A Glycoside Hydrolase Family 62 A-L-Arabinofuranosidase from Trichoderma Reesei and Its Applicable Potential during Mashing

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Enzyme Production
2.3. Protein Concentration Estimation
2.4. Enzyme Activity Assay
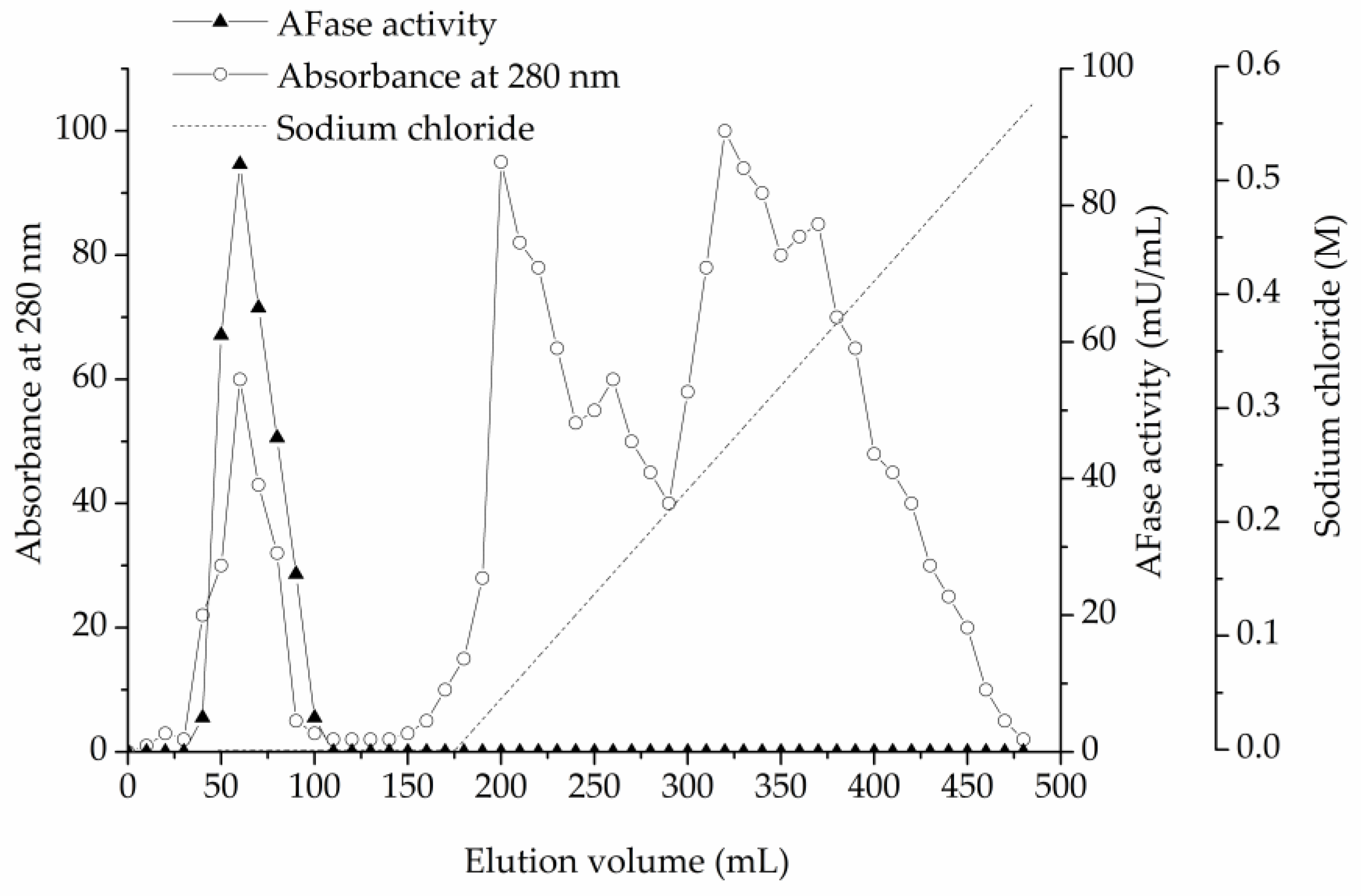
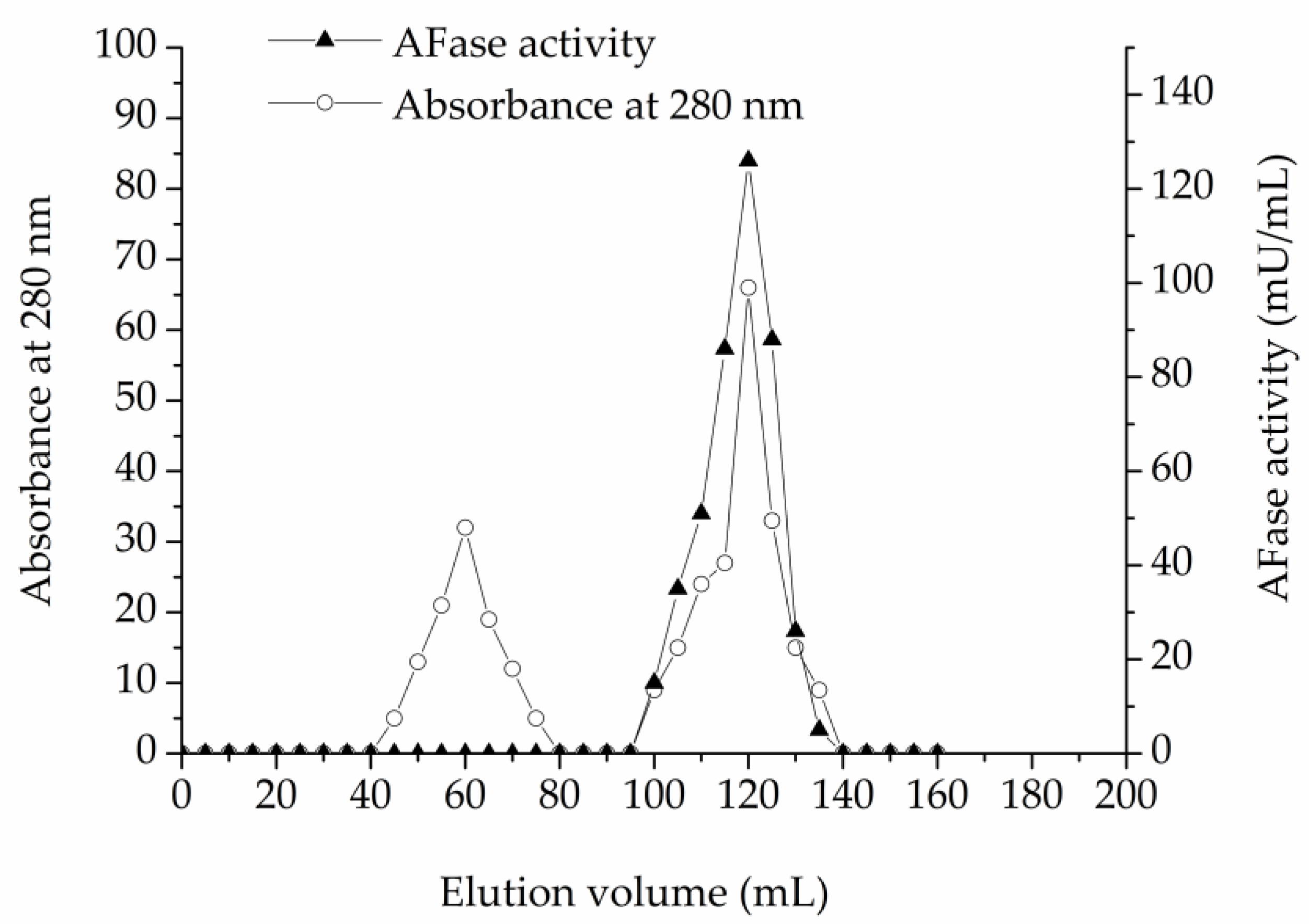
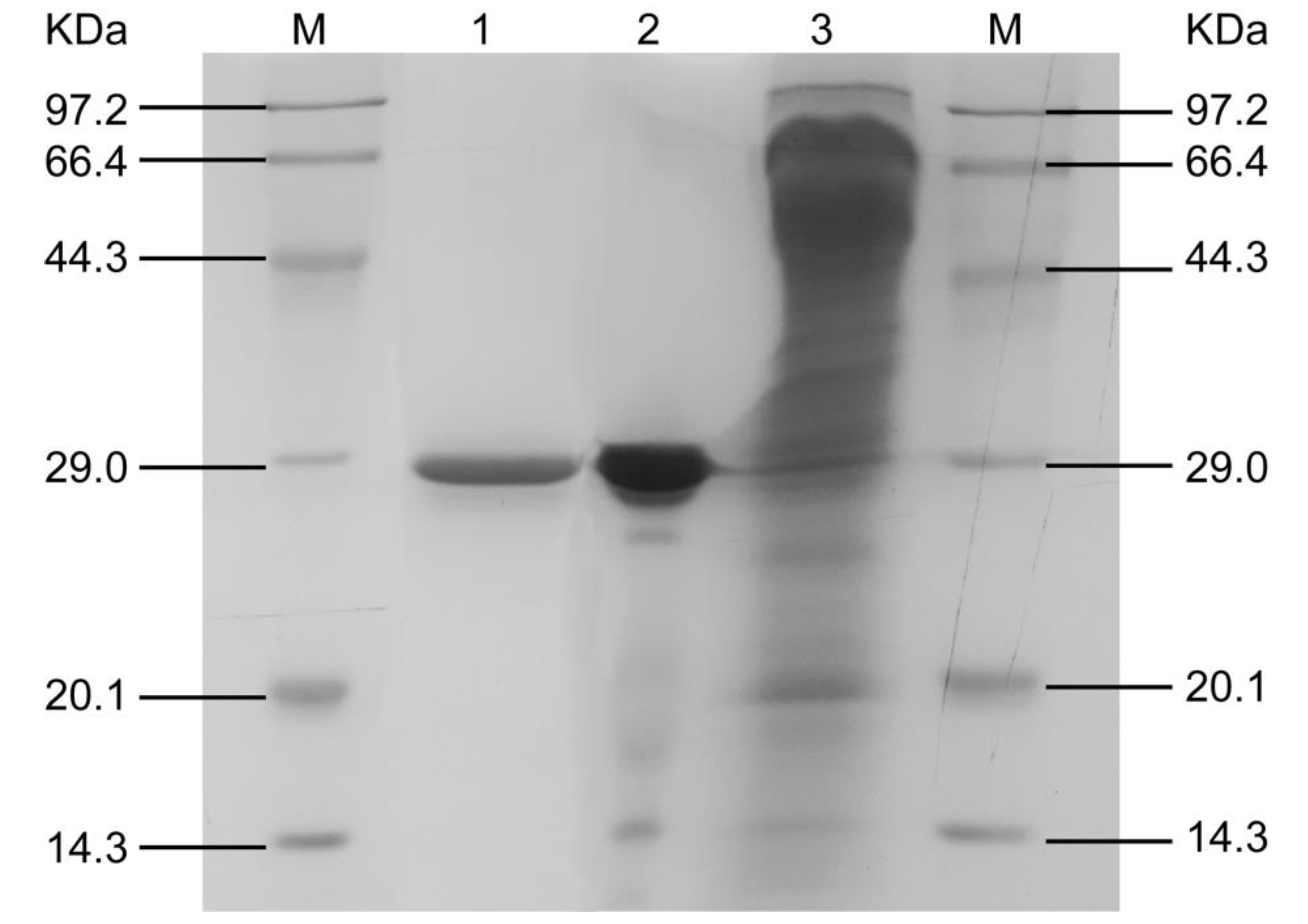
2.5. Enzyme Purification
2.6. Electrophoresis
2.7. Protein Identification
2.8. Substrate Specificity
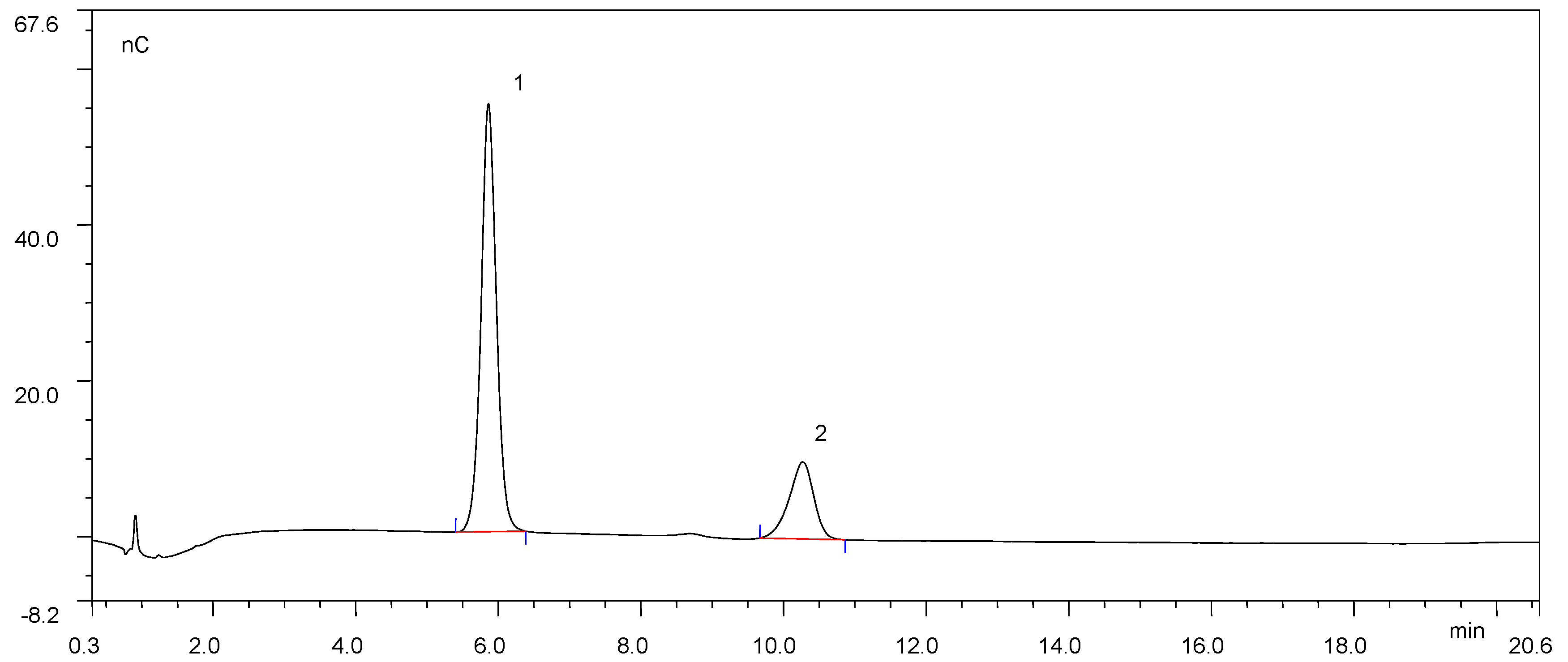
2.9. Hydrolytic Products Analysis
2.10. Mash Preparation with Supplemented AFase
3. Results and Discussion
3.1. Enzyme Purification
3.2. Substrate Specificity
3.3. Hydrolytic Products Analysis
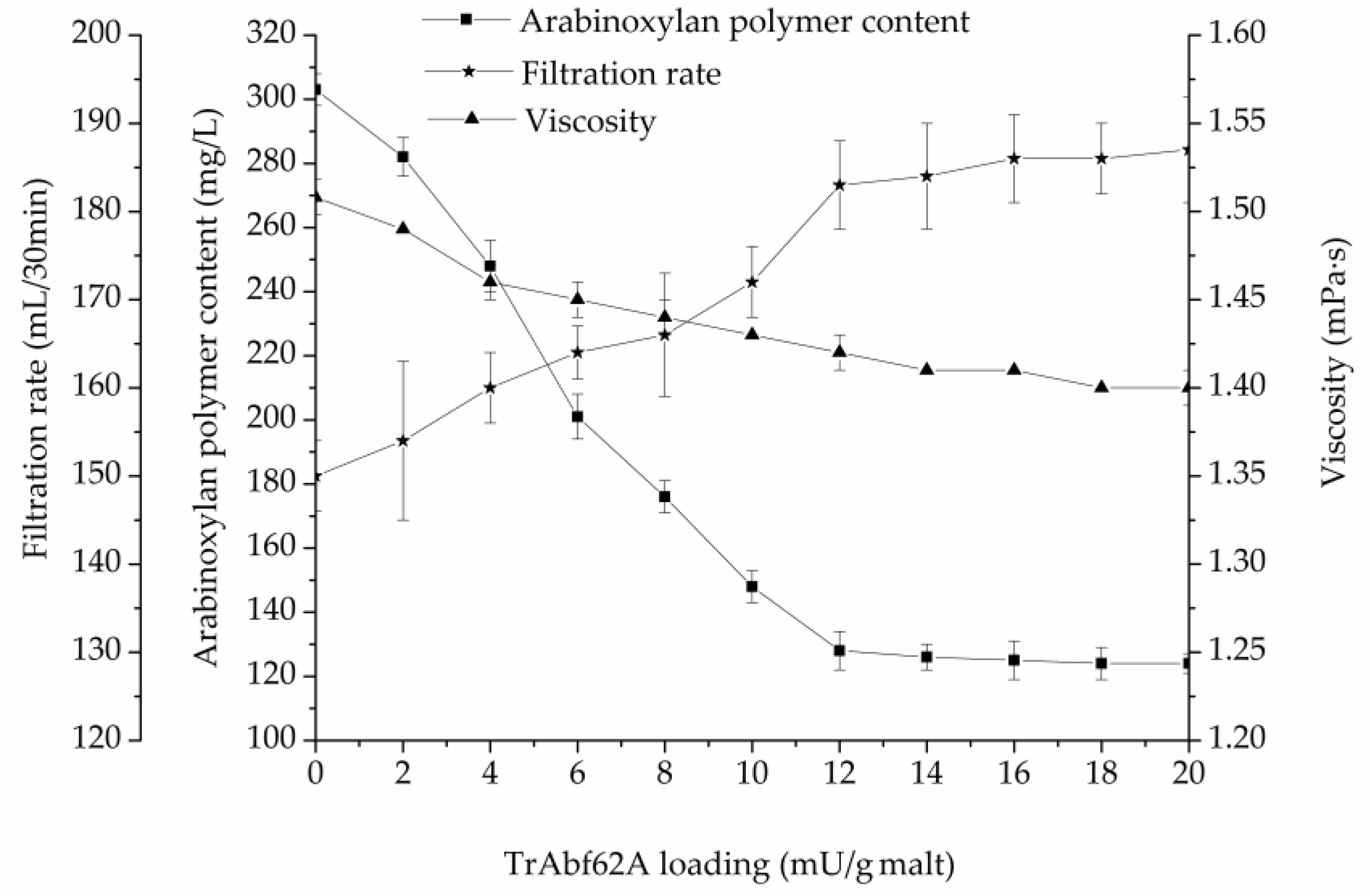
3.4. Supplementation of TrAbf62A during Mashing with Barley Malt
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kanauchi, M.; Ishikura, W.; Bamforth, C.W. β-Glucans and pentosans and their degradation products in commercial beers. J. Inst. Brew. 2011, 117, 120–124. [Google Scholar] [CrossRef]
- Kupetz, M.; Aumer, J.; Harms, D.; Zarnkow, M.; Sacher, B.; Becker, T. High-throughput β-glucan analyses and their relationship with beer filterability. Eur. Food Res. Technol. 2017, 243, 341–351. [Google Scholar] [CrossRef]
- Stewart, D.C.; Hawthorne, D.; Evans, D.E. Cold sterile filtration: A small scale filtration tests and investigation of membrane plugging. J. Inst. Brew. 1998, 104, 321–326. [Google Scholar] [CrossRef]
- Sadosky, P.; Schwarz, P.; Horsley, R. Effect of arabinoxylans, β-glucans, and dextrins on the viscosity and membrane filterability of a beer model solution. J. Am. Soc. Brew. Chem. 2002, 60, 153–162. [Google Scholar] [CrossRef]
