Bioactive Components and Antioxidant Activity Distribution in Pearling Fractions of Different Greek Barley Cultivars



Abstract
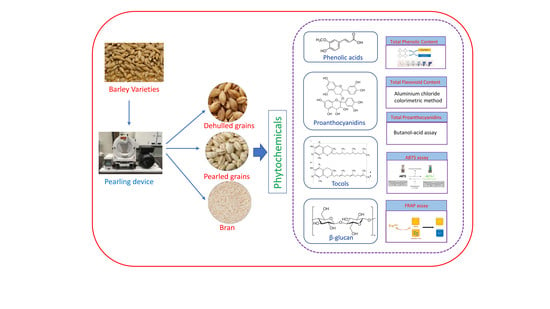
:
1. Introduction
2. Materials and Methods
2.1. Barley Samples and Chemicals
2.2. Grain Characteristics
2.3. Chemical Analyses
2.4. Phenolics Extraction
2.5. Total Phenolic Content (TPC)
2.6. Total Flavonoid Content (TFC)
2.7. Total Proanthocyanidins Content (TPAC)
2.8. ABTS Radical Scavenging Activity
2.9. Ferric Reducing Antioxidant Power (FRAP)
2.10. HPLC Analysis of Phenolic Compounds
2.11. Tocopherols and Tocotrienols Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. Barley Grain Characteristics
3.2. Chemical Characterization of Barley Pearling Fractions
3.3. Phenolic Contents and Antioxidant Activity of Barley Pearling Fractions
3.4. Quantification of Flavanols and Phenolic Acids in Barley Pearling Fractions
3.5. Tocopherols and Tocotrienols in Barley Pearling Fractions
3.6. Antioxidant Activity of Barley Pearling Fractions
3.7. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Baik, B.K.; Ullrich, S.E. Barley for food: Characteristics, improvement, and renewed interest. J. Cereal Sci. 2008, 48, 232–242. [Google Scholar] [CrossRef]
- Panfili, G.; Fratianni, A.; Criscio, T.D.; Marconi, E. Tocol and b-glucan levels in barley cultivars and in pearling by-products. Food Chem. 2008, 107, 84–91. [Google Scholar] [CrossRef]
- Do, T.D.T.; Cozzolino, D.; Muhlhausler, B.; Box, A.; Able, A.J. Antioxidant capacity and vitamin E in barley: Effect of genotype and storage. Food Chem. 2015, 187, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Aal, E.S.M.; Choo, T.M.; Dhillon, S.; Rabalski, I. Free and bound phenolic acids and total phenolics in black, blue, and yellow barley and their contribution to free radical scavenging capacity. Cereal Chem. 2012, 89, 198–204. [Google Scholar] [CrossRef]
- Rao, S.; Santhakumara, A.B.; Chinkwo, K.A.; Blanchard, C.L. Q-TOF LC/MS identification and UHPLC-Online ABTS antioxidant activity guided mapping of barley polyphenols. Food Chem. 2018, 266, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Barrows, F.T.; Obert, D. Dry fractionation methods to produce barley meals varying in protein, beta-glucan, and starch contents. J. Food Sci. 2009, 74, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.; O’Flaherty, J.; Brunton, N.; Gee, V.L.; Arendt, E.; Gallagher, E. Chemical composition and microstructure of milled barley fractions. Eur. Food Res. Technol. 2010, 230, 579–595. [Google Scholar] [CrossRef]
