Impact of Processing and Intestinal Conditions on in Vitro Digestion of Chia (Salvia hispanica) Seeds and Derivatives



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Chia Products
2.2.1. Milling Process for Chia Flour
2.2.2. Seeds Germination for Chia Sprouts
2.3. In Vitro Digestion
2.4. Analytical Determinations
2.4.1. Moisture, Lipid, Protein and Carbohydrates
2.4.2. Proteolysis
2.4.3. Lipolysis
2.4.4. Calcium
2.4.5. Polyphenols
2.4.6. Antioxidant Activity
2.4.7. Particle Size Measurement
2.5. Statistical Anlysis
3. Results
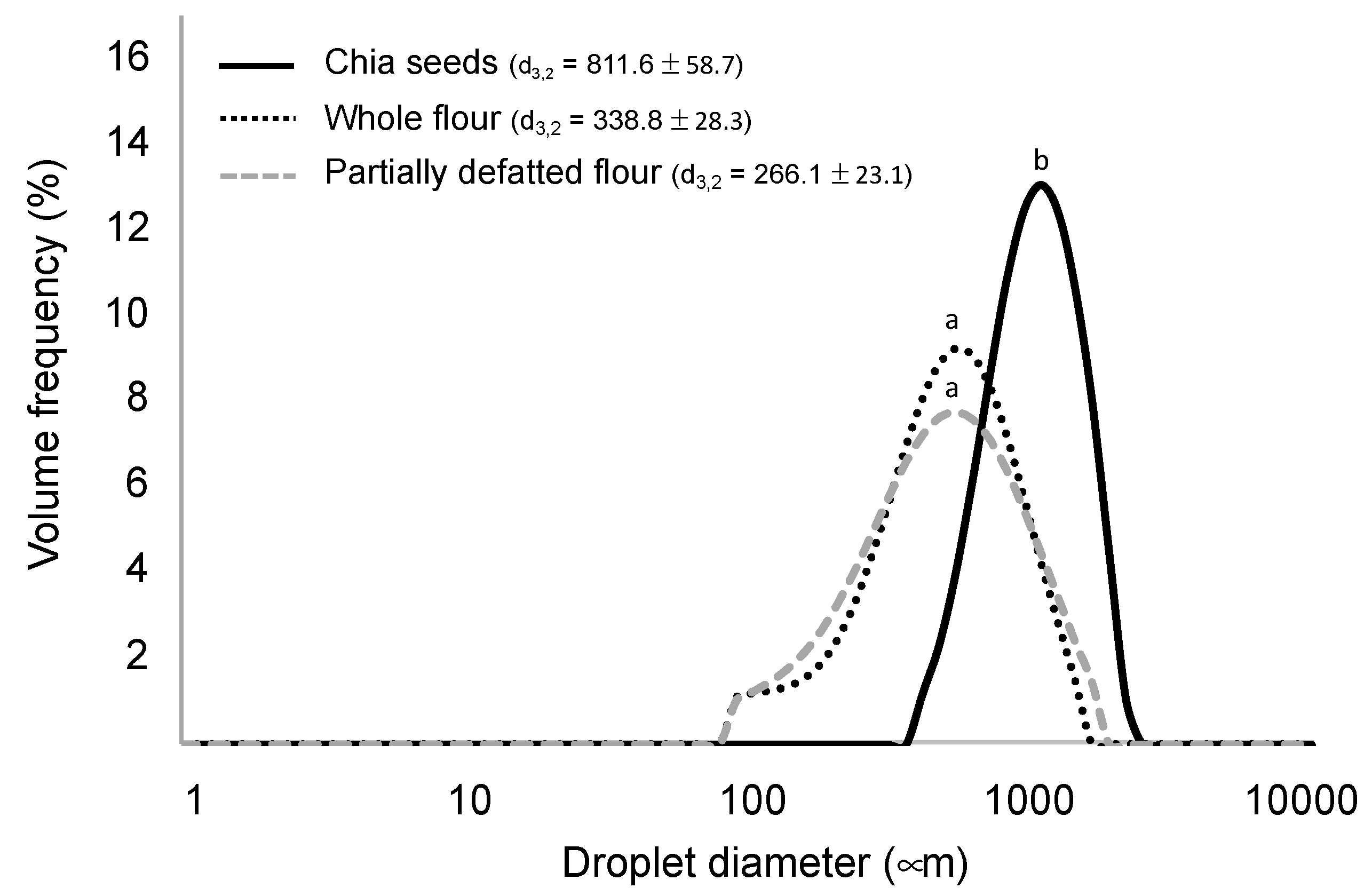
3.1. Chia Structures Matrix Characterisation
3.2. Lipid and Protein Digestibility
3.3. Bioaccessibility of Calcium and Polyphenols and Antioxidant Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Capitani, M.I.; Spotorno, V.; Nolasco, S.M.; Tomás, M.C. Physicochemical and functional characterization of by-products from chia (Salvia hispanica L.) seeds of Argentina. LWT-Food Sci. Technol. 2012, 45, 94–102. [Google Scholar] [CrossRef]
- Zettel, V.; Hitzmann, B. Applications of chia (Salvia hispanica L.) in food products. Trends Food Sci. Technol. 2018, 80, 43–50. [Google Scholar] [CrossRef]
- Reyes-Caudillo, E.; Tecante, A.; Valdivia-López, M.A. Dietary fibre content and antioxidant activity of phenolic compounds present in Mexican chia (Salvia hispanica L.) seeds. Food Chem. 2018, 107, 656–663. [Google Scholar] [CrossRef]
- Muñoz, L.A.; Cobos, A.; Diaz, O.; Aguilera, J.M. Chia seeds: Microstructure, mucilage extraction and hydration. J. Food Eng. 2018, 108, 216–224. [Google Scholar] [CrossRef]
- The European Commission. Commission implementing decision of 22 January 2013 authorizing an extension of use of chia seed as a novel food ingredient under Regulation (EC) No 258/97 of the European Parliament and of the Council. Off. J. Eur. Union 2013, 34–35. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32013D0050 (accessed on 31 January 2020).
- Inglett, G.E.; Chen, D.; Liu, S.X.; Lee, S. Pasting and rheological properties of oat products dry-blended with ground chia seeds. LWT-Food Sci. Technol. 2014, 55, 148–156. [Google Scholar] [CrossRef]
- Pellegrini, M.; Lucas-Gonzalez, R.; Fernández-López, J.; Ricci, A.; Pérez-Álvarez, J.A.; Sterzo, C.L.; Viuda-Martos, M. Bioaccessibility of polyphenolic compounds of six quinoa seeds during in vitro gastrointestinal digestion. J. Funct. Foods 2017, 38, 77–88. [Google Scholar] [CrossRef]
- Zieliński, H.; Frias, J.; Piskuła, M.K.; Kozłowska, H.; Vidal-Valverde, C. The effect of germination process on the superoxide dismutase-like activity and thiamine, riboflavin and mineral contents of rapeseeds. Food Chem. 2006, 99, 516–520. [Google Scholar] [CrossRef]
- Kylen, A.M.; McCready, R.M. Nutrients in seeds and sprouts of alfalfa, lentils, mung beans and soybeans. J. Food Sci. 1975, 40, 1008–1009. [Google Scholar] [CrossRef]
- Grundy, M.M.L.; Lapsley, K.; Ellis, P.R. A review of the impact of processing on nutrient bioaccessibility and digestion of almonds. Int. J. Food Sci. Technol. 2016, 51, 1937–1946. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Lerma, J.; Fornés-Ferrer, V.; Heredia, A.; Andrés, A. In vitro digestion models to assess lipolysis: The impact of the simulated conditions for gastrointestinal pH, bile salts and digestion fluids. Food Res. Int. 2019, 125, 108511. [Google Scholar] [CrossRef] [PubMed]