- Li, M.M.; Du, J.H.; Zheng, Y.X. Non-starch polysaccharides in wheat beers and barley malt beers: A comparative study. Foods 2020, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Rosicka-Kaczmarek, J.; Komisarczyk, A.; Nebesny, E.; Makowski, B. The influence of arabinoxylans on the quality of grain industry products. Eur. Food Res. Technol. 2016, 242, 295–303. [Google Scholar] [CrossRef]
- Kupetz, M.; Zeh, A.; Fischer, S.; Becker, T. Investigation of filtration-inhibitory substances in german wheat beer. Brew. Sci. 2017, 70, 1–8. [Google Scholar]
- Hassan, A.S.; Houston, K.; Lahnstein, J.; Shirley, N.; Schwerdt, J.G.; Gidley, M.J.; Waugh, R.; Little, A.; Burton, R.A. A genome wide association study of arabinoxylan content in 2-row spring barley grain. PLoS ONE 2017, 12, e0182537. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.; Freeman, G.; Evans, E. Development and assessment of a small-scale wort filtration test for the prediction of beer filtration efficiency. J. Inst. Brew. 2000, 106, 361–366. [Google Scholar] [CrossRef]
- Li, Y.; Lu, J.; Gu, G.X.; Shi, Z.P.; Mao, Z.G. Studies on water-extractable arabinoxylans during malting and brewing. Food Chem. 2005, 93, 33–38. [Google Scholar] [CrossRef]
- Du, Y.L.; Shi, P.J.; Huang, H.Q.; Zhang, X.; Luo, H.Y.; Wang, Y.R.; Yao, B. Characterization of three novel thermophilic xylanases from Humicola insolens Y1 with application potentials in the brewing industry. Bioresour. Technol. 2013, 130, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Meng, K.; Bai, Y.G.; Shi, P.J.; Huang, H.Q.; Luo, H.Y.; Wang, Y.R.; Yang, P.L.; Song, W.; Yao, B. Two family 11 xylanases from Achaetomium sp. Xz-8 with high catalytic efficiency and application potentials in the brewing industry. J. Agric. Food Chem. 2013, 61, 6880–6889. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Luo, H.Y.; Yu, W.N.; Ma, R.; You, S.; Liu, W.N.; Hou, L.Y.; Zheng, F.; Xie, X.M.; Yao, B. A thermostable Gloeophyllum trabeum xylanase with potential for the brewing industry. Food Chem. 2016, 199, 516–523. [Google Scholar] [CrossRef]
- Walia, A.; Guleria, S.; Mehta, P.; Chauhan, A.; Parkash, J. Microbial xylanases and their industrial application in pulp and paper biobleaching: A review. 3 Biotech 2017, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Wilkens, C.; Andersen, S.; Dumon, C.; Berrin, J.G.; Svensson, B. GH62 arabinofuranosidases: Structure, function and applications. Biotechnol. Adv. 2017, 35, 792–804. [Google Scholar] [CrossRef] [Green Version]
- Bischof, R.H.; Ramoni, J.; Seiboth, B. Cellulases and beyond: The first 70 years of the enzyme producer Trichoderma reesei. Microb. Cell Fact. 2016, 15, 106. [Google Scholar] [CrossRef] [Green Version]
- Ramoni, J.; Marchetti-Deschmann, M.; Seidl-Seiboth, V.; Seiboth, B. Trichoderma reesei xylanase 5 is defective in the reference strain QM6a but functional alleles are present in other wild-type strains. Appl. Microbiol. Biotechnol. 2017, 101, 4139–4149. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.Y.; Lu, J.; Xie, G.F. Secretome analysis of Trichoderma reesei CICC41495 for degradation of arabinoxylan in malted barley. J. Inst. Brew. 2018, 124, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Mandels, M.; Weber, J. The production of cellulases. Adv. Chem. 1969, 95, 391–414. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Oh, G.W.; Kang, Y.; Choi, C.Y.; Kang, S.Y.; Kang, J.H.; Lee, M.L.; Han, N.S.; Kim, T.J. Detailed mode of action of arabinan-debranching α-L-arabinofuranosidase GH51 from Bacillus velezensis. J. Microbiol. Biotechnol. 