ACE Inhibitory Properties and Phenolics Profile of Fermented Flours and of Baked and Digested Biscuits from Buckwheat

 and
and
Abstract
:
1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Fermentation of Buckwheat Flour and Preparation of Buckwheat Biscuits
2.3. In Vitro Digestion of Buckwheat Water Biscuits
2.4. Determination of Total Phenolic Content (TPC) of Buckwheat Fermented Flours and Buckwheat Water Biscuits before and after In Vitro Digestion
2.5. Extraction and Isolation of the Main Phenolic Compounds from Buckwheat Fermented Flour and Buckwheat Water Biscuits before and after In Vitro Digestion
2.6. Sample Preparation for ACE Inhibitory Activity Determination
2.7. Angiotensin-I Converting Enzyme Inhibitory Assay
2.8. Statistical Analysis
3. Results and Discussion
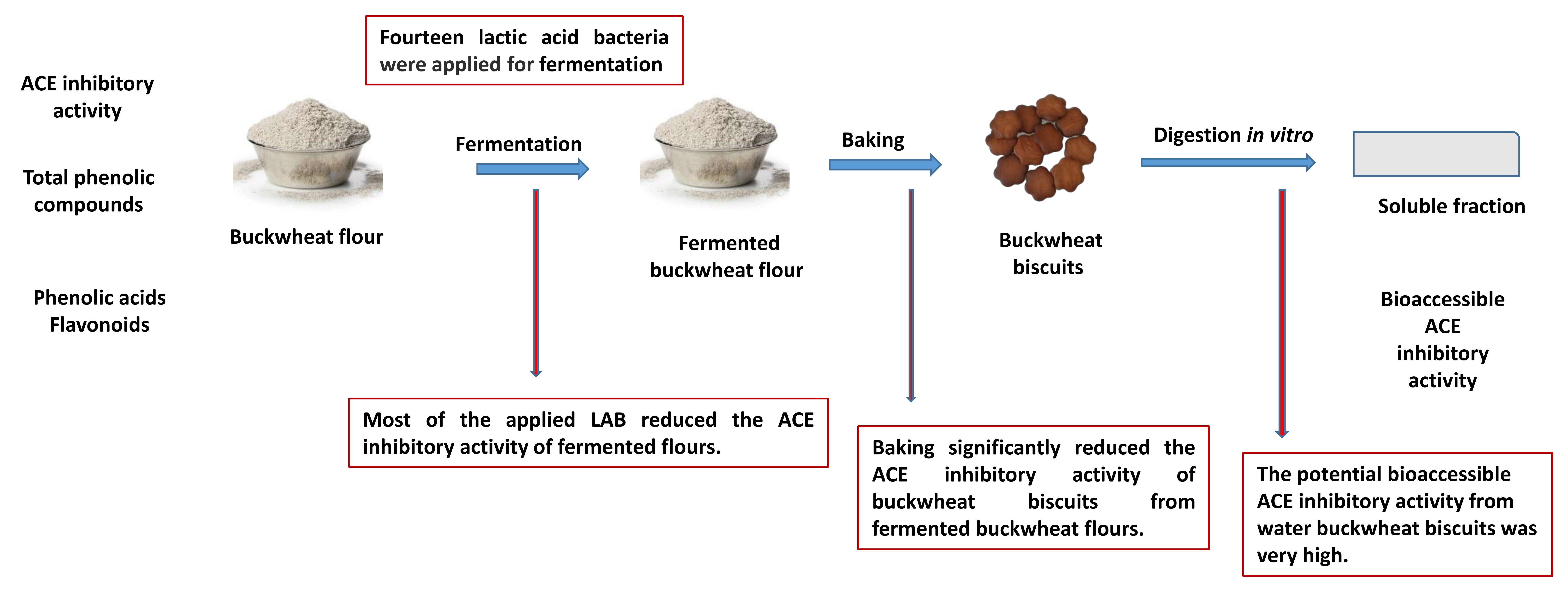
3.1. Effect of Fermentation on ACE Inhibitory Activity of Buckwheat Flour
3.2. Effect of Baking on ACE Inhibitory Activity of Biscuits Formulated from Fermented Buckwheat Flours
3.3. Effect of In Vitro Digestion on ACE Inhibitory Activity of Biscuits Formulated from Fermented Buckwheat Flours
3.4. The Bioaccessible ACE Inhibitory Activity of Biscuits Formulated from Fermented Buckwheat Flours
3.5. Effect of Fermentation, Baking, and In Vitro Digestion on the Content of Total Phenolic Compounds in Buckwheat Flours and Buckwheat Water Biscuits before and after Digestion In Vitro
3.6. Profile and Content of the Main Phenolic Compounds from Buckwheat Fermented Flour and Buckwheat Water Biscuits before and after In Vitro Digestion
3.7. Correlation between TPC and ACE Inhibitory Activity of Fermented Buckwheat Flours and Biscuits Formulated from Fermented Buckwheat Flours before and after In Vitro Digestion.