- Gangopadhyay, N.; Harrison, S.B.; Brunton, N.P.; Hidalgo-Ruiz, J.L.; Gallagher, E.; Rai, D.K. Brans of the roller-milled barley fractions rich in polyphenols and health-promoting lipophilic molecules. J. Cereal Sci. 2018, 83, 213–221. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G.; Izydorczyk, M.S. Cereal β-glucans: Structure, physical properties and physiological functions. In Functional Food Carbohydrates; Biliaderis, C.G., Izydorczyk, M.S., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–72. [Google Scholar]
- Holtekjølen, A.K.; Sahlstrøm, S.; Knutsen, S.H. Phenolic contents and antioxidant activities in covered whole-grain flours of Norwegian barley cultivars and in fractions obtained after pearling. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 67–74. [Google Scholar]
- Lazaridou, A.; Biliaderis, C.G. Molecular aspects of cereal β-glucan functionality: Physical properties, technological applications and physiological effects. J. Cereal Sci. 2007, 46, 101–118. [Google Scholar] [CrossRef]
- Gamel, T.H.; Abdel-Aal, E.M. Phenolic acids and antioxidant properties of barley wholegrain and pearling fractions. Agric. Food Sci. 2012, 21, 118–131. [Google Scholar] [CrossRef]
- AACC. Approved Methods of American Association of Cereal Chemists, 10th ed.; American Association of Cereal Chemists Inc: Eagan, MN, USA, 2000. [Google Scholar]
- Irakli, M.; Kleisiaris, F.; Mygdalia, A.; Katsantonis, D. Optimizing extraction conditions of free and bound phenolic compounds from rice by-products and their antioxidant effects. Foods 2018, 73, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar]
- Bao, J.S.; Cai, Y.; Sun, M.; Wang, G.Y.; Corke, H. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1986, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C.A. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, F.; Strain, J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–23. [Google Scholar]
- Skendi, A.; Irakli, M.; Chatzopoulou, P. Analysis of phenolic compounds in Greek plants of Lamiaceae family by HPLC. J. Appl. Res. Med. Arom. Plants 2017, 6, 62–69. [Google Scholar]
- Irakli, M.; Chatzopoulou, P.; Kadoglidou, K.; Tsivelika, N. Optimization and development of a high-performance liquid chromatography method for the simultaneous determination of vitamin E and carotenoids in tomato fruits. J. Sep. Sci. 2016, 39, 3348–3356. [Google Scholar] [CrossRef]
- Aprodu, I.; Banu, I. Milling, functional and thermo-mechanical properties of wheat, rye, triticale, barley and oat. J. Cereal. Sci. 2017, 77, 42–48. [Google Scholar] [CrossRef]