- Humbert, L.; Rainteau, D.; Tuvignon, N.; Wolf, C.; Seksik, P.; Laugier, R.; Carrière, F. Postprandial bile acid levels in intestine and plasma reveal altered biliary circulation in chronic pancreatitis patients. J. Lipid Res. 2018, 59, 2202–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelfond, D.; Ma, C.; Semler, J.; Borowitz, D. Intestinal pH and gastrointestinal transit profiles in cystic fibrosis patients measured by wireless motility capsule. Dig. Dis. Sci. 2013, 58, 2275–2281. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.J.; Smith, A.L.; Sly, P.D. Duodenal pH in cystic fibrosis and its relationship to fat malabsorption. Dig. Dis. Sci. 1990, 35, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Harries, J.T.; Muller, D.P.; McCollum, J.P.; Lipson, A.; Roma, E.; Norman, A.P. Intestinal bile salts in cystic fibrosis: Studies in the patient and experimental animal. Arch. Dis. Child. 1979, 54, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asensio-Grau, A.; Peinado, I.; Heredia, A.; Andrés, A. Effect of cooking methods and intestinal conditions on lipolysis, proteolysis and xanthophylls bioaccessibility of eggs. J. Funct. Foods. 2018, 46, 579–586. [Google Scholar] [CrossRef]
- Asensio-Grau, A.; Calvo-Lerma, J.; Heredia, A.; Andrés, A. Fat digestibility in meat products: Influence of food structure and gastrointestinal conditions. Int. J. Food. Sci. Nutr. 2019, 70, 530–539. [Google Scholar] [CrossRef]
- Paz-Yépez, C.; Peinado, I.; Heredia, A.; Andrés, A. Influence of particle size and intestinal conditions on in vitro lipid and protein digestibility of walnuts and peanuts. Food Res. Int. 2019, 119, 951–959. [Google Scholar] [CrossRef]
- Paz-Yépez, C.; Peinado, I.; Heredia, A.; Andrés, A. Lipids digestibility and polyphenols release under in vitro digestion of dark, milk and white chocolate. J. Funct. Foods 2019, 52, 196–203. [Google Scholar]
- Calvo-Lerma, J.; Fornés-Ferrer, V.; Heredia, A.; Andrés, A. In Vitro Digestion of Lipids in Real Foods: Influence of Lipid Organization Within the Food Matrix and Interactions with Nonlipid Components. J. Food Sci. 2018, 83, 2629–2637. [Google Scholar] [CrossRef] [Green Version]
- Ixtaina, V.Y.; Martínez, M.L.; Spotorno, V.; Mateo, C.M.; Maestri, D.M.; Diehl, B.W.; Nolasco, S.M.; Tomás, M.C. Characterization of chia seed oils obtained by pressing and solvent extraction. J. Food Compos. Anal. 2011, 24, 166–174. [Google Scholar] [CrossRef]
- Pająk, P.; Socha, R.; Broniek, J.; Królikowska, K.; Fortuna, T. Antioxidant properties, phenolic and mineral composition of germinated chia, golden flax, evening primrose, phacelia and fenugreek. Food Chem. 2019, 275, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Parker, M.L.; Grundy, M.M.-L.; Grassby, T.; Smeriglio, A.; Bisignano, C.; Raciti, R.; Trombetta, D.; Baer, D.; Wilde, P.J. Understanding the effect of particle size and processing on almond lipid bioaccessibility through microstructural analysis: From mastication to faecal collection. Nutrients 2018, 10, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aburub, A.; Fischer, M.; Camilleri, M.; Semler, J.R.; Fadda, H.M. Comparison of pH and motility of the small intestine of healthy subjects and patients with symptomatic constipation using the wireless motility capsule. Int. J. Pharm. 