Liquid Chromatography Mass Spectrometry Quantification of α-solanine, α-chaconine, and Solanidine in Potato Protein Isolates

 , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Potato Protein Samples
2.3. Sample Preparation
2.4. Liquid Chromatography–Electrospray Ionization Mass Spectrometry Analysis
2.5. Data Analysis
3. Results and discussion
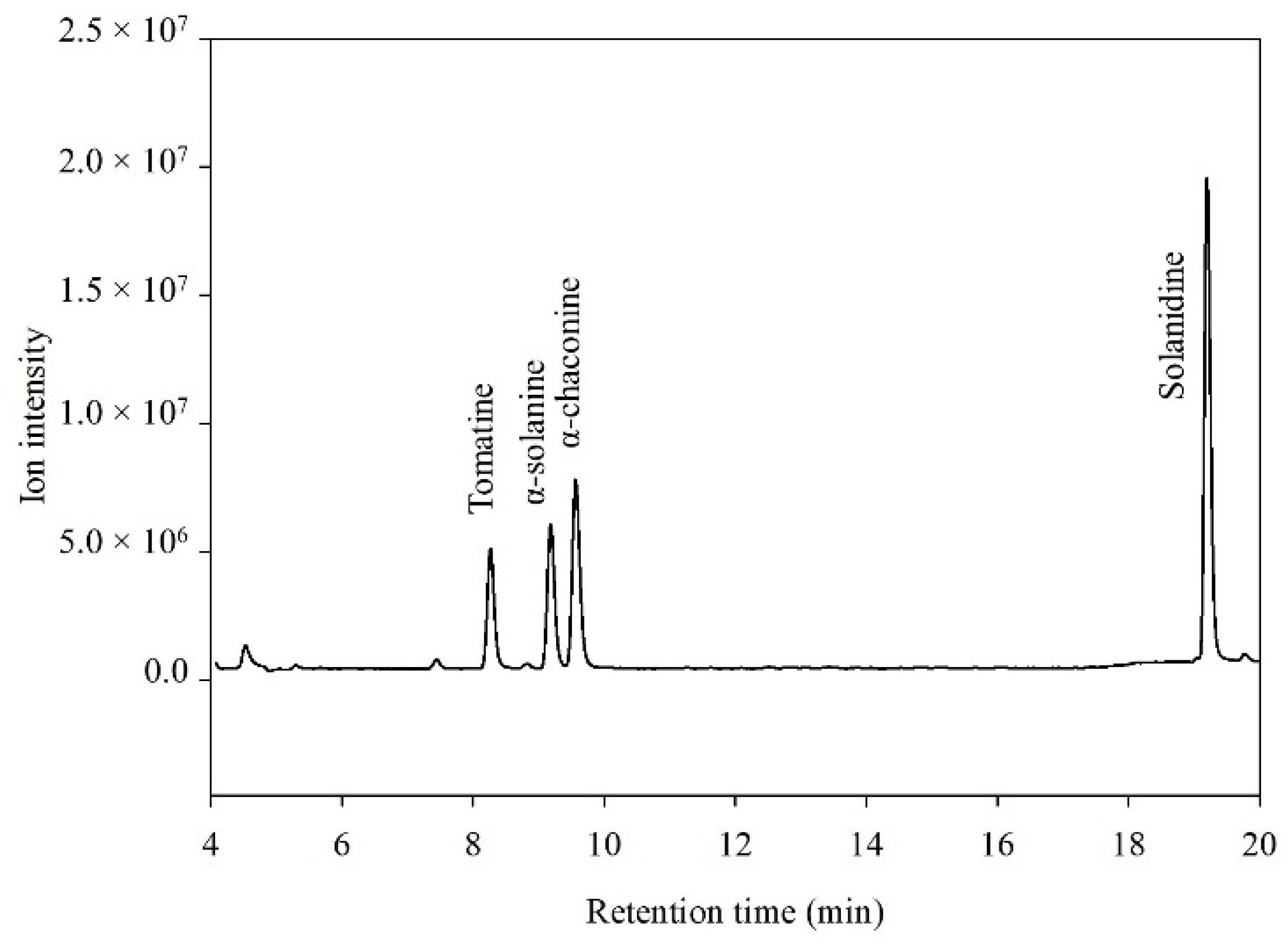
3.1. Liquid Chromatography–Electrospray Ionization Mass Spectrometry Single Quadrupole Analysis
3.2. Glycoalkaloids in Potato Protein Isolates
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knorr, D.; Kohler, G.O.; Betschart, A.A. Potato protein concentrates: The influence of various methods of recovery upon yield, compositional and functional characteristics. J. Food Process. Preserv. 1977, 1, 235–247. [Google Scholar] [CrossRef]
- Pouvreau, L.; Gruppen, H.; Piersma, S.R.; van den Broek, L.A.; van Koningsveld, G.A.; Voragen, A.G. Relative abundance and inhibitory distribution of protease inhibitors in potato juice from cv. Elkana. J. Agric. Food Chem. 2001, 49, 2864–2874. [Google Scholar] [CrossRef]
- Alting, A.C.; Pouvreau, L.; Giuseppin, K.L.F.; van Nieuwenhuijzen, N.H. Potato proteins. In Handbook of Food Proteins; Phillips, G.O., Williamn, P.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 316–334. [Google Scholar]
- Schmidt, J.M.; Damgaard, H.; Greve-Poulsen, M.; Larsen, L.B.; Hammershøj, M. Foam and emulsion properties of potato protein isolate and purified fractions. Food Hydrocoll. 2018, 74, 367–378. [Google Scholar] [CrossRef]
- Sinden, S.L.; Deahl, K.L.; Aulenbach, B.B. Effect of glycoalkaloids and phenolics on potato flavor. J. Food Sci. 1976, 41, 520–523. [Google Scholar] [CrossRef]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef]
- Lachman, J.; Hamouz, K.; Orsak, M.; Pivec, V. Potato glycoalkaloids and their significance in plant protection and human nutrition—Review. Rostl. Vyroba 2001, 47, 181–191. [Google Scholar]
- Kasnak, C.; Artik, N. Change in some glycoalkaloids of potato under different storage regimes. Potato Res. 2018, 61, 183–193. [Google Scholar] [CrossRef]
- McGehee, D.S.; Krasowski, M.D.; Fung, D.L.; Wilson, B.; Gronert, G.A.; Moss, J. Cholinesterase inhibition by potato glycoalkaloids slows mivacurium metabolism. Anesthesiology 2000, 93, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Nigg, H.N.; Ramos, L.E.; Graham, E.M.; Sterling, J.; Brown, S.; Cornell, J.A. Inhibition of human plasma and serum butyrylcholinesterase (ec 3.1.1.8) by alpha-chaconine and alpha-solanine. Fundam. Appl. Toxicol. Off. J. Soc. Toxicol. 1996, 33, 272–281. [Google Scholar]
