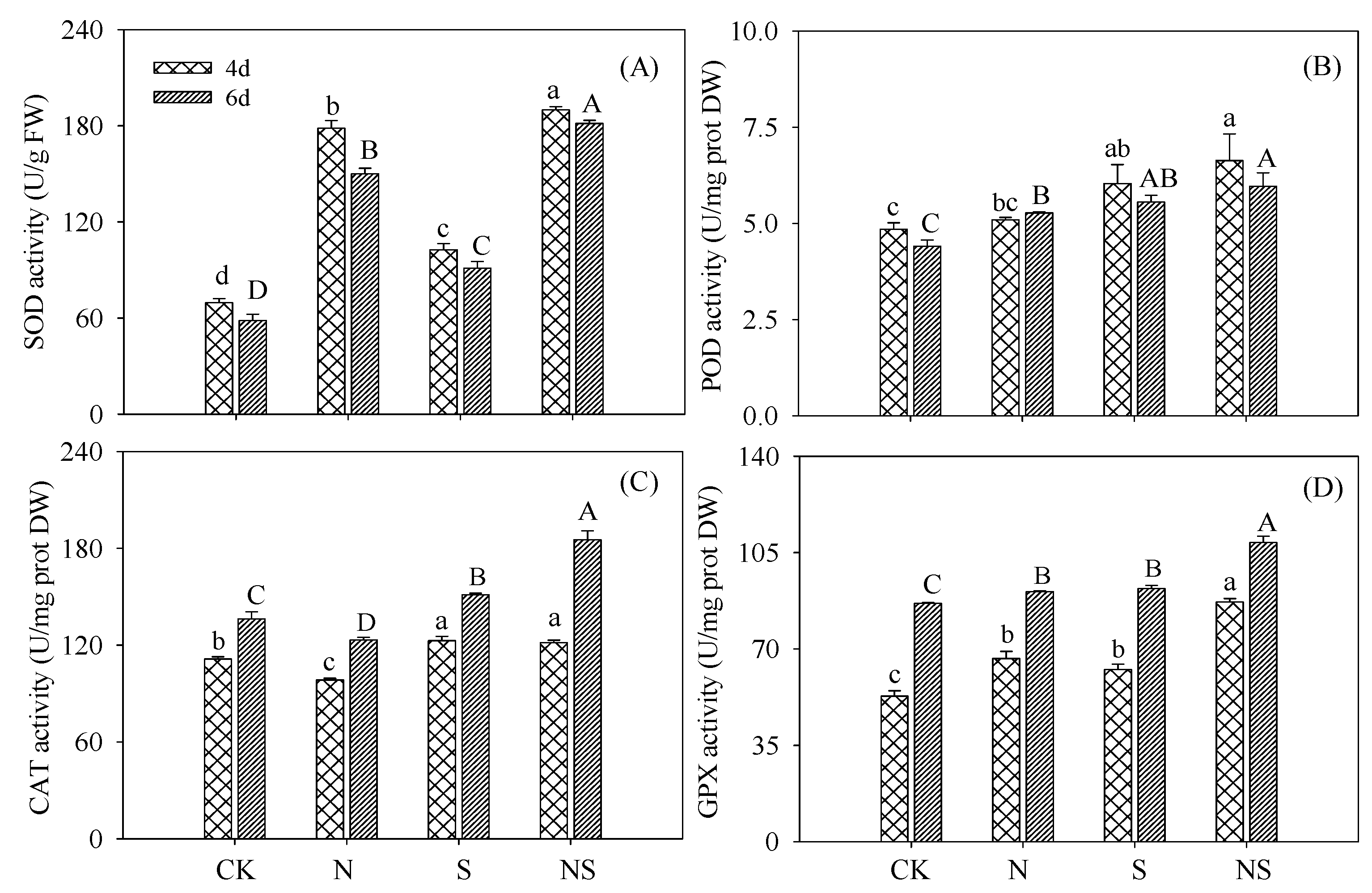
3.3. Antioxidant Enzyme Activity
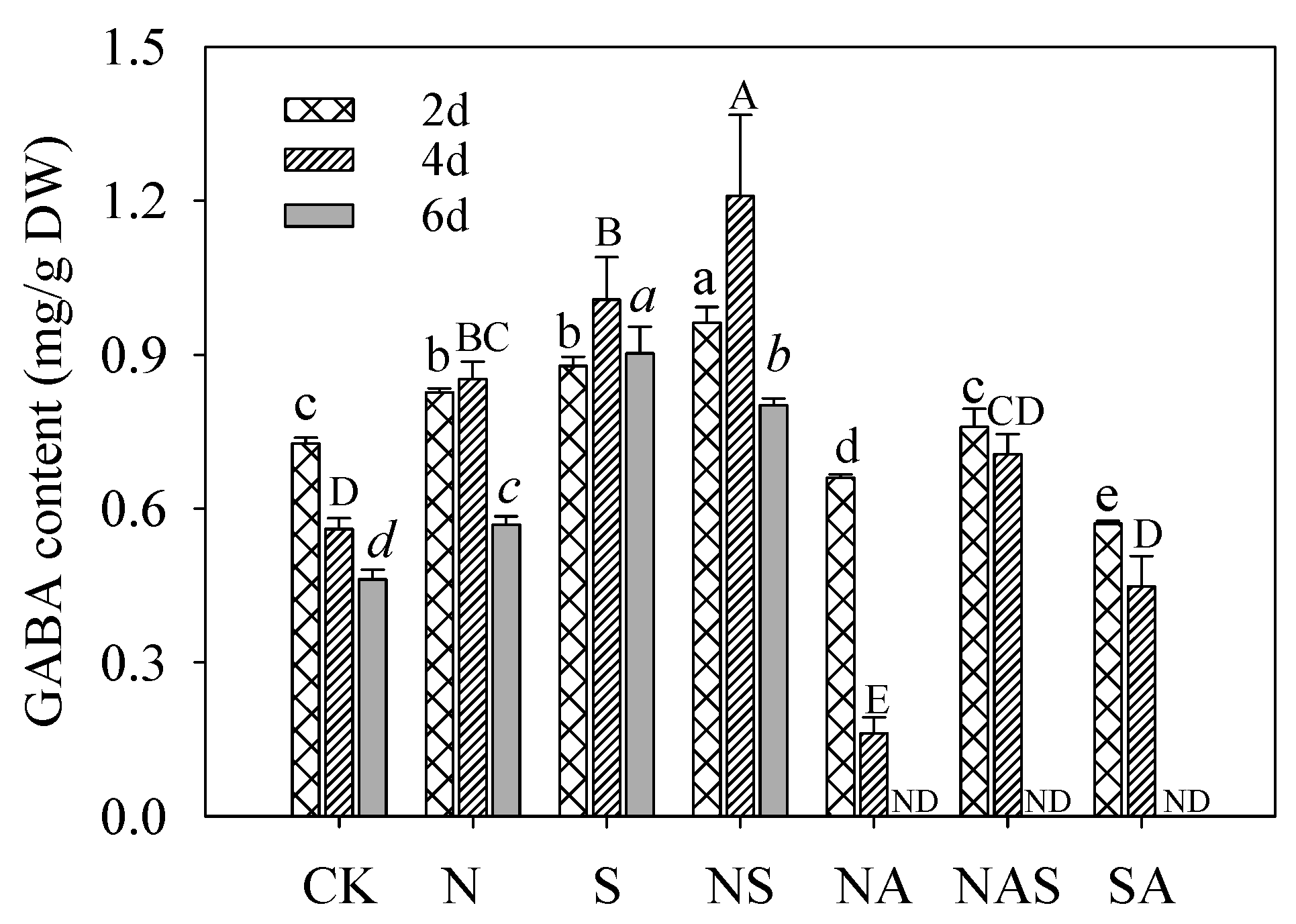
3.4. GABA Content in Germinating Soybeans
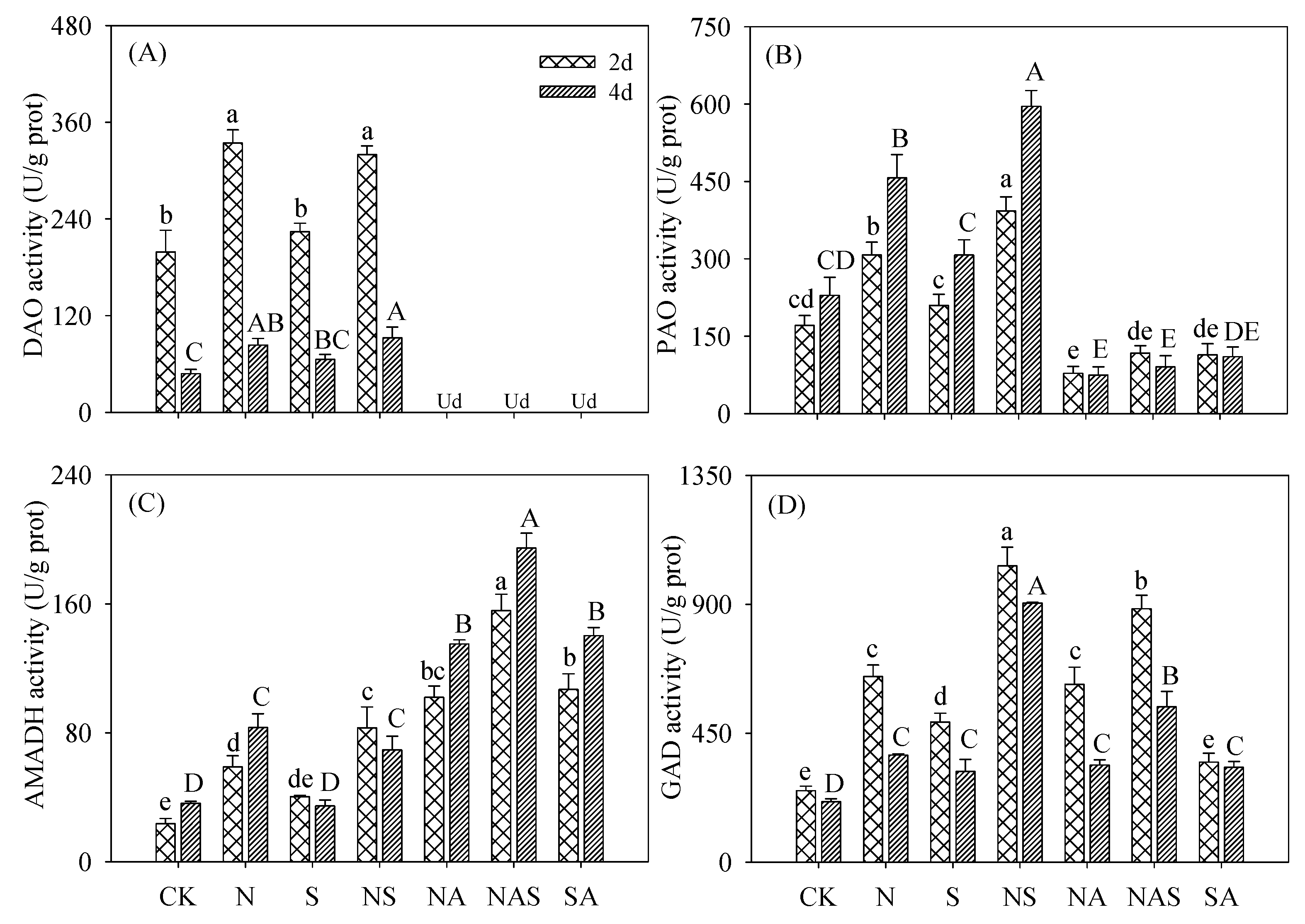
3.5. Changes in GABA Synthase Activity
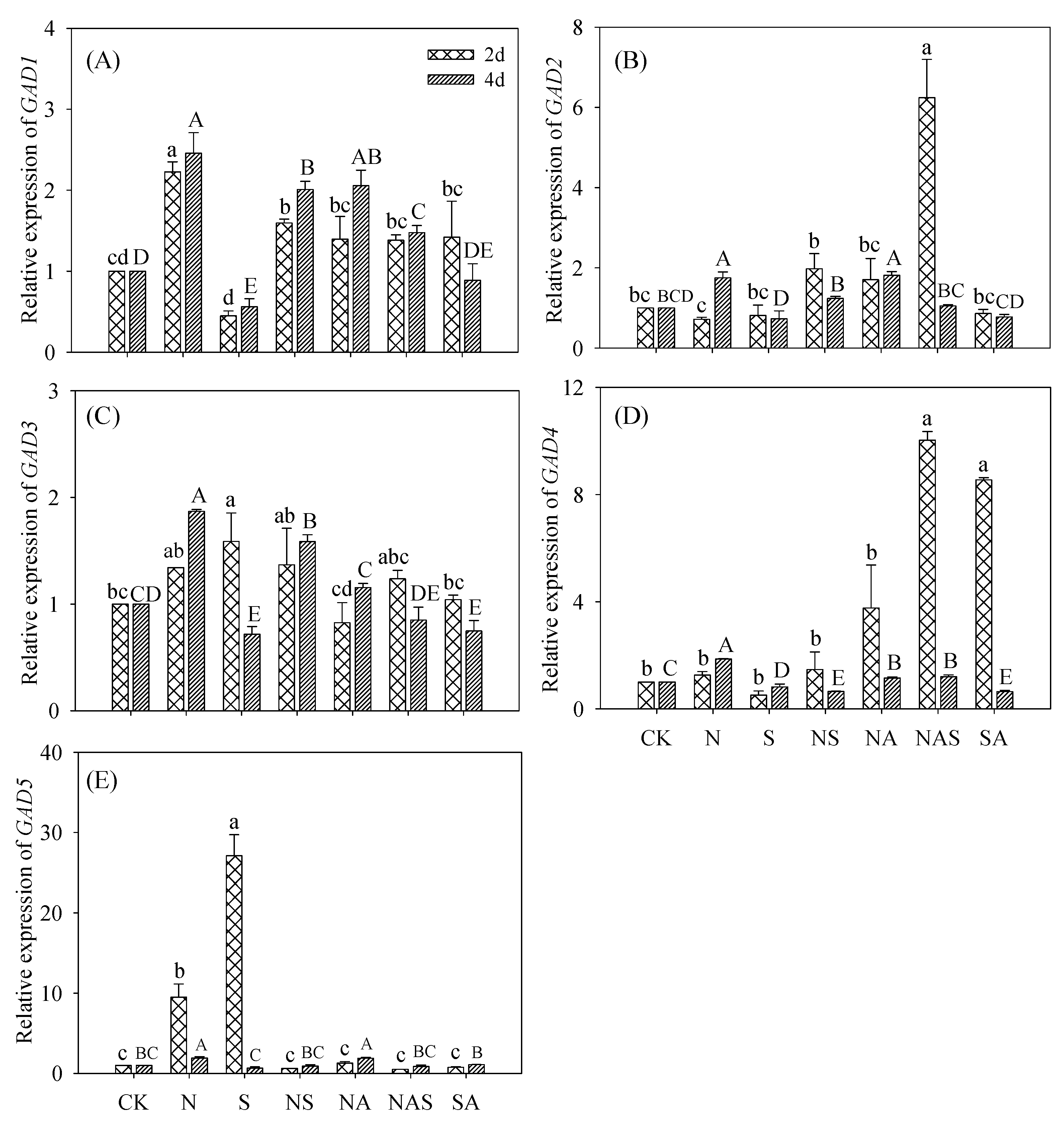
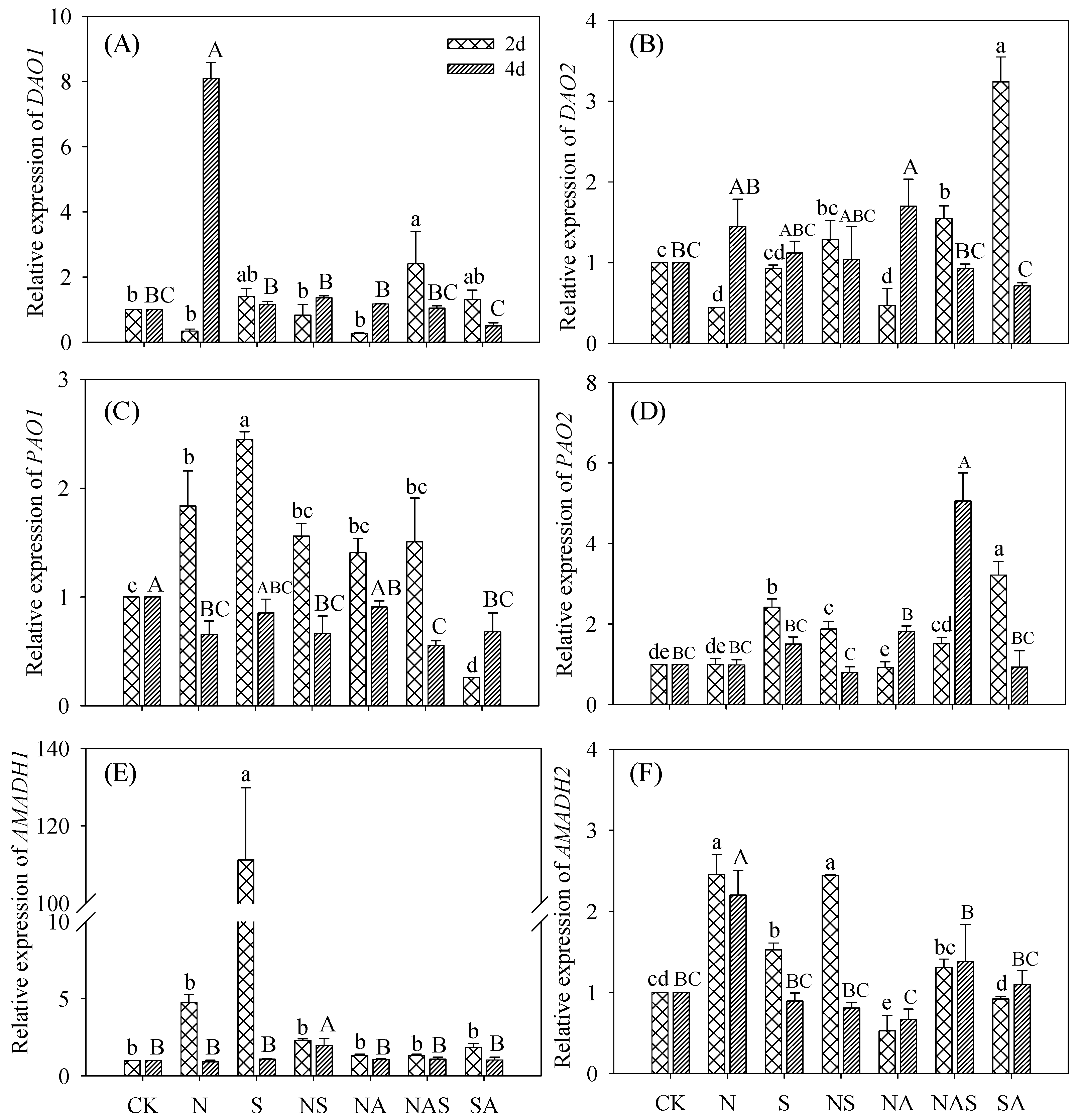
3.6. Changes in GABA Synthase Gene Expression
3.6.1. GADs Relative Expression
3.6.2. DAOs, PAOs, and AMADHs Relative Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lee, S.E.; Lee, Y.; Lee, G.H. The regulation of glutamic acid decarboxylases in GABA neurotransmission in the brain. Arch. Pharm. Res. 2019, 42, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Sved, A.F.; Sved, J.C. Plasticity of GABA function in the nucleus tractus solitarius in hypertension. Am. J. Physiol. Reg. Integr. Comp. Physiol. 2003, 285, R1272–R1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Wadei, H.A.; Ullah, M.F.; Al-Wadei, M. GABA (γ-aminobutyric acid), a non-protein amino acid counters the β-adrenergic cascade-activated oncogenic signaling in pancreatic cancer: A review of experimental evidence. Mol. Nutr. Food Res. 2011, 55, 1745–1758. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Luo, H.; Cheng, L.X.; Liu, K. Inhibitory role for GABA in atherosclerosis. Med. Hypotheses 2013, 81, 803–804. [Google Scholar] [CrossRef] [PubMed]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Poojary, M.M.; Dellarosa, N.; Roohinejad, S.; Koubaa, M.; Tylewicz, U.; Gómez-Galindo, F.; Saraiva, J.A.; Rosa, M.D.; Barba, F.J. Influence of innovative processing on γ-aminobutyric acid (GABA) contents in plant food materials. Compr. Rev. Food Sci. Food Saf. 2017, 16, 895–905. [Google Scholar] [CrossRef]
- Yin, Y.; Yang, R.; Guo, Q.; Gu, Z. NaCl stress and supplemental CaCl2 regulating GABA metabolism pathways in germinating soybean. Eur. Food Res. Tech. 2014, 238, 781–788. [Google Scholar] [CrossRef]
- Guo, Y.; Yang, R.; Chen, H.; Song, Y.; Gu, Z. Accumulation of γ-aminobutyric acid in germinated soybean (Glycine max L.) in relation to glutamate decarboxylase and diamine oxidase activity induced by additives under hypoxia. Eur. Food Res. Tech. 2012, 234, 679–687. [Google Scholar] [CrossRef]
- Wang, P.; Liu, K.; Gu, Z.; Yang, R. Enhanced γ-aminobutyric acid accumulation, alleviated componential deterioration and technofunctionality loss of germinated wheat by hypoxia stress. Food Chem. 2018, 269, 473–479. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945–1952. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Rahman, A.; Alam, M.; Mahmud, J.A.; Suzuki, T.; Fujita, M. Polyamines confer salt tolerance in mung bean (Vigna radiata L.) by reducing sodium uptake, improving nutrient homeostasis, antioxidant defense, and methylglyoxal detoxification systems. Front. Plant Sci. 2016, 7, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Seifikalhor, M.; Aliniaeifard, S.; Hassani, B.; Niknam, V.; Lastochkina, O. Diverse role of γ-aminobutyric acid in dynamic plant cell responses. Plant Cell Rep. 2019. [Google Scholar] [CrossRef] [PubMed]
- Nag, S.; Choudhuri, M. Role of endogenous auxin and polyamines in adventitious root formation in mungbean cuttings. Indian J. Plant Physiol. 2007, 12, 228–236. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, P.; Gu, Z.; Ma, M.; Yang, R. Spermidine improves antioxidant activity and energy metabolism in mung bean sprouts. Food Chem. 2020, 309, 125759. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.; Chen, Z.; Gong, B.; Wang, X.; Win, M.; Shi, Q.; Li, Y.; Yang, F. Effects of exogenous polyamines on nitrate tolerance in cucumber. Russ. J. Plant Physiol. 2016, 63, 549–557. [Google Scholar] [CrossRef]
- Yin, Y.; Qi, F.; Gao, L.; Rao, S.; Yang, Z.; Fang, W. iTRAQ-based quantitative proteomic analysis of dark-germinated soybeans in response to salt stress. RSC Adv. 2018, 8, 17905–17913. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of γ-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef]