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ondetti, M.A.; Rubin, B.; Cushman, D.W. Enzyme of the rennin angiotensin system and their inhibitors. Annu. Rev. Biochem. 1982, 51, 283–308. [Google Scholar] [CrossRef]
- Iroyukifujita, H.; Eiichiyokoyama, K.; Yoshikawa, M. Classification and Antihypertensive Activity of Angiotensin I-Converting Enzyme Inhibitory Peptides Derived from Food Proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar] [CrossRef]
- Guang, C.; Phillips, R.D. Plant Food-Derived Angiotensin I Converting Enzyme Inhibitory Peptides. J. Agric. Food Chem. 2009, 57, 5113–5120. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Byun, H.G.; Park, P.J.; Shahidi, F. Angiotensin I converting enzyme inhibitory peptides purified from bovine skin gelatin hydrolysate. J. Agric. Food Chem. 2001, 49, 2992–2997. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kumar, A.; Sharma, R.; Baruwa, A. Pharmacological review on natural ACE inhibitors. Der. Pharm. Lett. 2010, 2, 273–293. [Google Scholar]
- Iwaniak, A.; Mińkiewicz, P.; Darewicz, M. Food-Originating ACE Inhibitors, Including Antihypertensive Peptides, as Preventive Food Components in Blood Pressure Reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Miguel, M.; Contreras, M.D.M.; Recio, I.; Aleixandre, A. ACE-inhibitory and antihypertensive properties of a bovine casein hydrolysate. Food Chem. 2009, 112, 211–214. [Google Scholar] [CrossRef]
- Je, J.Y.; Qian, Z.J.; Byun, H.G.; Kim, S.K. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Kuba, M.; Tana, C.; Tawata, S.; Yasuda, M. Production of angiotensin I-converting enzyme inhibitory peptides from soybean protein with Monascus purpureus acid proteinase. Process Biochem. 2005, 40, 2191–2196. [Google Scholar] [CrossRef]
- Matsui, T.; Li, C.H.; Osajima, Y. Preparation and characterization of novel bioactive peptides responsible for angiotensin I-converting enzyme inhibition from wheat germ. J. Pept. Sci. 1999, 5, 289–297. [Google Scholar] [CrossRef]
- Torino, M.I.; Limón, R.I.; Martinez-Villaluenga, C.; Mäkinen, S.; Pihlanto, A.; Vidal-Valverde, C.; Frias, J. Antioxidant and antihypertensive properties of liquid and solid state fermented lentils. Food Chem. 2013, 136, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Tian, J.; Wang, Q. ACE inhibitory and antihypertensive properties of apricot almond meal hydrolysate. Eur. Food Res. Technol. 2011, 232, 549–556. [Google Scholar] [CrossRef]
- Ren, X.; Ma, H.; Mao, S.; Zhou, H. Effects of sweeping frequency ultrasound treatment on enzymatic preparations of ACE-inhibitory peptides from zein. Eur. Food Res. Technol. 2013, 238, 435–442. [Google Scholar] [CrossRef]
- Galleano, M.; Pechanova, O.; Fraga, C.G. Hypertension, nitric oxide, oxidants, and dietary plant polyphenols. Curr. Pharm. Biotechnol. 2010, 11, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Lacaille-Dubois, M.A.; Franck, U.; Wagner, H. Search for potential angiotensin converting enzyme (ACE)-inhibitors from plants. Phytomedicine 2001, 8, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G. Sourdough lactic acid bacteria: Exploration of non-wheat cereal-based fermentation. Food Microbiol. 2014, 37, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bastida, J.A.; Zieliński, H. Buckwheat as a Functional Food and Its Effects on Health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef]