- The Commission of the European Communities. 2002/150/EC, Commission Decision of 15 February 2002 Authorising the Placing on the Market of Coagulated Potato Proteins and Hydrolysates Thereof as Novel Food Ingredients under Regulation (ec) no 258/97 of the European Parliament and of the Council (Notified under Document Number C(2002) 506). 2002, OJ L 50, 21.2.2002, 92–93. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32002D0150 (accessed on 29 March 2020).
- Lokra, S.; Straetkvern, K.O. Industrial Proteins from Potato Juice. A Review. Food 2009, 3, 88–95. [Google Scholar]
- Laus, M.C.; Klip, G.; Giuseppin, M.L.F. Improved extraction and sample cleanup of tri-glycoalkaloids alpha-solanine and alpha-chaconine in non-denatured potato protein isolates. Food. Anal. Meth. 2017, 10, 845–853. [Google Scholar] [CrossRef]
- Simonovska, B.; Vovk, I. High-performance thin-layer chromatographic determination of potato glycoalkaloids. J. Chromatogr. A 2000, 903, 219–225. [Google Scholar] [CrossRef]
- Stanker, L.H.; Kamps-Holtzapple, C.; Friedman, M. Development and characterization of monoclonal antibodies that differentiate between potato and tomato glycoalkaloids and aglycons. J. Agric. Food Chem. 1994, 42, 2360–2366. [Google Scholar] [CrossRef]
- Driedger, D.R.; Sporns, P. Glycoalkaloid concentration in by-products of potato starch extraction as measured by matrix-assisted laser desorptioniionization mass spectrometry. J. Food Process. Preserv. 1999, 23, 377–390. [Google Scholar] [CrossRef]
- Horwitz, W. AOAC official method 997.13, Glycoalkaloids (α-solanine and α-chaconine) in potato tubers. AOAC Off. Methods Anal. 2000, 49, 62–64. [Google Scholar]
- Vaananen, T.; Ikonen, T.; Rokka, V.M.; Kuronen, P.; Serimaa, R.; Ollilainen, V. Influence of incorporated wild solanum genomes on potato properties in terms of starch nanostructure and glycoalkaloid content. J. Agric. Food Chem. 2005, 53, 5313–5325. [Google Scholar] [CrossRef] [PubMed]
- Lachman, J.; Hamouz, K.; Musilova, J.; Hejtmankova, K.; Kotikova, Z.; Pazderu, K.; Domkarova, J.; Pivec, V.; Cimr, J. Effect of peeling and three cooking methods on the content of selected phytochemicals in potato tubers with various colour of flesh. Food chem. 2013, 138, 1189–1197. [Google Scholar] [CrossRef]
- Bártová, V.; Bárta, J.; Brabcová, A.; Zdráhal, Z.; Horáčková, V. Amino acid composition and nutritional value of four cultivated south american potato species. J. Food Compos. Anal. 2015, 40, 78–85. [Google Scholar] [CrossRef]
- Kuronen, P.; Vaananen, T.; Pehu, E. Reversed-phase liquid chromatographic separation and simultaneous profiling of steroidal glycoalkaloids and their aglycones. J. Chromatogr. A 1999, 863, 25–35. [Google Scholar] [CrossRef]
