Control of Foodborne Biological Hazards by Ionizing Radiations



Abstract
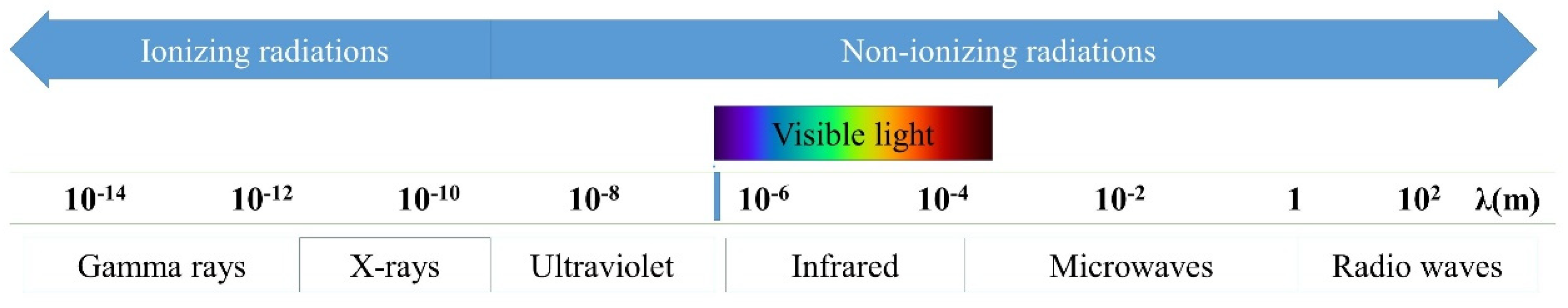
:1. Introduction
2. Methods of Production and Application of β and γ Rays
2.1. Electron Accelerators
2.2. Radioactive Sources
3. Effects on Microorganisms and Pests
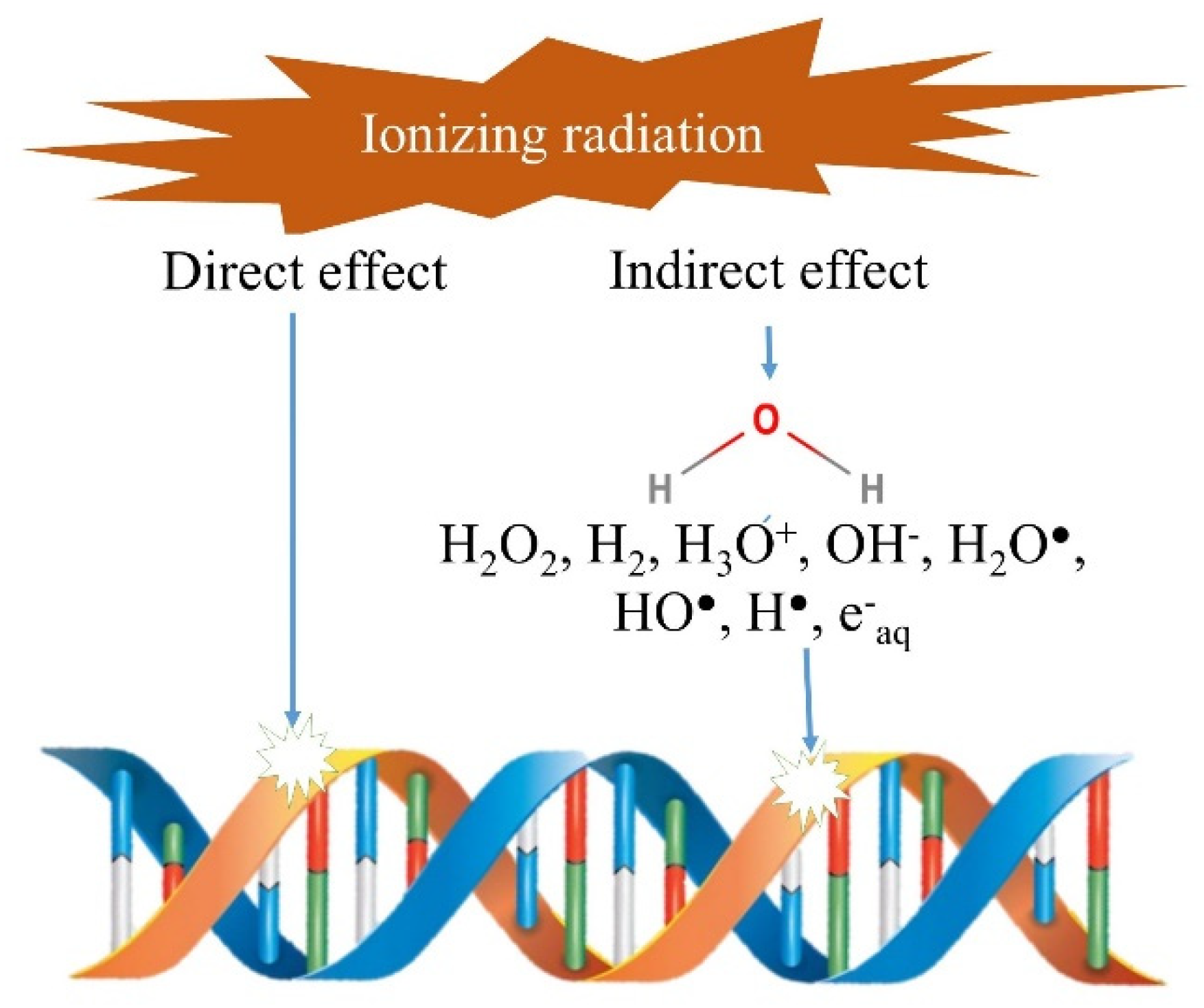
3.1. Mode of Action of β and γ Rays
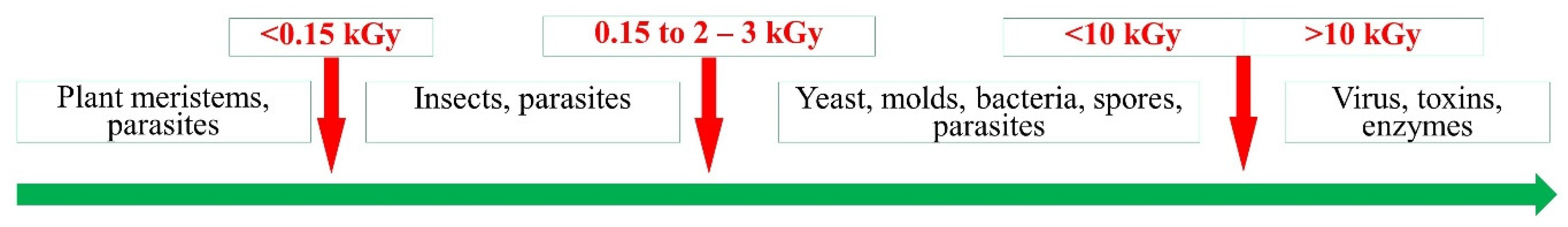
3.2. Doses of Irradiation for the Microbial Inactivation, Pest Sterilization and Inhibition of Germination
3.3. The Principle of Radiobacteriology
- That the number of microorganisms destroyed depends on the initial number (No).
- That for each equivalent dose of ionizing radiation, there is an identical proportion of microbial population destroyed (cumulative effect).
- That according to the exponential law we cannot reach zero.
3.4. Factors Affecting the Kinetics of Destruction of Microorganisms by Ionizing Radiation
3.4.1. Intrinsic Factors
3.4.2. Extrinsic Factors
3.5. Effect on Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | DRD (kGy) | Treatment Media | Temperature |
|---|---|---|---|
| Aeromonas hydrophila | 0.14–0.19 | Minced beef | +2 °C |
| Bacillus cereus (spores) | 3.2 | Nutrient broth | Room |
| Bacillus cereus (spores) | 2 | Water | Room |
| Brucella abortus | 0.34 | Minced beef | Room |
| Campylobacter jejuni | 0.16 | Minced beef | +0–5 °C |
| C. botulinum type A (spores) | 3.3 | Phosphate buffer | Room |
| C. botulinum type B (spores) | 3.3 | Phosphate buffer | Room |
| C. type E (spores) | 1.4 | Irish beef | Room |
| C. perfringens type C (spores) | 2.1 | Water | Room |
| Cronobacter spp. | 1.71 | Milk powder | Room |
| Cronobacter spp. | 0.76 | Milk powder | Room |
| Cronobacter sakazakii | 4.83 | Dehydrated milk | Room |
| E. coli O157:H7 | 0.27 | Chicken meat | +5 °C |
| Listeria monocytogenes | 0.42–0.49 | Chicken meat | +10 °C |
| Listeria monocytogenes | 0.21 | Endive | +2 °C |
| Listeria monocytogenes | 0.5 | Broccoli | −5 °C |
| Listeria monocytogenes | 0.76 | Broccoli | −20 °C |
| Salmonella Enteritidis | 0.33 | Whole egg | +15 °C |
| Salmonella Newport | 0.32 | Whole egg | 0 °C |