- Yin, Y.; Yang, R.; Gu, Z. Calcium regulating growth and GABA metabolism pathways in germinating soybean (Glycine max L.) under NaCl stress. Eur. Food Res. Tech. 2014, 239, 149–156. [Google Scholar] [CrossRef]
- Rao, K.M.; Sresty, T. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Yang, R.; Guo, Q.; Gu, Z. GABA shunt and polyamine degradation pathway on γ-aminobutyric acid accumulation in germinating fava bean (Vicia faba L.) under hypoxia. Food Chem. 2013, 136, 152–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tanou, G.; Molassiotis, A.; Diamantidis, G. Induction of reactive oxygen species and necrotic death-like destruction in strawberry leaves by salinity. Env. Exp. Bot. 2009, 65, 270–281. [Google Scholar] [CrossRef]
- Zhang, G.; Xu, S.; Hu, Q.; Mao, W.; Gong, Y. Putrescine plays a positive role in salt-tolerance mechanisms by reducing oxidative damage in roots of vegetable soybean. J. Integr. Agric. 2014, 13, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Ahanger, M.A.; Qin, C.; Maodong, Q.; Dong, X.X.; Ahmad, P.; Abd_Allah, E.F.; Zhang, L. Spermine application alleviates salinity induced growth and photosynthetic inhibition in Solanum lycopersicum by modulating osmolyte and secondary metabolite accumulation and differentially regulating antioxidant metabolism. Plant Physiol. Biochem. 