Changes in the Content and Bioavailability of Onion Quercetin and Grape Resveratrol During In Vitro Human Digestion


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Sample and Extracts
2.3. In Vitro Digestion System and Digestibility in Small Intestine
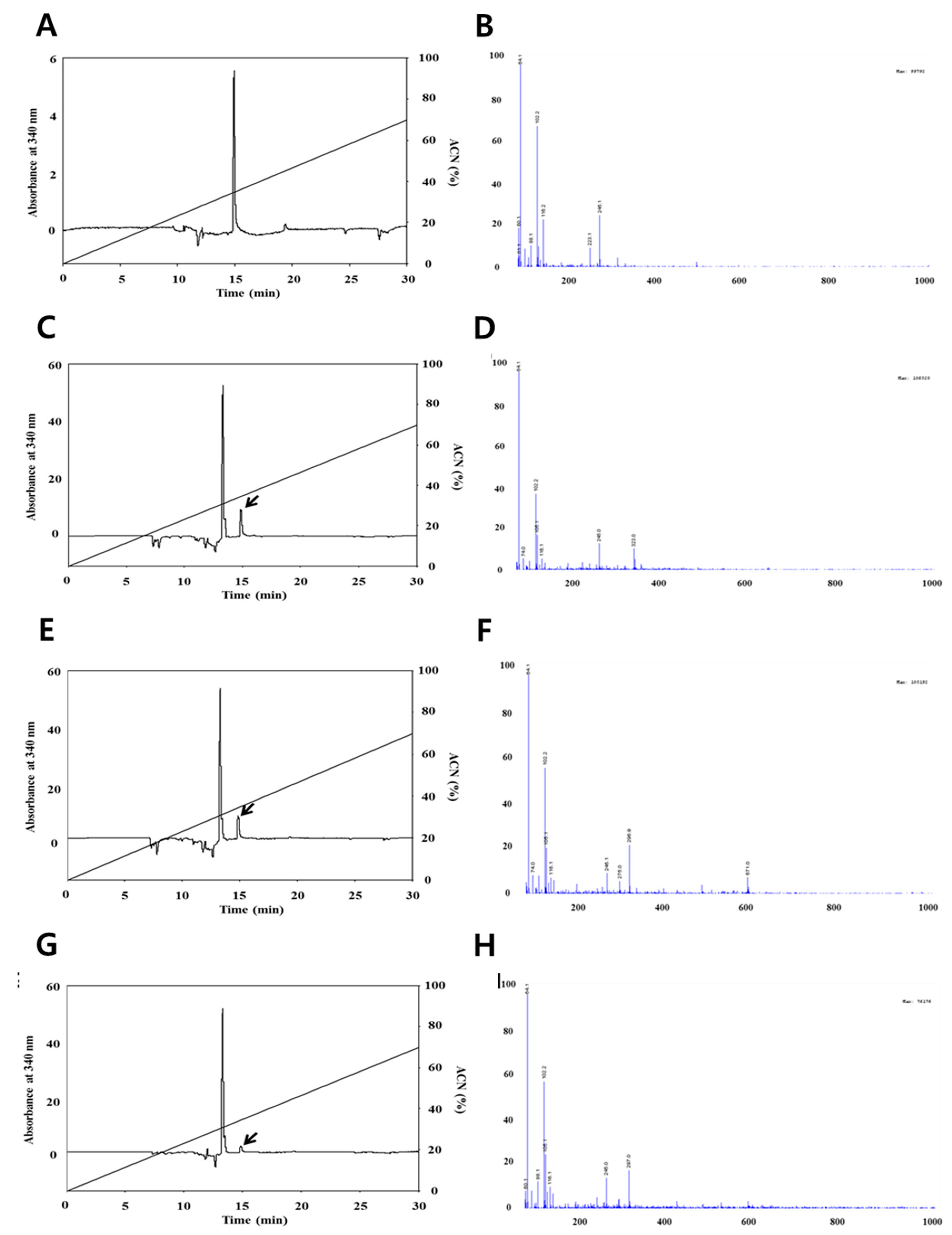
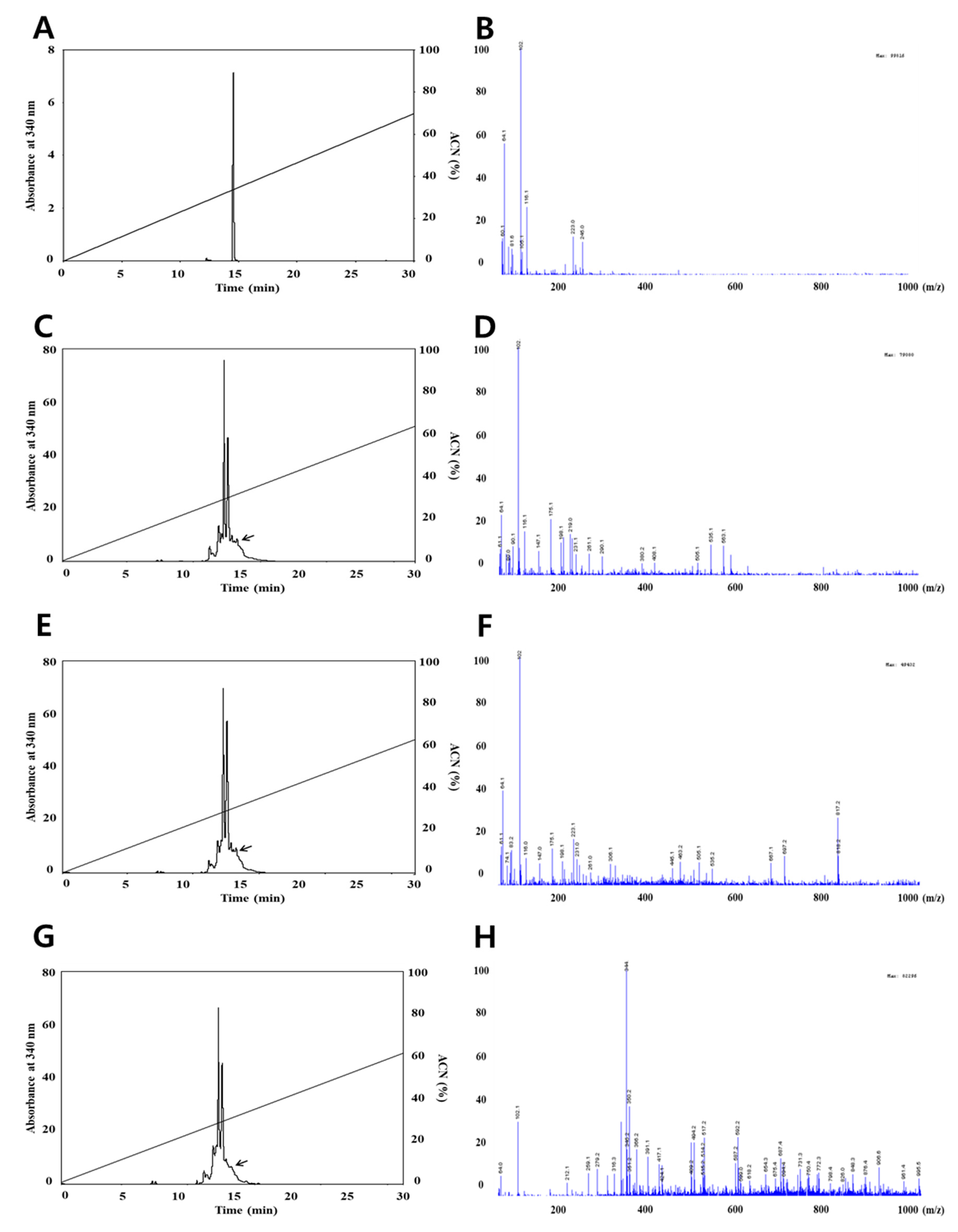
2.4. Measurement of Quercetin and Resveratrol by High-Performance Liquid Chromatography (HPLC)
2.5. Liquid Chromatography-Mass Spectroscopy (LC-MS) Analysis
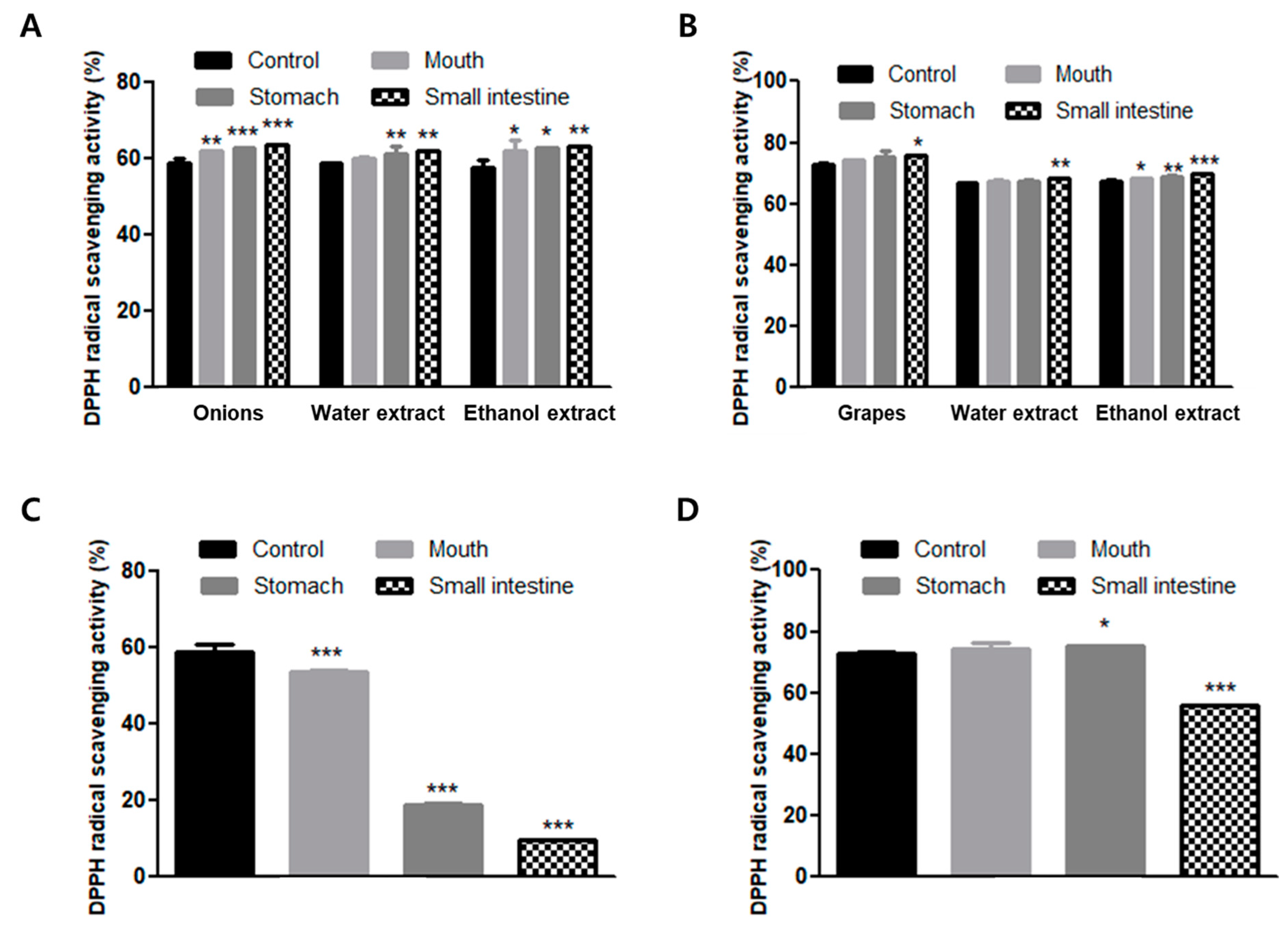
2.6. DPPH Radical Scavenging Activity
2.7. Statistical Analysis
3. Results and Discussion
3.1. Identification of Quercetin and Resveratrol
3.2. Changes in Contents and Digestibility of Onion Quercetin and Grape Resveratrol during In Vitro Human Digestion
3.3. DPPH Radical Scavenging Activity during In Vitro Human Digestion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Croft, K.D.; Zhang, D.; Jiang, R.; Ayer, A.; Shengule, S.; Payne, R.J.; Ward, N.C.; Stocker, R. Structural requirements of flavonoids to induce heme oxygenase-1 expression. Free Radic. Biol. Med. 2017, 113, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, W.; Yang, L.; Ji, Z.; Zhao, Y.; Wang, P.; Zhang, L. Development of electro-hydraulic proportion control system of track-laying machinery for high speed railway construction. Mechatronics 2016, 40, 167–177. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Ting, Y.; Chang, W.T.; Shiau, D.K.; Chou, P.H.; Wu, M.F.; Hsu, C.L. Antiobesity efficacy of quercetin-rich supplement on diet-induced obese rats: Effects on body composition, serum lipid profile, and gene expression. J. Agric. Food Chem. 2018, 66, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yao, J.; Zhou, B.; Yang, J.; Chaudry, M.T.; Wang, M.; Xiao, F.; Li, Y.; Yin, W. Bacteriostatic effect of quercetin as an antibiotic alternative in vivo and its antibacterial mechanism in vitro. J. Food Prot. 2017, 81, 68–78. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Delarami Far, A.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G. Anticancer and apoptosis-inducing effects of quercetin in vitro and in vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Ko, E.Y.; Nile, S.H.; Jung, Y.S.; Keum, Y.S. Antioxidant and antiplatelet potential of different methanol fractions and flavonols extracted from onion (Allium cepa L.). 3 Biotech 2018, 8, 155. [Google Scholar] [CrossRef]
- Grzelak-Błaszczyk, K.; Milala, J.; Kosmala, M.; Kołodziejczyk, K.; Sójka, M.; Czarnecki, A.; Klewicki, R.; Juśkiewicz, J.; Fotschki, B.; Jurgoński, A. Onion quercetin monoglycosides alter microbial activity and increase antioxidant capacity. J. Nutr. Biochem. 2018, 56, 81–88. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1, 35. [Google Scholar] [CrossRef] [Green Version]
- Soleymani, S.; Iranpanah, A.; Najafi, F.; Belwal, T.; Ramola, S.; Abbasabadi, Z.; Momtaz, S.; Farzaei, M.H. Implications of grape extract and its nanoformulated bioactive agent resveratrol against skin disorders. Arch. Dermatol. Res. 2019, 311, 577–588. [Google Scholar] [CrossRef]
- Chen, T.S.; Ju, D.T.; Day, C.H.; Yeh, Y.L.; Chen, R.J.; Viswanadha, V.P.; Chang, R.L.; Lin, Y.C.; Yao, C.H.; Huang, C.Y. Protective effect of autologous transplantation of resveratrol preconditioned adipose-derived stem cells in the treatment of diabetic liver dysfunction in rat model. J. Tissue Eng. Regen. Med. 2019, 13, 1629–1640. [Google Scholar] [CrossRef] [PubMed]
- Petrella, C.; Carito, V.; Carere, C.; Ferraguti, G.; Ciafrè, S.; Natella, F.; Bello, C.; Greco, A.; Ralli, M.; Mancinelli, R.; et al. Oxidative stress inhibition by resveratrol in alcohol dependent mice. Nutrition 2020, 2020, 110783. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food. Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef] [PubMed]