| Salmonella Panama | 0.46 | Chicken meat | +22 °C |
| Salmonella Paratyphi B | 0.3 | Crab meat | NA |
| Salmonella Typhimurium | 0.4–0.48 | Chicken meat | +22 °C |
| Salmonella Typhimurium | 0.5–0.55 | Whole egg | 0 °C |
| S. Typhimurium | 0.45 | Ham | +10 °C |
| Shigella dysentariae | 0.22 | Shrimps | −18 °C |
| Shigella flexneri | 0.41 | Shrimps | −18 °C |
| Staphylococcus aureus | 0.58 | Minced beef | Room |
| Staphylococcus aureus | 0.39 | Ham | +10 °C |
| Vibrio parahaemolyticus | 0.03–0.06 | Fish | Room |
| Yersinia enterocolitica | 0.1–0.21 | Minced beef | Room |
| Bacteria | DRD (kGy) | Treatment Media | Temperature |
|---|---|---|---|
| B. pumilus (spores) | 1.35 | Nutrient broth | Room |
| B. pumilus (spores) | 3 | Dry medium | Room |
| B. subtilis (spores) | 0.64 | Saline water | Room |
| B.stearothermophilus (spores) | 1 | Phosphate buffer | Room |
| Lactobacillus spp | 0.3–0.88 | Minced beef | Room |
| Lactobacillus spp | 0.55–0.81 | Nutrient broth | Room |
| Leuconostoc mesenteroïdes | 0.12–0.14 | Water | Room |
| Moraxella sp. | 4.7 | Nutrient broth | Room |
| Pseudomonas fluorescens | 0.12 | Minced beef | Room |
| P. putida | 0.08 | Chicken meat | +10 °C |
| P. aeruginosa | 0.07 | Nutrient broth | NA |
| Proteus vulgaris | 0.2 | Oyster | +5 °C |
3.6. Effect on Yeast and Molds
3.7. Effect on Viruses (and Prions)
3.8. Effect on Parasites
3.9. Effect on Insects
4. Regulatory and Enforcement Status of Application of Ionizing Irradiation
4.1. Regulatory Aspects
4.2. Applications of Authorized Ionizing Treatments
5. Controversies Concerning Consumer Perception and Acceptance of Food Ionization
- The first step is communicating scientific information in public [105]. It can be done at different platforms, especially targeting consumer values. Galati et al. [103] emphasized the importance of specific food education programs and promotional campaigns supported by the ministry of education and other public and private institutions, aimed at creating greater awareness and attitude among consumers about irradiated products.