- Higasa, S.; Fujihara, S.; Hayashi, A.; Kimoto, K.; Aoyagi, Y. Distribution of a novel angiotensin I-converting enzyme inhibitory substance (2″-hydroxynicotianamine) in the flour, plant parts, and processed products of buckwheat. Food Chem. 2011, 125, 607–613. [Google Scholar] [CrossRef]
- Tsai, H.; Deng, H.; Tsai, S.; Hsu, Y. Bioactivity comparison of extracts from various parts of common and tartary buckwheats: Evaluation of the antioxidant- and angiotensin-converting enzyme inhibitory activities. Chem. Cent. J. 2012, 6, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.H.; Matsui, T.; Matsumoto, K.; Yamasaki, R.; Kawasaki, T. Latent production of angiotensin I-converting enzyme inhibitors from buckwheat protein. J. Pept. Sci. 2002, 8, 267–274. [Google Scholar] [CrossRef]
- Aoyagi, Y. An angiotensin-I converting enzyme inhibitor from buckwheat (Fagopyrum esculentum Moench) flour. Phytochemistry 2006, 67, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.-S.; Bae, I.Y.; Lee, H.G.; Yang, C.-B. Purification and identification of angiotensin I-converting enzyme inhibitory peptide from buckwheat (Fagopyrum esculentum Moench). Food Chem. 2006, 96, 36–42. [Google Scholar] [CrossRef]
- Dziadek, K.; Kopeć, A.; Pastucha, E.; Piątkowska, E.; Leszczyńska, T.; Pisulewska, E.; Witkowicz, R.; Francik, R. Basic chemical composition and bioactive compounds content in selected cultivars of buckwheat whole seeds, dehulled seeds and hulls. J. Cereal Sci. 2016, 69, 1–8. [Google Scholar] [CrossRef]
- Müller, M.R.; Wolfrum, G.; Stolz, P.; Ehrmann, M.; Vogel, R.F. Monitoring the growth of Lactobacillus species during a rye flour fermentation. Food Microbiol. 2001, 18, 217–227. [Google Scholar] [CrossRef]
- Zieliński, H.; Ciesarová, Z.; Kukurová, K.; Zielińska, D.; Szawara-Nowak, D.; Starowicz, M.; Wronkowska, M. Effect of fermented and unfermented buckwheat flour on functional properties of gluten-free muffins. J. Food Sci. Technol. 2017, 54, 1425–1432. [Google Scholar] [CrossRef]
- Guerra, A.; Etienne-Mesmin, L.; Livrelli, V.; Denis, S.; Blanquet-Diot, S.; Alric, M. Relevance and challenges in modeling human gastric and small intestinal digestion. Trends Biotechnol. 2012, 30, 591–600. [Google Scholar] [CrossRef]
- Spínola, V.; Llorent-Martínez, E.J.; Castilho, P.C. Antioxidant polyphenols of Madeira sorrel (Rumex maderensis): How do they survive to in vitro simulated gastrointestinal digestion? Food Chem. 2018, 259, 105–112. [Google Scholar] [CrossRef]
- Wronkowska, M.; Jeliński, T.; Majkowska, A.; Zieliński, H. Physical Properties of Buckwheat Water Biscuits Formulated from Fermented Flours by Selected Lactic Acid Bacteria. Pol. J. Food Nutr. Sci. 2018, 68, 25–31. [Google Scholar] [CrossRef]