- Felizardo, M.P.; Freire, J.T. Characterization of barley grains in different levels of pearling process. J. Food Eng. 2018, 232, 29–35. [Google Scholar] [CrossRef]
- Edney, M.J.; Rossnagel, B.G.; Endo, Y.; Ozawa, S.; Brophyk, M. Pearling quality of Canadian barley cultivars and their potential use as rice extenders. J. Cereal Sci. 2002, 36, 295–305. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Dexter, J.E.; Desjardins, R.G. Roller milling of Canadian hull-less barley: Optimization of roller milling conditions and composition of mill streams. Cereal Chem. 2003, 80, 637–644. [Google Scholar] [CrossRef]
- Papageorgiou, M.; Lakhdara, N.; Lazaridou, A.; Biliaderis, C.G.; Izydorczyk, M.S. Water extractable (1→3, 1→4)-β-D-glucans from barley and oats: An intervarietal study on their structural features and rheological behaviour. J. Cereal Sci. 2005, 42, 213–224. [Google Scholar] [CrossRef]
- Zheng, X.; Li, L.; Wang, Q. Distribution and molecular characterization of β-glucans from hull-less barley bran, shorts and flour. Int. J. Mol. Sci. 2011, 12, 1563–1574. [Google Scholar] [CrossRef] [Green Version]
- Cavallero, A.; Gianinetti, A.; Finocchiaro, F.; Delogu, G.; Stanca, M. Tocols in hull-less and hulled barley genotypes grown in contrasting environments. J. Cereal Sci. 2004, 39, 175–180. [Google Scholar] [CrossRef]
- Šimić, G.; Horvat, D.; Lalic, A.; Komlenić, D.K.; Abicic, I.; Zdunić, Z. Distribution of β-glucan, phenolic acids, and proteins as functional phytonutrients of hull-less barley grain. Foods 2019, 8, 680. [Google Scholar] [CrossRef] [Green Version]
- Moza, J.; Gujral, H.S. Influence of non-starchy polysaccharides on barley milling behavior and evaluating bioactive composition of milled fractions. Food Chem. 2017, 218, 137–143. [Google Scholar] [CrossRef]
- Dvorakova, M.; Moreira, M.M.; Dostalek, P.; Skulilova, Z.; Guido, L.F.; Barros, A.A. Characterization of monomeric and oligomeric flavan-3-ols from barley and malt by liquid chromatography–ultraviolet detection–electrospray ionization mass spectrometry. J. Chrom. A 2008, 1189, 398–405. [Google Scholar] [CrossRef]
- Kim, M.-J.; Hyun, J.-N.; Kim, J.-A.; Park, J.-C.; Kim, M.-Y.; Kim, J.-G.; Lee, S.-J.; Chun, S.-C.; Chung, I.-M. Relationship between phenolic compounds, anthocyanins content and antioxidant activity in colored barley germplasm. J. Agric. Food Chem. 2007, 55, 4802–4809. [Google Scholar] [CrossRef]