- De Sá Coutinho, D.; Pacheco, M.T.; Frozza, R.L.; Bernardi, A. Anti-inflammatory effects of resveratrol: Mechanistic insights. Int. J. Mol. Sci. 2018, 19, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.; Kroon, P.A.; Das, D.K.; Das, S.; Tosaki, A.; Chan, V.; Singer, M.V.; Feick, P. The biological responses to resveratrol and other polyphenols from alcoholic beverages. Alcohol.Clin. Exp. Res. 2009, 33, 1513–1523. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef]
- Chen, H.Y.; Lin, P.H.; Shih, Y.H.; Wang, K.L.; Hong, Y.H.; Shieh, T.M.; Huang, T.C.; Hsia, S.M. Natural antioxidant resveratrol suppresses uterine fibroid cell growth and extracellular matrix formation in vitro and in vivo. Antioxidants 2019, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, C.; Iezzi, M.; Ciolli, L.; Hysi, A.; Bucci, I.; Di Santo, S.; Rossi, C.; Zucchelli, M.; Napolitano, G. Resveratrol has anti-thyroid effects both in vitro and in vivo. Food Chem. Toxicol. 2017, 107, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, M.; Chen, Y.; Wang, X.J.; Tang, X. Resveratrol: An overview of its anti-cancer mechanisms. Life Sci. 2018, 207, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, S.Y.; Chung, M.S.; Hur, S.J. Development of novel in vitro human digestion systems for screening the bioavailability and digestibility of foods. J. Funct. Foods 2016, 22, 113–121. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Lee, S.J. Effects of biopolymers encapsulations on the lipid digestibility of emulsion-type sausages using a simulated human gastrointestinal digestion model. Food Bioprocess Technol. 2014, 7, 2198–2206. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Gumienna, M.; Lasik, M.; Czarnecki, Z. Bioconversion of grape and chokeberry wine polyphenols during simulated gastrointestinal in vitro digestion. Int. J. Food Sci. Nutr. 2011, 62, 226–233. [Google Scholar] [CrossRef]
- Gao, L.; Liu, G.; Wang, X.; Liu, F.; Xu, Y.; Ma, J. Preparation of a chemically stable quercetin formulation using nanosuspension technology. Int. J. Pharm. 2011, 404, 231–237. [Google Scholar] [CrossRef]