2018, 544, 158–164. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrier, F.; Boutrou, R.; Correding, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food–an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourieu-Lacanal, C.; Boutrou, R.; Carrier, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of AOAC International, 17th ed.; AOAC International: Gaithersberg, MD, USA, 2000. [Google Scholar]
- Lamothe, S.; Azimy, N.; Bazinet, L. Couillard, C.; Britten, M. Function Interaction of green tea poly-phenols with dairy matrices in a simulated gastrointestinal environment. Food Funct. 2014, 5, 2621–2631. [Google Scholar] [CrossRef]
- Bax, M.-L.; Aubry, L.; Ferreira, C.; Daudin, J.-D.; Gatellier, P.; Rémond, D.; Santé-Lhoutellier, V. Cooking temperature is a key derminant of in vitro meat protein digestion rate: Investigation of underlying mechanisms. J. Agric. Food Chem. 2012, 60, 2569–2576. [Google Scholar] [CrossRef]
- Lamothe, S.; Corbeil, M.-M.; Turgeon, S.L.; Britten, M. Influence of cheese matrix on lipid digestion in a simulated gastro-intestinal environment. Food Funct. 2012, 3, 724–731. [Google Scholar] [CrossRef]
- Barrera, C.; Betoret, N.; Corell, P.; Fito, P. Effect of osmotic dehydration on the stabilization of calcium-fortified apple slices (var. Granny Smith): Influence of operating variables on process kinetics and compositional changes. J. Food Eng. 2009, 2, 416–424. [Google Scholar] [CrossRef]
- Noël, L.; Carl, M.; Vastel, C.; Guérin, T. Determination of sodium, potassium, calcium and magnesium content in milk products by flame atomic absorption spectrometry (FAAS): A joint ISO/IDF collaborative study. Int. Dairy J. 2008, 18, 899–904. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Antolovich, M.; Prenzler, P.; Patsalides, E.; McDonald, S.; Robards, K. Methods for testing antioxidant activity. Analyst 2002, 127, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; McClements, D.J.; Decker, E.A. Impact of whey protein emulsifiers on the oxidative stability of salmon oil-in-water emulsions. J. Agr. Food Chem. 2003, 51, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Ye, A.; Lad, M.; Ferrua, M.; Dalgleish, D.; Singh, H. Disintegration kinetics of food gels during gastric digestion and its role on gastric emptying: An in vitro analysis. Food Funct. 2015, 6, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.Y.; Hettiarachchy, N.S.; Rath, N. Extraction, denaturation and hydrophobic properties of rice flour proteins. J. Food Sci. 2001, 66, 229–232. [Google Scholar] [CrossRef]
- Mostafa, M.M.; Rahma, E.H.; Rady, A.H. Chemical and nutritional changes in soybean during germination. Food Chem. 1987, 23, 257–275. [Google Scholar] [CrossRef]