2019, 29, 37–43. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural protein during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bienvenut, W.V.; Déon, C.; Pasquarello, C.; Campbell, J.M.; Sanchez, J.C.; Vestal, M.L.; Hochstrasser, D.F. Matrix-assisted laser desorption/ionization-tandem mass spectrometry with high resolution and sensitivity for identification and characterization of proteins. Proteomics 2002, 2, 868–876. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Biochem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Wu, S.J. Mulberry leaf polysaccharides suppress renal fibrosis. Int. J. Biol. Macromol. 2019, 124, 1090–1093. [Google Scholar] [CrossRef] [PubMed]
- European Brewery Convention. Analytica–EBC, 5th ed.; Fachverlag Hans Carl: Nürnberg, Germany, 1998; Volume 4, pp. 1–3. [Google Scholar]
- Cleemput, G.; Roels, S.P.; Van Oort, M.; Grobet, J.; Delcour, J.A. Heterogeneity in the structure of water-soluble arabinoxylans in European wheat flours of variable bread-making quality. Cereal Chem. 1993, 70, 324–329. [Google Scholar]
- Douglas, S.G. A rapid method for the determination of pentosans in wheat flour. Food Chem. 1981, 7, 139–145. [Google Scholar] [CrossRef]
- Martinez, D.; Berka, R.M.; Henrissat, B.; Saloheimo, M.; Arvas, M.; Baker, S.E.; Chapman, J.; Chertkov, O.; Coutinho, P.M.; Cullen, D.; et al. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat. Biotechnol. 2008, 26, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.K.; Schmoll, M.; Herrera-Estrella, A.; Upadhyay, R.S.; Druzhinina, I.; Tuohy, M.G. Biotechnology and Biology of Trichoderm, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 339–344. [Google Scholar]