- Zieliński, H.; Szawara-Nowak, D.; Bączek, N.; Wronkowska, M. Effect of liquid-state fermentation on the antioxidant and functional properties of raw and roasted buckwheat flours. Food Chem. 2019, 271, 291–297. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A. Heat damage of water biscuits from einkorn, durum and bread wheat flours. Food Chem. 2011, 128, 471–478. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Conde-Aguilera, J.A.; Haro, A.; De La Cueva, S.P.; Rufián-Henares, J.Á. A combined procedure to evaluate the global antioxidant response of bread. J. Cereal Sci. 2010, 52, 239–246. [Google Scholar] [CrossRef]
- Wiczkowski, W.; Szawara-Nowak, D.; Sawicki, T.; Mitrus, J.; Kasprzykowski, Z.; Horbowicz, M.; Information, R. Profile of Phenolic Acids and Antioxidant Capacity in Organs of Common Buckwheat Sprout. Acta Aliment. 2016, 45, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Sentandreu, M.Á.; Toldrá, F. A rapid, simple and sensitive fluorescence method for the assay of angiotensin-I converting enzyme. Food Chem. 2006, 97, 546–554. [Google Scholar] [CrossRef]
- Rui, X.; Wen, D.; Li, W.; Chen, X.; Jiang, M.; Dong, M.S. Enrichment of ACE inhibitory peptides in navy bean (Phaseolus vulgaris) using lactic acid bacteria. Food Funct. 2015, 6, 622–629. [Google Scholar] [CrossRef]
- Xiao, Y.; Xing, G.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M.S. Effect of solid-state fermentation with Cordyceps militaris SN-18 on physicochemical and functional properties of chickpea (Cicer arietinum L.) flour. LWT Food Sci. Technol. 2015, 63, 1317–1324. [Google Scholar] [CrossRef]
- Pyo, Y.H.; Lee, T.C. The Potential Antioxidant Capacity and Angiotensin I-Converting Enzyme Inhibitory Activity of Monascus-Fermented Soybean Extracts: Evaluation of Monascus-Fermented Soybean Extracts as Multifunctional Food Additives. J. Food Sci. 2007, 72, S218–S223. [Google Scholar] [CrossRef]
- Juan, M.Y.; Wu, C.H.; Chou, C.C. Fermentation with Bacillus spp. as a bioprocess to enhance anthocyanin content, the angiotensin converting enzyme inhibitory effect, and the reducing activity of black soybeans. Food Microbiol. 2010, 27, 918–923. [Google Scholar] [CrossRef]
- Mojica, L.; Chen, K.; De Mejia, E.G. Impact of Commercial Precooking of Common Bean (Phaseolus vulgaris) on the Generation of Peptides, After Pepsin-Pancreatin Hydrolysis, Capable to Inhibit Dipeptidyl Peptidase-IV. J. Food Sci. 2014, 80, 188–198. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; Pinto, M.D.S. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef]
- Kawakami, A.; Inbe, T.; Kayahara, H.; Horii, A. Preparations of enzymatic hydrolysates of buckwheat globulin and their angiotensin I converting enzyme inhibitory activities. Curr. Adv. Buckwheat Res. 1995, 2, 927–934. [Google Scholar]
- Burton-Freeman, B. Postprandial metabolic events and fruit-derived phenolics: A review of the science. Br. J. Nutr. 2010, 104, S1–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Lee, Y.J.; Yoon, S. Total flavonoids and phenolics in fermented soy products and their effects on antioxidant activities determined by different assays. J. Korean Soc. Food Cult. 2007, 22, 353–358. [Google Scholar]