- Mayer, A.; Poljakoff-Mayber, A. The Germination of Seeds, 3rd ed.; Pergamon: Oxford, UK, 1982. [Google Scholar]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted grains: A comprehensive review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Ortiz, F.A.; San Martín-Martínez, E.; Valverde, M.E.; Rodríguez-Aza, Y.; Berríos, J.D.J.; Mora-Escobedo, R. Profile analysis and correlation across phenolic compounds, isoflavones and antioxidant capacity during germination of soybeans (Glycine max L.). CyTA-J. Food 2017, 15, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Hsu, W.H.; Hollis, J.H. The impact of food viscosity on eating rate, subjective appetite, glycemic response and gastric emptying rate. PLoS ONE 2013, 8, e67482. [Google Scholar] [CrossRef] [Green Version]
- Logan, K.; Wright, A.J.; Goff, H.D. Correlating the structure and in vitro digestion viscosities of different pectin fibers to in vivo human satiety. Food Funct. 2015, 6, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Desnuelle, P.; Savary, P. Specificities of lipases. J. Lip. Res. 1963, 4, 369–384. [Google Scholar]
- Cui, L.; Gao, L.; Zheng, M.; Li, J.; Zhang, L.; Wu, Y.; Quiao, L.; Xu, C.; Wang, K.; Huang, D. Bioaccessibility of short chain chlorinated paraffins in meat and seafood. Sci. Total Environ. 2019, 668, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Benarous, K.; Djeridane, A.; Kameli, A.; Yousfi, M. Inhibition of Candida rugosa lipase by secondary metabolites extracts of three Algerian plants and their antioxidant activities. Cur. Enz. Inhib. 2013, 9, 75–82. [Google Scholar] [CrossRef]
- Alpers, D. Digestion and absorption of carbohydrates and proteins. In Physiology of the Gastrointestinal Tract; Johnson, L., Ed.; Raven Press: New York, NY, USA, 1994; Volume 2, pp. 1723–1749. [Google Scholar]
- Taylor, J.R.N.; Novellie, L.; Liebenberg, N.V.D.W. Protein body degradation in the starchy endosperm of germinating sorghum. J. Exp. Bot. 1985, 36, 1287–1295. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Kirleis, A.W.; Mertz, E.T.; Axtell, J.D. Effect of cooking on the protein profiles and in vitro digestibility of sorghum and maize. J. Agric. Food Chem. 1986, 34, 647–649. [Google Scholar] [CrossRef]
- James, W.P.T.; Branch, W.J.; Southgate, D.A.T. Calcium binding by dietary fibre. Lancet 1978, 311, 638–639. [Google Scholar] [CrossRef]
- Hu, M.; Li, Y.; Decker, E.A.; McClements, D.J. Role of calcium and calcium-binding agents on the lipase digestibility of emulsified lipids using an in vitro digestion model. Food Hydrocoll. 2010, 24, 719–725. [Google Scholar] [CrossRef]
- Govers, M.J.; Van der Meet, R. Effects of dietary calcium and phosphate on the intestinal interactions between calcium, phosphate, fatty acids, and bile acids. Gut 1993, 34, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; Da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.J.; De Camargo, A.C.; Shahidi, F. Phenolic and polyphenolic profiles of chia seeds and their in vitro biological activities. J. Funct. Food 2017, 35, 622–634. [Google Scholar] [CrossRef]
- Akillioglu, H.G.; Karakaya, S. Changes in total phenols, total flavonoids, and antioxidant activities of common beans and pinto beans after soaking, cooking, and in vitro digestion process. Food Sci. Biotech. 2010, 19, 633–639. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Hidalgo, M.; Sánchez-Moreno, C.; De Pascual-Teresa, S. Flavonoid–flavonoid interaction and its effect on their antioxidant activity. Food Chem. 2010, 121, 691–696. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Stage | Oral | Gastric | Intestinal |