- Gong, W.L.; Zhang, H.Q.; Liu, S.J.; Zhang, L.L.; Gao, P.J.; Chen, G.J.; Wang, L.S. Comparative secretome analysis of Aspergillus niger, Trichoderma reesei, and Penicillium oxalicum during solid-state fermentation. Appl. Biochem. Biotechnol. 2015, 177, 1252–1271. [Google Scholar] [CrossRef] [PubMed]
- Poutanen, K. An α-L-arabinofuranosidase of Trichoderma reesei. J. Biotechnol. 1988, 7, 271–281. [Google Scholar] [CrossRef]
- Nogawa, M.; Yatsui, K.; Tomioka, A.; Okada, H.; Morikawa, Y. An α-L-arabinofuranosidase from Trichoderma reesei containing a noncatalytic xylan-binding domain. Appl. Environ. Microbiol. 1999, 65, 3964–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormelink, F.J.M.; Voragen, A.G.J. Degradation of different [(glucurono)arabino]xylans by a combination of purified xylan-degrading enzymes. Appl. Microbiol. Biot. 1993, 38, 688–695. [Google Scholar] [CrossRef]
- Barron, C.; Robert, P.; Guillon, F.; Saulnier, L.; Rouau, X. Structural heterogeneity of wheat arabinoxylans revealed by Raman spectroscopy. Carbohydr. Res. 2006, 341, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Geng, A.L.; Wu, J.; Xie, R.G.; Wang, H.C.; Wu, Y.F.; Li, X.; Chang, F.X.; Sun, J.Z. Highly thermostable GH51 α-arabinofuranosidase from Hungateiclostridium clariflavum DSM 19732. Appl. Microbiol. Biot. 2019, 103, 3783–3793. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, H.R.; Pedersen, S.; Meyer, A.S. Synergistic enzyme mechanisms and effects of sequential enzyme additions on degradation of water insoluble wheat arabinoxylan. Enzyme Microb. Technol. 2007, 40, 908–918. [Google Scholar] [CrossRef]