- Schmidt, J.M.; Greve-Poulsen, M.; Damgaard, H.; Sunds, A.V.; Zdrahal, Z.; Hammershoj, M.; Larsen, L.B. A new two-step chromatographic procedure for fractionation of potato proteins with potato fruit juice and spray-dried protein as source materials. Food Bioprocess Technol. 2017, 10, 1946–1958. [Google Scholar] [CrossRef]
- Alt, V.; Steinhof, R.; Lotz, M.; Ulber, R.; Kasper, C.; Scheper, T. Optimization of glycoalkaloid analysis for use in industrial potato fruit juice downstreaming. Eng. Life Sci. 2005, 5, 562–567. [Google Scholar] [CrossRef]
- Friedman, M.; Roitman, J.N.; Kozukue, N. Glycoalkaloid and calystegine contents of eight potato cultivars. J. Agric. Food Chem. 2003, 51, 2964–2973. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; McDonald, G.; Haddon, W.F. Kinetics of acid-catalyzed hydrolysis of carbohydrate groups of potato glycoalkaloids alpha-chaconine and alpha-solanine. J. Agric. Food Chem. 1993, 41, 1397–1406. [Google Scholar] [CrossRef]
- Takagi, K.; Toyoda, M.; Fujiyama, Y.; Saito, Y. Effect of cooking on contents of α-chaconine and α-solanine in potatoes. Shokuhin Eiseigaku Zasshi 1990, 31, 67–73. [Google Scholar] [CrossRef]
- Jensen, P.H.; Pedersen, R.B.; Svensmark, B.; Strobel, B.W.; Jacobsen, O.S.; Hansen, H.C. Degradation of the potato glycoalkaloid alpha-solanine in three agricultural soils. Chemosphere 2009, 76, 1150–1155. [Google Scholar] [CrossRef]
- Giuseppin, M.L.F.; Spelbrink, R.E.J. Glyckoalkaloid Removal. WO 2008/069651 A1, 12 June 2008. [Google Scholar]
- Marchal, M.L.J.; Nijssen, J.M.H.; Knott, R.E.; Krol, R.A.F. Method for Improving a Protein Product. WO 02/100187 A1, 19 December 2002. [Google Scholar]
- Backleh, M.; Ekici, P.; Leupold, G.; Coelhan, M.; Parlar, H. Enrichment of the glycoalkaloids alpha-solanine and alpha-chaconine from potato juice by adsorptive bubble separation using a ph gradient. J. Sep. Sci. 2004, 27, 1042–1044. [Google Scholar] [CrossRef]
- Ralla, K.; Sohling, U.; Suck, K.; Kasper, C.; Ruf, F.; Scheper, T. Separation of patatins and protease inhibitors from potato fruit juice with clay minerals as cation exchangers. J. Sep. Sci. 2012, 35, 1596–1602. [Google Scholar] [CrossRef]
- Friedman, M.; Rayburn, J.R.; Bantle, J.A. Developmental toxicology of potato alkaloids in the frog embryo teratogenesis assay xenopus (fetax). Food Chem. Toxicol. 1991, 29, 537–547. [Google Scholar] [CrossRef]
- Rayburn, J.R.; Bantle, J.A.; Friedman, M. Role of carbohydrate side-chains of potato glycoalkaloids in developmental toxicity. J. Agric. Food Chem. 1994, 42, 1511–1515. [Google Scholar] [CrossRef]
- Zhao, J.X.; Camire, M.E.; Bushway, R.J.; Bushway, A.A. Glycoalkaloid content and in-vitro glycoalkaloid solubility of extruded potato peels. J. Agric. Food Chem. 1994, 42, 2570–2573. [Google Scholar] [CrossRef]