- Holtekjolen, A.K.; Kinitz, C.; Knutsen, S.H. Flavanol and bound phenolic acid contents in different barley cultivars. J. Agric. Food Chem. 2006, 54, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Sonoda, K.; Nogata, Y.; Nagamine, T.; Sato, M.; Oki, T.; Hashimoto, S.; Ohta, H. Determination of free and bound phenolic acids, and evaluation of antioxidant activities and total polyphenolic contents in selected pearled barley. Food Sci. Technol. Res. 2010, 16, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-J.; Dang, B.; Fan, M.-T. Free and bound phenolic compound content and antioxidant activity of different cultivated blue highland barley cultivars from the Qinghai-Tibet plateau. Molecules 2018, 23, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | TGW (g) | Length (mm) | Width (mm) | Thickness (mm) | Pearling Time (min) | PG Whiteness | PG L* |
|---|---|---|---|---|---|---|---|
| ANDA | 43.5 c | 8.57 cd | 3.58 cd | 2.93 ab | 6.5 fg | 38.8 e | 91.2 ab |
| NIKI | 46.7 a | 8.25 fg | 3.82 a | 2.99 a | 6.9 ef | 44.0 cd | 91.0 abc |
| PENI | 37.0 f | 8.20 g | 3.36 fg | 2.92 ab | 7.5 ef | 21.4 j | 90.5 de |
| DIMA | 43.2 c | 8.05 h | 3.78 a | 2.92 ab | 7.7 e | 45.3 bc | 91.4 a |
| CONS | 37.0 f | 7.90 i | 3.45 e | 2.74 cd | 13.2 b | 38.8 e | 90.1 ef |
| MAKA | 43.1 c | 8.50 cde | 3.60 c | 2.91 ab | 9.0 c | 36.6 fg | 89.5 g |
| THKI | 40.2 e | 8.60 c | 3.42 ef | 2.86 bc | 8.0 de | 26.8 i | 89.4 g |
| SIOS | 45.5 ab | 8.58 cd | 3.66 b | 2.92 ab | 9.0 cd | 46.3 ab | 90.7 cd |
| KOS | 35.6 g | 8.19 gh | 3.34 g | 2.75 cd | 9.2 c | 31.1 h | 88.4 h |
| TIOS | 41.8 d | 10.13 a | 3.25 h | 2.99 a | 8.9 cd | 35.6 g | 90.3 def |
| THMI | 43.1 cd | 8.22 fg | 3.52 d | 2.91 ab | 9.2 c | 35.9 g | 90.5 de |
| BIZA | 45.2 b | 9.08 b | 3.59 c | 2.92 ab | 10.0 c | 47.8 a | 90.9 bc |
| CYPS | 36.6 fg | 8.45 de | 3.26 h | 2.77 c | 13.7 b | 43.0 d | 90.7 cd |
| IPOS | 27.8 h | 7.72 j | 3.06 i | 2.64 d | 15.5 a | 38.4 ef | 89.4 g |
| HLES | 45.2 b | 8.36 ef | 3.39 fg | 2.78 c | 5.7 g | 48.0 a | 89.2 g |
| Mean | 40.6 | 8.45 | 3.47 | 2.86 | 9.3 | 38.5 | 90.3 |
| CV% | 12.0 | 6.6 | 5.8 | 3.6 | 29.1 | 19.7 | 0.9 |
| Pearling Time | Pearled Whiteness | Pearled Brightness | Weight | Length | Thick-ness | Width | |
|---|---|---|---|---|---|---|---|
| Pearling time | 1.00 | ||||||
| Pearled whiteness | 0.05 | 1.00 | |||||
| Pearled brightness | −0.17 | 0.30 * | 1.00 | ||||
| Weight | −0.71 *** | 0.41 ** | 0.52 *** | 1.00 | |||
| Length | −0.25 | 0.04 | 0.18 | 0.42 ** | 1.00 | ||
| Thickness | −0.64 *** | 0.02 | 0.59 *** | 0.81 *** | 0.55 *** | 1.00 | |
| Width | −0.56 *** | 0.38 ** | 0.55 *** | 0.84 *** | −0.05 | 0.65 *** | 1.00 |
| Cultivar | Dehulled Flour (DF) | Pearled Flour (PF) | Bran | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Protein | Ash | β-glucan | Protein | Ash | β-glucan | Protein | Ash | β-glucan | |