- Priprem, A.; Watanatorn, J.; Sutthiparinyanont, S.; Phachonpai, W.; Muchimapura, S. Anxiety and cognitive effects of quercetin liposomes in rats. Nanomedicine 2008, 4, 70–78. [Google Scholar] [CrossRef]
- Kwak, J.H.; Seo, J.M.; Kim, N.H.; Arasu, M.V.; Kim, S.; Yoon, M.K.; Kim, S.J. Variation of quercetin glycoside derivatives in three onion (Allium cepa L.) varieties. Saudi J. Biol. Sci. 2017, 24, 1387–1391. [Google Scholar] [CrossRef] [Green Version]
- Mullen, W.; Rouanet, J.M.; Auger, C.; Teissedre, P.L.; Caldwell, S.T.; Hartley, R.C.; Lean, M.E.; Edwards, C.A.; Crozier, A. Bioavailability of [2-14C] quercetin-4′-glucoside in rats. J. Agric. Food Chem. 2008, 56, 12127–12137. [Google Scholar] [CrossRef]
- Walle, T. Absorption and metabolism of flavonoids. Free Radic. Biol. Med. 2004, 36, 829–837. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.; Kim, D.; Chun, S.; Lee, S. Onion extract structural changes during in vitro digestion and its potential antioxidant effect on brain lipids obtained from low-and high-fat-fed mice. Free Radic. Res. 2013, 47, 1009–1015. [Google Scholar] [CrossRef]
- Hur, S.J.; Park, S.J.; Jeong, C.H. Effect of buckwheat extract on the antioxidant activity of lipid in mouse brain and its structural change during in vitro human digestion. J. Agric. Food Chem. 2011, 59, 10699–10704. [Google Scholar] [CrossRef]
- Boyer, J.; Brown, D.; Liu, R.H. In vitro digestion and lactase treatment influence uptake of quercetin and quercetin glucoside by the Caco-2 cell monolayer. Nutr. J. 2005, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Zupančič, Š.; Lavrič, Z.; Kristl, J. Stability and solubility of trans-resveratrol are strongly influenced by pH and temperature. Eur. J. Pharm. Biopharm. 2015, 93, 196–204. [Google Scholar] [CrossRef]
- Sessa, M.; Tsao, R.; Liu, R.; Ferrari, G.; Donsì, F. Evaluation of the stability and antioxidant activity of nanoencapsulated resveratrol during in vitro digestion. J. Agric. Food Chem. 2011, 59, 12352–12360. [Google Scholar] [CrossRef]