- Adams, E.L.; Kroon, P.A.; Williamson, G.; Gilbert, H.J.; Morris, V.J. Inactivated enzymes as probes of the structure of arabinoxylans as observed by atomic force microscopy. Carbohydr. Res. 2004, 339, 579–590. [Google Scholar] [CrossRef]
- Phuengmaunga, P.; Kunishigeb, Y.; Sukhumsiricharta, W.; Sakamotob, T. Identification and characterization of GH62 bacterial α-L-arabinofuranosidase from thermotolerant Streptomyces sp. SWU10 that preferentially degrades branched L-arabinofuranoses in wheat arabinoxylan. Enzyme Microb. Technol. 2018, 112, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Vincent, P.; Shareck, F.; Dupont, C.; Kluepfel, D. New α-L-arabinofuranosidase produced by Streptomyces lividans: Cloning and DNA sequence of the abfB gene and characterization of the enzyme. Biochem. J. 1997, 15, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Ogura, A.; Inui, M.; Tokuda, S.; Hosokawa, S.; Kasai, N. Identification of a GH62 α-L-arabinofuranosidase specific for arabinoxylan produced by Penicillium chrysogenum. Appl. Microbiol. Biotechnol. 2011, 90, 137–146. [Google Scholar] [CrossRef]
- Hu, Y.B.; Yan, X.C.; Zhang, H.; Liu, J.Q.; Luo, F.; Cui, Y.Y.; Wang, W.Y.; Zhou, Y.F. Cloning and expression of a novel α-1,3-arabinofuranosidase from Penicillium oxalicum sp. 68. AMB Express 2018, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Sarch, C.; Suzuki, H.; Master, E.R.; Wang, W.J. Kinetics and regioselectivity of three GH62 α-L-arabinofuranosidases from plant pathogenic fungi. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Tanaka, M.; Tsumuraya, Y.; Hashimoto, Y. α-L-Arabinofuranosidase from radish (Raphanus sativus L.) seeds. Plant Physiol. 