|---|---|---|---|
| Input | Food sample (5 g) | Oral bolus (10 mL) | Gastric chyme (20 mL) |
| Digestive fluid | Simulated salivary fluid (5 mL) | Simulated gastric fluid (10 mL) | Simulated intestinal fluid (20 mL) |
| Composition of digestive fluid | α-amylase (75 U/mL) | Pepsin (2000 U/mL) | Pancreatin (Lipase: 30 LU/mL; Protease 1.8 U/mL; α-amylase: 12 U/mL) Bile salts (1 or 10 mM *) |
| Medium pH | 7 | 3 | 6 or 7 * |
| Temperature | 37 °C | 37 °C | 37 °C |
| Duration | 5 min | 120 min | 120 min |
| Mechanical force | Grinder pulses | Head-over heels agitation (100 rpm) | Head-over heels agitation (60 rpm) |
| Chia Seeds and Whole Flour | Partially Defatted Flour | Chia Sprouts | |
|---|---|---|---|
| Moisture (g/g dry matter) | 0.065 (0.008) a | 0.068 (0.009) a | 9.87 (0.010) b |
| Lipids (g/g dry matter) | 0.325 (0.011) c | 0.197 (0.007) b | 0.097 (0.003) a |
| Protein (g/g dry matter) | 0.201 (0.007) a | 0.291 (0.011) c | 0.229 (0.009) b |
| Carbohydrate (g/g dry matter) | 0.472 (0.016) a | 0.501 (0.017) b | 0.644 (0.022) c |
| Calcium (mg/g dry matter) | 6.46 (0.046) b | 3.45 (0.032) a | 7.26 (0.18) c |
| Polyphenols (mg GA eq./g dry matter) | 1.78 (0.03) b | 1.24 (0.02) a | 2.87 (0.06) c |
| Antioxidant activity (mg TX eq./g dry matter) | 3.49 (0.11) b | 2.58 (0.09) a | 5.69 (1.6) c |
| Intestinal Conditions | Chia Seeds | Whole Chia Flour | Partially Defatted Chia Flour | Chia Sprouts | |
|---|---|---|---|---|---|
| Calcium (mg/g dry matter) | Standard | 3.82 (0.03) Ca | 4.22 (0.03) Da | 2.09 (0.07) Ba | 0.15 (0.02) Aa |
| Altered | 3.77 (0.04) Ca | 4.28 (0.02) Da | 2.07 (0.03) Ba | 0.32 (0.04) Ab | |
| Polyphenols mg (GA eq./g dry matter) | Standard | 1.81 (0.17) Ab | 6.13 (0.38) Db | 2.67 (0.28) Bb | 4.41 (0.51) Cb |
| Altered | 1.51 (0.15) Aa | 3.91 (0.16) Da | 2.13 (0.13) Ba | 3.55 (0.13) Ca | |
| Antioxidant activity (mg TX eq./g dry matter) | Standard | 1.17 (0.06) Ab | 4.8 (0.38) Cb | 3.97 (0.75) Bb | 4.21 (1.17) Bb |
| Altered | 1.03 (0.09) Aa | 3.31 (0.33) Ca | 2.53 (0.39) Ba | 2.23 (0.74) Ba |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvo-Lerma, J.; Paz-Yépez, C.; Asensio-Grau, A.; Heredia, A.; Andrés, A. Impact of Processing and Intestinal Conditions on in Vitro Digestion of Chia (Salvia hispanica) Seeds and Derivatives. Foods 2020, 9, 290. https://doi.org/10.3390/foods9030290
Calvo-Lerma J, Paz-Yépez C, Asensio-Grau A, Heredia A, Andrés A. Impact of Processing and Intestinal Conditions on in Vitro Digestion of Chia (Salvia hispanica) Seeds and Derivatives. Foods. 2020; 9(3):290. https://doi.org/10.3390/foods9030290
Chicago/Turabian StyleCalvo-Lerma, Joaquim, Carolina Paz-Yépez, Andrea Asensio-Grau, Ana Heredia, and Ana Andrés. 2020. "Impact of Processing and Intestinal Conditions on in Vitro Digestion of Chia (Salvia hispanica) Seeds and Derivatives" Foods 9, no. 3: 290. https://doi.org/10.3390/foods9030290
APA StyleCalvo-Lerma, J., Paz-Yépez, C., Asensio-Grau, A., Heredia, A., & Andrés, A. (2020). Impact of Processing and Intestinal Conditions on in Vitro Digestion of Chia (Salvia hispanica) Seeds and Derivatives. Foods, 9(3), 290. https://doi.org/10.3390/foods9030290