- Francioso, A.; Mastromarino, P.; Masci, A.; d’Erme, M.; Mosca, L. Chemistry, stability and bioavailability of resveratrol. Med. Chem. 2014, 10, 237–245. [Google Scholar] [CrossRef]
- Bermúdez-Soto, M.J.; Tomás-Barberán, F.A.; García-Conesa, M.T. Stability of polyphenols in chokeberry (Aronia melanocarpa) subjected to in vitro gastric and pancreatic digestion. Food Chem. 2007, 102, 865–874. [Google Scholar] [CrossRef]
- Duarte, A.; Martinho, A.; Luís, Â.; Figueiras, A.; Oleastro, M.; Domingues, F.C.; Silva, F. Resveratrol encapsulation with methyl-β-cyclodextrin for antibacterial and antioxidant delivery applications. LWT-Food Sci. Technol. 2015, 63, 1254–1260. [Google Scholar] [CrossRef]
- Tavares, L.; Figueira, I.; Macedo, D.; McDougall, G.J.; Leitão, M.C.; Vieira, H.L.A.; Stewart, D.; Alves, P.M.; Ferreira, R.B.; Santos, C.N. Neuroprotective effect of blackberry (Rubus sp.) polyphenols is potentiated after simulated gastrointestinal digestion. Food Chem. 2012, 131, 1443–1452. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Smith, G.E. In vitro binding of bile acids by blueberries (Vaccinium spp.), plums (Prunus spp.), prunes (Prunus spp.), strawberries (Fragaria X ananassa), cherries (Malpighia punicifolia), cranberries (Vaccinium macrocarpon) and apples (Malus sylvestris). Food Chem. 2007, 100, 1182–1187. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Saliva | Gastric Juice | Duodenal Juice | Bile Juice | |
|---|---|---|---|---|
| Organic and inorganic components | 1.7 mL NaCl a (175.3 g/L) b | 6.5 mL HCl (37 g/L) | 6.3 mL KCl (89.6 g/L) | 68.3 mL NaHCO3 (84.7 g/L) |
| 8 mL urea (25 g/L) | 18 mL CaCl2·2H2O (22.2 g/L) | 9 mL CaCl2·2H2O (22.2 g/L) | 10 mL CaCl2·2H2O (22.2 g/L) | |
| 15 mg uric acid | 1 g bovine serum albumin | 1 g bovine serum albumin | 1.8 g bovine serum albumin | |
| 30 g bile | ||||
| Enzymes | 290 mg α-amylase | 2.5 g pepsin | 9 g pancreatin | |
| 25 mg mucin | 3 g mucin | 1.5 g lipase | ||