2019, 144, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Li, J.; Liu, S.; Zhu, X.; Chen, Y.; Shen, X. Effects of spermidine and salinity stress on growth and biochemical response of paraquat-susceptibe and-resistant goosegrass (Eleusine indica L.). Weed Biol. Manag. 2019, 19, 75–84. [Google Scholar] [CrossRef]
- Ramos-Ruiz, R.; Martinez, F.; Knauf-Beiter, G. The effects of GABA in plants. Cogent Food Agric. 2019, 5, 12–18. [Google Scholar] [CrossRef]
- Yang, R.; Feng, L.; Wang, S.; Yu, N.; Gu, Z. Accumulation of gamma-aminobutyric acid in soybean by hypoxia germination and freeze-thawing incubation. J. Sci. Food Agric. 2016, 96, 2090–2096. [Google Scholar] [CrossRef]
- Liao, J.; Wu, X.; Xing, Z.; Li, Q.; Duan, Y.; Fang, W.; Zhu, X. Gamma-aminobutyric acid (GABA) accumulation in tea (Camellia sinensis L.) through the GABA shunt and polyamine degradation pathways under anoxia. J. Agric. Food Chem. 2017, 65, 3013–3018. [Google Scholar] [CrossRef]
- Hyun, T.; Eom, S.; Jeun, Y.; Han, S.; Kim, J. Identification of glutamate decarboxylases as a γ-aminobutyric acid (GABA) biosynthetic enzyme in soybean. Ind. Crop. Prod. 2013, 49, 864–870. [Google Scholar] [CrossRef]