| ANDA | 13.48 d | 1.61 bcd | 3.94 ef | 11.69 ef | 1.10 abc | 4.23 fg | 23.65 e | 4.87 bcd | 4.09 bc |
| NIKI | 11.39 f | 1.22 g | 3.87 f | 9.98 h | 0.72 f | 4.29 f | 21.39 f | 3.94 f | 3.58 fg |
| PENI | 12.31 e | 1.33 efg | 3.88 ef | 10.78 g | 0.96 de | 4.39 ef | 21.71 f | 4.07 f | 3.49 g |
| DIMA | 9.60 g | 1.27 fg | 4.29 cd | 8.51 i | 0.54 g | 4.54 de | 18.32 g | 4.69 de | 3.85 de |
| CONS | 16.06 a | 1.59 bcd | 4.67 b | 14.58 a | 0.91 e | 4.85 bc | 27.21 a | 4.73 de | 4.26 b |
| MAKA | 15.69 ab | 1.48 cf | 3.96 ef | 13.91 ab | 0.89 e | 4.64 cd | 26.87 ab | 4.51 e | 3.68 efg |
| THKI | 13.71 d | 1.61 bcd | 4.16 de | 12.14 de | 0.87 e | 3.99 gh | 26.00 c | 4.60 de | 3.75 ef |
| SIOS | 13.21 d | 1.73 ab | 5.08 a | 12.15 de | 0.69 f | 5.18 a | 23.94 de | 4.73 de | 4.78 a |
| KOS | 15.21 bc | 1.87 a | 4.57 bc | 13.47 bc | 1.09 abc | 4.83 bc | 25.76 c. | 4.87 bcd | 4.00 cd |
| TIOS | 11.90 ef | 1.64 bc | 4.27 d | 10.61 gh | 1.00 cde | 4.48 def | 21.84 f | 5.13 bc | 2.99 h |
| THMI | 13.51 d | 1.55 be | 3.41 g | 11.76 ef | 1.08 bcd | 3.75 h | 24.51 d | 4.79 cde | 3.00 h |
| BIZA | 12.17 e | 1.61 bcd | 3.91 ef | 11.08 fg | 0.88 e | 4.25 f | 23.82 de | 5.17 b | 2.98 h |
| CYPS | 15.18 bc | 1.89 a | 4.76 b | 12.63 d | 1.08 abc | 4.90 b | 26.03 bc | 5.20 b | 4.65 a |
| IPOS | 14.99c | 1.70 abc | 4.07 def | 12.80 cd | 1.21 a | 4.30 ef | 26.03 bc | 4.73 de | 3.60 fg |
| HLES | 15.34 bc | 1.40 dg | 5.25 a | 14.39 a | 1.16 a | 5.27 a | 25.42 c | 5.58 a | 3.00 h |
| Mean ± SD | 13.58 ± 1.81 | 1.57 ± 0.20 | 4.27 ± 0.50 | 12.03 ± 1.66 | 0.95 ± 0.19 | 4.53 ± 0.42 | 24.17 ± 2.42 | 4.77 ± 0.42 | 3.71 ± 0.57 |
| Fraction | Cultivar | Phenolic Acids | Flavanols | Tocotrienols | Tocopherols | Tocols |
|---|---|---|---|---|---|---|
| mg/100 g | μg/g | |||||
| Dehulled flour (DF) | ANDA | 47.8 ± 0.8 de | 51.6 ± 2.7 e | 41.2 ± 0.6 de | 10.4 ± 0.6 cd | 51.6 ± 0.8 def |
| NIKI | 40.1 ± 0.7 ef | 39.6 ± 1.4 f | 44.9 ± 1.0 cd | 8.3 ± 0.1 e | 53.2 ± 4.8 de | |
| PENI | 55.1 ± 0.4 a-d | 61.9 ± 3.0 abc | 53.9 ± 2.1 ab | 11.3 ± 0.1 c | 65.2 ± 2.0 ab | |
| DIMA | 48.8 ± 7.4 d | 54.0 ± 2.1 de | 45.2 ± 0.4 cd | 7.6 ± 0.4 e | 52.8 ± 3.2 de | |
| CONS | 51.2 ± 1.0 cd | 42.1 ± 1.6 f | 25.0 ± 0.5 f | 13.3 ± 0.5 b | 38.3 ± 1.3 gh | |
| MAKA | 60.4 ± 6.8 ab | 60.5 ± 0.8 bcd | 29.0 ± 0.2 f | 8.9 ± 0.3 e | 37.9 ± 1.9 gh | |
| THKI | 62.6 ± 3.0 a | 63.4 ± 5.9 ab | 47.9 ± 0.2 bc | 8.9 ± 0.3 e | 56.8 ± 0.2 cd | |
| SIOS | 52.6 ± 0.7 bcd | 54.5 ± 0.3 de | 40.6 ± 1.4 de | 8.4 ± 0.2 e | 49.0 ± 1.5 ef | |
| KOS | 58.8 ± 7.3 abc | 55.5 ± 7.9 cde | 36.7 ± 0.1 e | 8.2 ± 0.1 e | 44.9 ± 0.9 fg | |