- Verni, M.; Verardo, V.; Rizzello, C.G. How Fermentation Affects the Antioxidant Properties of Cereals and Legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Aal, E.-S.M.; Rabalski, I. Effect of baking on free and bound phenoli acids in wholegrain bakery products. J. Cereal Sci. 2013, 57, 312–318. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Strain/Sample | Buckwheat Flour | Buckwheat Biscuits | Digested Buckwheat Biscuits |
|---|---|---|---|
| Control (non-fermented) | 4.16 ± 0.21 b | 26.50 ±1.99 a | 1.52 ± 0.16 b |
| L. plantarum IB | 14.49 ± 0.47 *b | 23.42 ± 4.33 a | 1.80 ± 0.10 *c |
| L. plantarum W42 | 17.05 ± 0.12 *b | 44.39 ± 7.58 *a | 2.11 ± 0.29 *c |
| L. delbrucki subsp. bulgaricus 151 | 3.65 ± 0.14 b | 19.46 ± 3.83 *a | 1.55 ± 0.05 c |
| L. casei Lcy | 8.71 ± 0.51 *b | 27.46 ± 4.11 a | 1.55 ± 0.02 c |
| Streptococcus thermophilus MK-10 | 13.41 ± 0.16 *b | 30.60 ± 1.07 *a | 1.51 ± 0.09 c |
| L. acidophilus La5 | 7.50 ± 0.33 *b | 15.70 ± 0.24 *a | 1.62 ± 0.07 c |
| L. acidophilus V | 11.98 ± 0.04 *b | 25.47 ± 4.97a | 1.21 ± 0.21 c |
| L. acidophilus 145 | 4.80 ± 0.33 b | 7.18 ± 1.23 *a | 1.38 ± 0.20 c |
| L. casei 2K | 8.96 ± 0.01 *b | 23.64 ± 2.48 a | 1.38 ± 0.18 c |
| L. delbrucki subsp. bulgaricus K | 11.02 ± 0.25 *b | 21.05 ± 2.78 *a | 2.39 ± 0.11 *c |
| L. rhamnosus GG | 7.00 ± 0.16 *b | 22.97 ± 1.54 a | 2.05 ± 0.10 *c |
| L. rhamnosus 8/4 | 5.40 ± 0.29 *b | 9.14 ± 1.62 *a | 1.62 ± 0.06 c |
| L. rhamnosus K | 8.95 ± 0.27 *b | 19.05 ± 0.68 *a | 0.99 ± 0.09 *c |
| L. salivarius AWH | 9.74 ± 0.16 *b | 30.19 ± 5.24 a | 2.17 ± 0.05 *b |
| Strain/Sample | Buckwheat Flour | Buckwheat Biscuits | Digested Buckwheat Biscuits |
|---|---|---|---|
| Control | 1.60 ± 0.15 b | 1.22 ± 0.04 c | 7.46 ± 0.26 a |
| L. plantarum IB | 2.16 ± 0.21 *b | 1.83 ± 0.03 *c | 8.33 ± 0.25 *a |
| L. plantarum W42 | 2.03 ± 0.23 *b | 1.47 ± 0.02 *c | 7.32 ±0.25 a |
| L. delbrucki subsp. bulgaricus 151 | 1.95 ± 0.32 *b | 1.42 ± 0.06 *c | 7.99 ± 0.34 a |
| L. casei Lcy | 1.92 ± 0.08 *b | 1.95 ± 0.02 *b | 7.41 ± 0.20 a |
| Streptococcus thermophilus MK-10 | 1.88 ± 0.10 *b | 1.42 ± 0.04 *c | 7.57 ± 0.18 a |
| L. acidophilus La5 | 1.81 ± 0.10 b | 1.94 ± 0.04 *b | 7.69 ± 0.08 a |
| L. acidophilus V | 1.70 ± 0.12 c | 1.90 ± 0.04 *b | 8.49 ± 0.09 *a |
| L. acidophilus 145 | 2.04 ± 0.15 *b | 1.70 ± 0.03 *c | 7.24 ± 0.23 a |
| L. casei 2K | 1.82 ± 0.13 b | 1.27 ± 0.02 c | 6.78 ± 0.24 *a |
| L. delbrucki subsp. bulgaricus K | 1.62 ± 0.10 b | 1.09 ± 0.03 *c | 8.15 ± 0.14 *a |
| L. rhamnosus GG | 1.61 ± 0.18 b | 1.25 ± 0.07 c | 7.54 ± 0.15 a |
| L. rhamnosus 8/4 | 1.78 ± 0.19 b | 1.42 ± 0.09 *c | 7.28 ± 0.26 a |
| L. rhamnosus K | 1.60 ± 0.13 b | 1.21 ± 0.01 c | 6.72 ± 0.10 *a |
| L. salivarius AWH | 1.72 ± 0.10 b | 1.34 ± 0.03 *b | 7.03 ± 0.47 a |
| Sample | Chlorogenic | p-Coumaric | Sinapic | Ferulic | t-Cinnamic | Syringic | Vanillic | Isovanillic | Protocatechuic | Caffeic |
|---|---|---|---|---|---|---|---|---|---|---|
| Non-fermented flour | 0.04 ± 0.01 b | 22.1 ± 0.2 b | 6.0 ± 0.3 b | 122.7 ± 4.1 a | 7.1 ± 0.1 b | 78.8 ± 0.9 bc | 55.5 ± 4.2 c | n.d. | 26.6 ± 0.8 c | 21.8 ± 0.1 c |