- Hennessy, R.C.; Nielsen, S.D.; Greve-Poulsen, M.; Larsen, L.B.; Sørensen, O.B.; Stougaard, P. Discovery of a bacterial gene cluster for deglycosylation of toxic potato steroidal glycoalkaloids α-chaconine and α-solanine. J. Agric. Food Chem. 2020, 68, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Hellenas, K.E.; Nyman, A.; Slanina, P.; Loof, L.; Gabrielsson, J. Determination of potato glycoalkaloids and their aglycone in blood-serum by high-performance liquid-chromatography—Application to pharmacokinetic studies in humans. J. Chromatogr. Biomed. Appl. 1992, 573, 69–78. [Google Scholar] [CrossRef]
- Mensinga, T.T.; Sips, A.; Rompelberg, C.J.M.; van Twillert, K.; Meulenbelt, J.; van den Top, H.J.; van Egmond, H.P. Potato glycoalkaloids and adverse effects in humans: An ascending dose study. Regul. Toxicol. Pharmacol. 2005, 41, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Dalvi, R.R.; Bowie, W.C. Toxicology of solanine—An overview. Vet. Hum. Toxicol. 1983, 25, 13–15. [Google Scholar]
- Yamashoji, S.; Matsuda, T. Synergistic cytotoxicity induced by alpha-solanine and alpha-chaconine. Food Chem. 2013, 141, 669–674. [Google Scholar] [CrossRef]
- Rayburn, J.R.; Friedman, M.; Bantle, J.A. Synergistic interaction of glycoalkaloids alpha-chaconine and alpha-solanine on developmental toxicity in xenopus embryos. Food Chem. Toxicol. 1995, 33, 1013–1019. [Google Scholar] [CrossRef]
- Langkilde, S.; Mandimika, T.; Schroder, M.; Meyer, O.; Slob, W.; Peijnenburg, A.; Poulsen, M. A 28-day repeat dose toxicity study of steroidal glycoalkaloids, alpha-solanine and alpha-chaconine in the syrian golden hamster. Food Chem. Toxicol. 2009, 47, 1099–1108. [Google Scholar] [CrossRef]

 ) and qualifier ions in single ion monitoring mode (
) and qualifier ions in single ion monitoring mode (  ).
) and qualifier ions in single ion monitoring mode ( ).
).
) and qualifier ions in single ion monitoring mode ( ).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | [M+H]+ | Ions (m/z) | Peak usage |
|---|---|---|---|
| α-solanine | 868.4 | 868.4 | Target Ion |
| 560.4 | Qualifier Ion | ||
| 398.2 | Qualifier Ion | ||
| α-chaconine | 852.4 | 852.4 | Target Ion |
| 560.4 | Qualifier Ion | ||
| 398.2 | Qualifier Ion | ||
| Solanidine | 398.2 | 398.2 | Target Ion |
| Tomatine (ISTD) | 1034.4 | 1034.4 | Qualifier Ion |
| 576.4 | Target Ion | ||
| 416.2 | Qualifier Ion |
| Compound | Linearity (R2) | LOD (µg mL−1) | LOQ (µg mL−1) | Recovery | |
|---|---|---|---|---|---|
| 0.06 µg mL−1 | 2 µg mL−1 | ||||
| α-Solanine | 0.994 | 0.012 | 0.03 | 81.6% | 106.4% |
| α-Chaconine | 0.996 | 0.011 | 0.03 | 82.7% | 101.5% |
| Solanidine | 0.997 | 0.003 | 0.01 | 90.3% | 83.9% a |
| Concentration (µg mL−1) | Intra-Day Precision (% CV, n = 6) | Intra-Day Accuracy (%, n = 6) | Inter-Day Precision (% CV, n = 4) | Inter-Day Accuracy (%, n = 4) |
|---|---|---|---|---|