| TIOS | 53.2 ± 2.0 bcd | 54.1 ± 0.2 de | 53.8 ± 0.6 ab | 8.5 ± 0.5 e | 62.3 ± 0.1 bc | |
| THMI | 34.1 ± 0.6 f | 58.7 ± 1.5 bcd | 54.5 ± 0.1 a | 14.2 ± 0.9 b | 68.7 ± 0.4 ab | |
| BIZA | 59.2 ± 1.0 abc | 61.5 ± 2.1 abc | 46.5 ± 1.7 cd | 7.9 ± 0.5 e | 54.5 ± 1.9 de | |
| CYPS | 60.1 ± 2.7 ab | 63.9 ± 0.1 ab | 40.6 ± 1.5 d | 11.0 ± 1.0 c | 51.6 ± 2.0 def | |
| IPOS | 58.6 ± 4.5 abc | 67.2 ± 1.1 a | 55.1 ± 3.6 a | 16.6 ± 0.6 a | 71.7 ± 3.9 a | |
| HLES | 58.2 ± 2.9 abc | 39.2 ± 4.5 f | 27.7 ± 2.1 f | 8.7 ± 0.3 e | 36.3 ± 2.3 h | |
| Mean | 53.4 ± 8.2 | 55.2 ± 8.8 | 42.8 ± 9.8 | 10.1 ± 2.6 | 53.0 ± 10.8 | |
| Pearled flour (PF) | ANDA | 20.0 ± 0.3 e-h | 52.9 ± 1.3 cd | 26.7 ± 0.2 cd | 10.5 ± 0.3 a | 37.3 ± 0.1 b |
| NIKI | 26.4 ± 2.5 bcd | 37.2 ± 0.4 f | 19.8 ± 0.2 fg | 4.5 ± 0.1 hij | 24.3 ± 0.3 ef | |
| PENI | 29.2 ± 0.8 bc | 74.5 ± 1.5 a | 35.6 ± 0.4 b | 7.5 ± 0.2 c | 43.2 ± 0.5 a | |
| DIMA | 36.2 ± 3.2 a | 56.5 ± 6.1 bc | 15.9 ± 1.2 h | 3.8 ± 0.2 jk | 19.7 ± 1.2 g | |
| CONS | 19.5 ± 0.8 fgh | 46.5 ± 2.1 e | 9.1 ± 0.5 i | 4.7 ± 0.3 ghi | 13.8 ± 0.2 h | |
| MAKA | 20.7 ± 0.7 e-h | 56.0 ± 0.4 bc | 15.8 ± 0.6 h | 4.7 ± 0.3 i | 20.5 ± 0.8 g | |
| THKI | 38.7 ± 4.1 a | 56.2 ± 1.8 bc | 24.1 ± 1.1 de | 4.7 ± 0.4 i | 28.1 ± 0.9 cd | |
| SIOS | 17.3 ± 0.4 h | 53.2 ± 0.1 cd | 18.3 ± 0.2 gh | 4.8 ± 0.1 fgh | 23.1 ± 0.3 efg | |
| KOS | 23.1 ± 1.5 d-g | 56.9 ± 3.9 bc | 23.2 ± 0.8 e | 5.4 ± 0.1 efg | 28.6 ± 0.8 cd | |
| TIOS | 24.7 ± 0.1 cde | 54.3 ± 5.8 c | 35.6 ± 2.4 b | 3.8 ± 0.2 jk | 39.4 ± 2.5 b | |
| THMI | 18.8 ± 0.4 gh | 45.1 ± 2.7 e | 40.5 ± 1.1 a | 5.6 ± 0.0 e | 46.1 ± 1.0 a | |
| BIZA | 24.25 ± 1.1 c-f | 62.0 ± 1.4 b | 22.7 ± 1.2 ef | 3.6 ± 0.2 k | 26.3 ± 1.4 de | |
| CYPS | 24.0 ± 4.0 def | 44.2 ± 5.3 e | 24.7 ± 1.1 de | 6.6 ± 0.2 d | 31.4 ± 1.2 c | |
| IPOS | 31.2 ± 5.2 b | 47.2 ± 0.4 de | 28.2 ± 2.1 c | 9.3 ± 0.3 b | 37.5 ± 2.2 b | |
| HLES | 20.5 ± 0.1 e-h | 54.0 ± 0.7 c | 15.7 ± 0.9 h | 5.5 ± 0.2 ef | 21.2 ± 0.9 fg | |
| Mean | 25.0 ± 6.4 | 53.1 ± 8.7 | 26.2 ± 8.6 | 5.6 ± 2.0 | 29.4 ± 9.3 | |
| Bran | ANDA | 132.7 ± 4.1 e | 164.5 ± 6,5 bc | 152.1 ± 6.1 cd | 26.0 ± 2.8 bcd | 178.1 ± 3.3 cd |
| NIKI | 136.3 ± 1.4 de | 124.6 ± 0,6 ef | 176.9 ± 6.9 b | 25.6 ± 1.0 b-e | 202.5 ± 7.9 b | |
| PENI | 159.4 ± 9.0 ab | 197.2 ± 5,0 a | 159.0 ± 5.0 bc | 25.5 ± 0.5 b-e | 184.5 ± 4.5 bc | |
| DIMA | 135.5 ± 2.1 de | 149.5 ± 0,7 cd | 210.8 ± 10.8 a | 21.4 ± 0.6 fg | 232.2 ± 10.1 a | |
| CONS | 150.3 ± 2.4 bc | 93.2 ± 2.8 g | 124.7 ± 4.7 ef | 22.9 ± 0.9 def | 147.6 ± 5.6 ef | |