- Souza, S.; Mazzafera, P.; Sodek, L. Flooding of the root system in soybean: Biochemical and molecular aspects of N metabolism in the nodule during stress and recovery. Amino Acids 2016, 48, 1285–1295. [Google Scholar] [CrossRef]
- Hyun, T.; Eom, S.; Han, X.; Kim, J. Evolution and expression analysis of the soybean glutamate decarboxylase gene family. J. Biosci. 2014, 39, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Lapuz, R.R.; Javier, S.; Aquino, J.D.C.; Undan, J.R. Gene Expression and Sequence Analysis of BADH1 Gene in CLSU Aromatic Rice (Oryza sativa L.) Accessions Subjected to Drought and Saline Condition. J. Nutr. Sci. Vitam. 2019, 65, S196–S199. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences (5′–3′) |
|---|---|
| GAD1-F 1 | AGCAGGTGAAGAAAATGACGA |
| GAD1-R 2 | TCTTCTTCCTGTCCATCACAAA |
| GAD2-F | CCACTCACCCAGATGAAAAAG |
| GAD2-R | AGCTGCATCCTTTGGTATTGA |
| GAD3-F | TTCCTGTGTGAGTACGTGTGC |
| GAD3-R | CATCATTGCGCTCATAATCCT |
| GAD4-F | GGTGAGAAGATTAAGAAAGCTGC |
| GAD4-R | GGTAAGCCTAGCATGCTCCA |
| GAD5-F | CTCAGTGCAGAAGAAAATGGC |
| GAD5-R | ACACCCCCTTGAAGCTAACAC |
| DAO1-F | AGATGATACTTTAGCCGTTTGGAC |
| DAO1-R | CAATAAGCAGATCGCATTTCAC |
| DAO2-F | TCCTTGCAAGCAGTTGTGTC |
| DAO2-R | TGGTTTGATGGATTCCCATT |
| PAO2-F | GGCAGCTTTCTTGAAACACA |
| PAO2-R | TAACCCAAGTAGCCAAACCC |
| PAO2-F | CTTCCTCCAAGGAAAGCAAG |
| PAO2-R | GTGTCCACCTCGAGTTGTTG |