| Fermented flour | 0.08 ± 0.05 b | 28.8 ± 8.2 a | 13.2 ± 5.3 b | 104.8 ± 20.3 a | 9.2 ± 4.7 b | 82.2 ± 26.7 b | 9.5 ± 3.8 d | n.d. | 35.0 ± 6.8 bc | 66.7 ± 16.9 a |
| Water biscuits | 0.13 ± 0.17 b | 10.8 ± 6.3 b | 7.6 ± 2.5 b | 3.4 ± 1.0 b | 6.8 ± 2.3 b | 50.6 ± 13.0 c | 100.7 ± 14.2 b | 6.1 ± 4.9 b | 62.6 ± 16.3 bc | 5.8 ± 4.0 c |
| Digested water biscuits | 0.96 ± 0.76 a | 28.7 ± 9.9 a | 24.2 ± 7.9 a | 10.0 ± 1.5 b | 22.5 ± 11.0 a | 177.2 ± 45.1 a | 239.5 ± 22.0 a | 69.2 ± 9.5 a | 267.1 ± 46.7 a | 42.0 ± 11.2 b |
| Sample | Luteolin | Kaempferol | Quercetin | Rutin | Vitexin | Apigenin | Orientin | Epicatechin | Total |
|---|---|---|---|---|---|---|---|---|---|
| Non-fermented flour | 0.26 ± 0.02 b | 1.12 ± 0.02 bc | 8.3 ± 0.1 bc | 376.4 ± 4.3 a | 21.2 ± 1.0 a | 0.30 ± 0.04 bc | 17.6 ± 0.3 a | 183.3 ± 0.6 b | 608.5 |
| Fermented flour | 0.41 ± 0.08 a | 1.16 ± 0.39 b | 8.1 ± 2.0 bc | 355.7 ± 54.9 a | 21.9 ± 2.9 a | 0.6 ± 0.4 c | 18.8 ± 3.4 a | 248.0 ± 38.1 a | 654.7 |
| Water biscuits | 0.15 ± 0.04 c | 0.74 ± 0.31 c | 4.9 ± 2.9 c | 127.6 ± 26.3 b | 12.2 ± 3.3 b | 2.4 ± 0.7 ab | 3.7 ± 2.4 c | 55.5 ± 36.1 c | 207.2 |
| Digested water biscuits | 0.20 ± 0.05b c | 2.06 ± 0.41 a | 11.3 ± 5.3 a | 4.2 ± 1.4 c | 14.1 ± 2.3 b | 3.6 ± 2.7 a | 6.0 ± 1.1 bc | 35.9 ± 10.8 c | 77.4 |
| Strain/Samples | Material Based on Fermented Flour, Water Biscuit, and the Sample after Digestion Flour |
|---|---|
| Control | −0.64 |
| L. plantarum IB | −0.93 |
| L. plantarum W42 | −0.82 |
| L. delbrucki subsp. bulgaricus 151 | −0.65 |
| L. casei Lcy | −0.71 |
| Streptococcus thermophilus MK-10 | −0.85 |
| L. acidophilus La5 | −0.80 |
| L. acidophilus V | −0.82 |
| L. acidophilus 145 | −0.93 |
| L. casei 2K | −0.82 |
| L. delbrucki subsp. bulgaricus K | −0.88 |
| L. rhamnosus GG | −0.72 |
| L. rhamnosus 8/4 | −0.89 |
| L. rhamnosus K | −0.86 |
| L. salivarius AWH | −0.75 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieliński, H.; Honke, J.; Topolska, J.; Bączek, N.; Piskuła, M.K.; Wiczkowski, W.; Wronkowska, M. ACE Inhibitory Properties and Phenolics Profile of Fermented Flours and of Baked and Digested Biscuits from Buckwheat. Foods 2020, 9, 847. https://doi.org/10.3390/foods9070847
Zieliński H, Honke J, Topolska J, Bączek N, Piskuła MK, Wiczkowski W, Wronkowska M. ACE Inhibitory Properties and Phenolics Profile of Fermented Flours and of Baked and Digested Biscuits from Buckwheat. Foods. 2020; 9(7):847. https://doi.org/10.3390/foods9070847
Chicago/Turabian StyleZieliński, Henryk, Joanna Honke, Joanna Topolska, Natalia Bączek, Mariusz Konrad Piskuła, Wiesław Wiczkowski, and Małgorzata Wronkowska. 2020. "ACE Inhibitory Properties and Phenolics Profile of Fermented Flours and of Baked and Digested Biscuits from Buckwheat" Foods 9, no. 7: 847. https://doi.org/10.3390/foods9070847
APA StyleZieliński, H., Honke, J., Topolska, J., Bączek, N., Piskuła, M. K., Wiczkowski, W., & Wronkowska, M. (2020). ACE Inhibitory Properties and Phenolics Profile of Fermented Flours and of Baked and Digested Biscuits from Buckwheat. Foods, 9(7), 847. https://doi.org/10.3390/foods9070847