| pH | 6.8 ± 0.2 | 1.50 ± 0.02 | 8.0 ± 0.2 | 7.0 ± 0.2 |
| Quercetin (mg)/Onion (g) | Resveratrol (mg/)/Grape (g) | ||
|---|---|---|---|
| Homogenized | Before digestion | 5.23 ± 0.18 a | 1.87 ± 0.64 a |
| Mouth | 5.05 ± 0.22 a | 1.56 ± 0.51 a | |
| Stomach | 4.77 ± 0.11 a | 1.54 ± 0.86 a | |
| Small intestine | 3.90 ± 0.26 b | 0.89 ± 0.47 a | |
| Water extract | Before digestion | 5.22 ± 0.21 a | 1.47 ± 0.26 a |
| Mouth | 4.41 ± 0.30 ab | 1.40 ± 0.47 a | |
| Stomach | 4.37 ± 0.44 b | 1.15 ± 0.54 a | |
| Small intestine | 4.18 ± 0.27 b | 1.09 ± 0.65 a | |
| Ethanol extract | Before digestion | 6.02 ± 0.28 a | 1.97 ± 0.71 a |
| Mouth | 5.99 ± 0.17 a | 1.66 ± 0.65 a | |
| Stomach | 5.82 ± 0.29 ab | 1.24 ± 0.48 a | |
| Small intestine | 5.33 ± 0.09 b | 1.09 ± 0.64 a |
| Small Intestine | ||
|---|---|---|
| Quercetin (%) | Resveratrol (%) | |
| Homogenized | 45.68 ± 6.45 | 52.45 ± 3.48 |
| Water extract | 40.14 ± 4.65 | 51.08 ± 5.54 |
| Ethanol extract | 46.48 ± 6.04 | 49.54 ± 3.47 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.Y.; Lee, S.J.; Yim, D.G.; Hur, S.J. Changes in the Content and Bioavailability of Onion Quercetin and Grape Resveratrol During In Vitro Human Digestion. Foods 2020, 9, 694. https://doi.org/10.3390/foods9060694
Lee SY, Lee SJ, Yim DG, Hur SJ. Changes in the Content and Bioavailability of Onion Quercetin and Grape Resveratrol During In Vitro Human Digestion. Foods. 2020; 9(6):694. https://doi.org/10.3390/foods9060694
Chicago/Turabian StyleLee, Seung Yun, Seung Jae Lee, Dong Gyun Yim, and Sun Jin Hur. 2020. "Changes in the Content and Bioavailability of Onion Quercetin and Grape Resveratrol During In Vitro Human Digestion" Foods 9, no. 6: 694. https://doi.org/10.3390/foods9060694
APA StyleLee, S. Y., Lee, S. J., Yim, D. G., & Hur, S. J. (2020). Changes in the Content and Bioavailability of Onion Quercetin and Grape Resveratrol During In Vitro Human Digestion. Foods, 9(6), 694. https://doi.org/10.3390/foods9060694