| MAKA | 102.5 ± 4.0 g | 114.9 ± 5.7 ef | 131.2 ± 4.3 ef | 23.5 ± 0.5 c-f | 155.2 ± 4.8 e | |
| THKI | 139.4 ± 1.8 cde | 120.1 ± 7.0 ef | 204.4 ± 7.6 a | 22.3 ± 0.9 ef | 226.8 ± 8.4 a | |
| SIOS | 118.0 ± 1.8 f | 133.3 ± 1.0 de | 163.7 ± 13.7 bc | 29.9 ± 0.1 a | 193.5 ± 13.5 bc | |
| KOS | 112.0 ± 7.6 fg | 132.0 ± 3.0 de | 131.3 ± 4.7 ef | 26.4 ± 0.6 bc | 157.7 ± 5.3 de | |
| TIOS | 163.6 ± 9.6 a | 155.1 ± 8.3 c | 217.6 ± 7.6 a | 29.8 ± 0.6 a | 247.4 ± 7.0 a | |
| THMI | 109.5 ± 5.1 fg | 110.5 ± 3.5 fg | 206.1 ± 5.2 a | 30.9 ± 1.1 a | 237.0 ± 6.3 a | |
| BIZA | 140.8 ± 3.8 cde | 175.6 ± 9.0 b | 160.2 ± 2.9 bc | 22.9 ± 1.3 def | 183.1 ± 4.2 bc | |
| CYPS | 150.6 ± 0.5 bc | 208.0 ± 12.7 a | 132.3 ± 3.3 de | 27.7 ± 1.3 ab | 160.0 ± 4.6 de | |
| IPOS | 165.8 ± 5.5 a | 214.0 ± 24.0 a | 112.0 ± 2.9 f | 18.4 ± 0.5 g | 130.4 ± 3.4 f | |
| HLES | 146.3 ± 0.1 cd | 149.1 ± 11.1 cd | 124.9 ± 5.1 ef | 28.7 ± 1.3 ab | 153.7 ± 6.3 e | |
| Mean | 137.5 ± 19.6 | 149.4 ± 36.1 | 160.5± 35.0 | 25.5 ± 3.6 | 186.0 ± 36.1 | |
| TPC | TFC | TPAC | PAs | Flavanols | Tocols | ABTS | FRAP | |
|---|---|---|---|---|---|---|---|---|
| TPC | 1.00 | |||||||
| TFC | 0.97 *** | 1.00 | ||||||
| TPAC | 0.91 *** | 0.85 *** | 1.00 | |||||
| PAs | 0.95 *** | 0.90 *** | 0.92 *** | 1.00 | ||||
| Flavanols | 0.88 *** | 0.82 *** | 0.93 *** | 0.91 *** | 1.00 | |||
| Tocols | 0.92 *** | 0.88 *** | 0.86 *** | 0.91 *** | 0.83 *** | 1.00 | ||
| ABTS | 0.98 *** | 0.97 *** | 0.90 *** | 0.95 *** | 0.89 *** | 0.93 *** | 1.00 | |
| FRAP | 0.98 *** | 0.97 *** | 0.88 *** | 0.95 *** | 0.86 *** | 0.93 *** | 0.99 *** | 1.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irakli, M.; Lazaridou, A.; Mylonas, I.; Biliaderis, C.G. Bioactive Components and Antioxidant Activity Distribution in Pearling Fractions of Different Greek Barley Cultivars. Foods 2020, 9, 783. https://doi.org/10.3390/foods9060783
Irakli M, Lazaridou A, Mylonas I, Biliaderis CG. Bioactive Components and Antioxidant Activity Distribution in Pearling Fractions of Different Greek Barley Cultivars. Foods. 2020; 9(6):783. https://doi.org/10.3390/foods9060783
Chicago/Turabian StyleIrakli, Maria, Athina Lazaridou, Ioannis Mylonas, and Costas G. Biliaderis. 2020. "Bioactive Components and Antioxidant Activity Distribution in Pearling Fractions of Different Greek Barley Cultivars" Foods 9, no. 6: 783. https://doi.org/10.3390/foods9060783
APA StyleIrakli, M., Lazaridou, A., Mylonas, I., & Biliaderis, C. G. (2020). Bioactive Components and Antioxidant Activity Distribution in Pearling Fractions of Different Greek Barley Cultivars. Foods, 9(6), 783. https://doi.org/10.3390/foods9060783