1992, 100, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.X.; Bai, Y.G.; Yang, P.L.; Luo, H.Y.; Huang, H.Q.; Meng, K.; Shi, P.J.; Wang, Y.R.; Yao, B. A novel bifunctional GH51 exo-α-L-arabinofuranosidase/endo-xylanase from Alicyclobacillus sp. A4 with significant biomass-degrading capacity. Biotechnol. Biofuels 2015, 8, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, T.M.; Macrae, S.I. Arabinoxylan-degrading enzymes systems of the fungus Aspergillus awamori: Purification and properties of an α-L-arabinofuranosidase. App. Microbiol. Biotechnol. 1996, 45, 538–545. [Google Scholar] [CrossRef]
- Lee, R.C.; Hrmova, M.; Burton, R.A.; Lahnstein, J.; Fincher, G.B. Bifunctional family 3 glycoside hydrolases from barley with α-L-arabinofuranosidase and β-D-xylosidase activity. Characterization, primary structures, and COOH-terminal processing. J. Biol. Chem. 2003, 278, 5377–5387. [Google Scholar] [CrossRef] [Green Version]
- Debyser, W.; Schooneveld-Bergmans, M.E.F.; Derdelinckx, G.; Grobet, P.J.; Delcour, J.A. Nuclear magnetic resonance and methylation analysis-derived structural features of water-extractable arabinoxylans from barley (Hordeum vulgare L.) malts. J. Agric. Food Chem. 1997, 45, 2914–2918. [Google Scholar] [CrossRef]
- Mechelke, M.; Koeck, D.E.; Broeker, J.; Roessler, B.; Krabichler, F.; Schwarz, W.H.; Zverlov, V.V.; Liebl, W. Characterization of the arabinoxylan-degrading machinery of the thermophilic bacterium Herbinix hemicellulosilytica—Six new xylanases, three arabinofuranosidases and one xylosidase. J. Biotechnol. 2017, 257, 122–130. [Google Scholar] [CrossRef]
- Thakur, A.; Sharma, K.; Goyal, A. α-L-Arabinofuranosidase: A potential enzyme for the food industry. In Green Bio-Processes Enzymes in Industrial Food Processing; Agarwal, A.K., Pandey, A., Eds.; Springer: Singapore, 2019; pp. 229–244. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate (1.0%, w/v) | Relative Activity (%) 1 |