| AMADH1-F | TGAAGCTGGTGCTCCTTTGT |
| AMADH1-R | AAGATGGTCCATTCAGCAGT |
| AMADH2-F | TGAAGCGGGTGCTCCTTTAG |
| AMADH2-R | AATATGGTCCATTCAGCAGC |
| UBi-F | GTGTAATGTTGGATGTGTTCCC |
| UBi-R | ACACAATTGAGTTCAACACAAACCG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, W.; Qi, F.; Yin, Y.; Yang, Z. Exogenous Spermidine Promotes γ-Aminobutyric Acid Accumulation and Alleviates the Negative Effect of NaCl Stress in Germinating Soybean (Glycine max L.). Foods 2020, 9, 267. https://doi.org/10.3390/foods9030267
Fang W, Qi F, Yin Y, Yang Z. Exogenous Spermidine Promotes γ-Aminobutyric Acid Accumulation and Alleviates the Negative Effect of NaCl Stress in Germinating Soybean (Glycine max L.). Foods. 2020; 9(3):267. https://doi.org/10.3390/foods9030267
Chicago/Turabian StyleFang, Weiming, Fei Qi, Yongqi Yin, and Zhengfei Yang. 2020. "Exogenous Spermidine Promotes γ-Aminobutyric Acid Accumulation and Alleviates the Negative Effect of NaCl Stress in Germinating Soybean (Glycine max L.)" Foods 9, no. 3: 267. https://doi.org/10.3390/foods9030267
APA StyleFang, W., Qi, F., Yin, Y., & Yang, Z. (2020). Exogenous Spermidine Promotes γ-Aminobutyric Acid Accumulation and Alleviates the Negative Effect of NaCl Stress in Germinating Soybean (Glycine max L.). Foods, 9(3), 267. https://doi.org/10.3390/foods9030267