| α-Solanine | ||||
| 0.03 | 8.07 | 112.41 | 1.76 | 128.81 |
| 0.1 | 8.15 | 104.01 | 0.70 | 108.21 |
| 0.3 | 8.58 | 103.26 | 0.12 | 105.62 |
| 1 | 8.04 | 88.41 | 6.93 | 85.88 |
| 3 | 5.62 | 101.24 | 6.77 | 101.46 |
| α-Chaconine | ||||
| 0.03 | 7.04 | 114.33 | 8.89 | 79.93 |
| 0.1 | 5.26 | 99.68 | 10.53 | 89.34 |
| 0.3 | 7.12 | 101.80 | 8.47 | 107.85 |
| 1 | 6.45 | 82.26 | 10.71 | 99.42 |
| 3 | 4.26 | 95.76 | 4.90 | 107.96 |
| Solanidine | ||||
| 0.01 | 2.96 | 85.97 | 5.49 | 89.93 |
| 0.03 | 4.30 | 87.48 | 6.14 | 83.41 |
| 0.1 | 4.50 | 86.38 | 4.23 | 102.35 |
| 0.3 | 4.74 | 111.89 | 2.92 | 106.34 |
| 1 | 5.98 | 99.09 | 0.65 | 99.43 |
| α-Solanine | α-Chaconine | Solanidine | ||||||
|---|---|---|---|---|---|---|---|---|
| Sample | Treatment | Total GA | µg g−1 (ppm) | %CV | µg g−1 (ppm) | %CV | µg g−1 (ppm) | %CV |
| Feed-grade protein | Early a | 2061.7 ± 94.9 | 782.7 ± 40.0 | 5.1 | 1279.0 ± 54.9 | 9.8 | 38.1 ± 4.4 | 11.5 |
| Mid a | 2456.8 ± 47.8 * | 905.0 ± 23.3 * | 2.57 | 1551.82 ± 24.5 * | 1.6 | 95.6 ± 7.4 * | 7.7 | |
| Late a | 3188.5 ± 303.0 *** | 1129.0±101.9 *** | 9.0 | 2059.5 ± 201.1 *** | 9.8 | 161.2 ± 26.6 *** | 16.5 | |
| Food-grade protein b | High b | 316.0 ± 41.3 | 118.4 ± 16.7 | 14.1 | 197.6± 24.6 | 10.4 | 131.3 ± 21.4 | 16.4 |
| Low b | 28.9 ± 2.3 *** | 10.9 ± 1.0 *** | 8.7 | 18.0± 1.3 *** | 7.4 | 41.9 ± 7.4 *** | 15.7 | |
| PIc fraction | IEX chromatography d | 641.1 ± 17.9 | 606.0 ± 16.3 | 2.7 | 35.1 ± 1.6 | 4.6 | 549.3 ± 100.3 | 18.3 |
| Patatin fraction | IEX chromatography d | 50.6 ± 3.5 *** | 43.2 ± 3.4 *** | 7.8 | 7.4 ± 0.1 *** | 1.2 | 69.6 ± 4.6 *** | 6.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, S.D.; Schmidt, J.M.; Kristiansen, G.H.; Dalsgaard, T.K.; Larsen, L.B. Liquid Chromatography Mass Spectrometry Quantification of α-solanine, α-chaconine, and Solanidine in Potato Protein Isolates. Foods 2020, 9, 416. https://doi.org/10.3390/foods9040416
Nielsen SD, Schmidt JM, Kristiansen GH, Dalsgaard TK, Larsen LB. Liquid Chromatography Mass Spectrometry Quantification of α-solanine, α-chaconine, and Solanidine in Potato Protein Isolates. Foods. 2020; 9(4):416. https://doi.org/10.3390/foods9040416
Chicago/Turabian StyleNielsen, Søren D., Jesper M. Schmidt, Gitte H. Kristiansen, Trine K. Dalsgaard, and Lotte B. Larsen. 2020. "Liquid Chromatography Mass Spectrometry Quantification of α-solanine, α-chaconine, and Solanidine in Potato Protein Isolates" Foods 9, no. 4: 416. https://doi.org/10.3390/foods9040416
APA StyleNielsen, S. D., Schmidt, J. M., Kristiansen, G. H., Dalsgaard, T. K., & Larsen, L. B. (2020). Liquid Chromatography Mass Spectrometry Quantification of α-solanine, α-chaconine, and Solanidine in Potato Protein Isolates. Foods, 9(4), 416. https://doi.org/10.3390/foods9040416