|---|---|
| arabinoxylo-oligosaccharide | 100 |
| Oat-spelt xylan | 44.1 ± 0.3 |
| Soluble wheat arabinoxylan | 87.5 ± 0.6 |
| Insoluble wheat arabinoxylan | 7.8 ± 0.05 |
| Birchwood xylan | 0 |
| Beechwood xylan | 0 |
| Variable | Value |
|---|---|
| Moisture (%) | 4.4 ± 0.2 |
| Color (EBC) | 4.33 ± 0.14 |
| Turbidity (EBC) | 1.36 ± 0.03 |
| Free amino nitrogen (mg/L) | 182 ± 2 |
| Extract (%) | 79.1 ± 0.1 |
| Total protein (%) | 12.85 ± 0.21 |
| Kolbach index (%) | 44.6 ± 0.4 |
| β-glucan (mg/L) | 96 ± 3 |
| Filtration rate (mL/30min) | 150 ± 7 |
| Arabinoxylan polymer content (mg/L) | 303 ± 5 |
| Friability (%) | 74.2 ± 1.2 |
| Viscosity (mPa·s) | 1.51 ± 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Xu, F.; Lu, J. A Glycoside Hydrolase Family 62 A-L-Arabinofuranosidase from Trichoderma Reesei and Its Applicable Potential during Mashing. Foods 2020, 9, 356. https://doi.org/10.3390/foods9030356
Sun J, Xu F, Lu J. A Glycoside Hydrolase Family 62 A-L-Arabinofuranosidase from Trichoderma Reesei and Its Applicable Potential during Mashing. Foods. 2020; 9(3):356. https://doi.org/10.3390/foods9030356
Chicago/Turabian StyleSun, Junyong, Feng Xu, and Jian Lu. 2020. "A Glycoside Hydrolase Family 62 A-L-Arabinofuranosidase from Trichoderma Reesei and Its Applicable Potential during Mashing" Foods 9, no. 3: 356. https://doi.org/10.3390/foods9030356
APA StyleSun, J., Xu, F., & Lu, J. (2020). A Glycoside Hydrolase Family 62 A-L-Arabinofuranosidase from Trichoderma Reesei and Its Applicable Potential during Mashing. Foods, 9(3), 356. https://doi.org/10.3390/foods9030356