- The policymakers and food chain managers should also be involved in the promotion campaigns aimed to familiarize the consumers about the principles, aims and benefits of irradiation technology.
- The values of consumers should be addressed rather than the product. For example, highlighting the advantages of technology in keeping food “fresh” and “natural” would be more effective rather than just pointing out the technology.
- The information in promotional campaigns should take into account the positive and negative aspects of technology that will coexist in any food debate.
- Labeling the products to show advantage information as an assurance of food safety and customer values can decrease the consumer opposition to irradiated food.
- Create a partnership with food small or medium-sized retailers, so they can promote the marketing of irradiated food.
- Finally, take the stakeholders on the board who believe in the value of food irradiation, and thus food retailers will be seen as less biased and consumer trust will increase.
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. What Is Ionizing Radiation? Available online: http://www.who.int/ionizing_radiation/about/what_is_ir/en/ (accessed on 19 March 2020).
- Lima, F.; Vieira, K.; Santos, M.; deSouza, P.M. Effects of radiation technologies on food nutritional quality. Descr. Food Sci. 2018. [Google Scholar] [CrossRef] [Green Version]
- FDA. Food Irradiation: What You Need to Know; FDA: Washington, DC, USA, 2020.
- Bouzarjomehri, F.; Dad, V.; Hajimohammadi, B.; Shirmardi, S.P.; Yousefi-Ghaleh Salimi, A. The effect of electron-beam irradiation on microbiological properties and sensory characteristics of sausages. Radiat. Phys. Chem. 2020, 168, 108524. [Google Scholar] [CrossRef]
- Federighi, M. Les rayonnements ionisants comme méthode de préservation des aliments: État de l’art. Rev. Med. Vet. 2019, 4–6, 58–72. [Google Scholar]
- Ehlermann, D.A.E. The early history of food irradiation. Radiat. Phys. Chem. 2016, 129, 10–12. [Google Scholar] [CrossRef]
- Farkas, J.; Mohácsi-Farkas, C. History and future of food irradiation. Trends Food Sci. Technol. 2011, 22, 121–126. [Google Scholar] [CrossRef]
- Praveen, C.; Dancho, B.A.; Kingsley, D.H.; Calci, K.R.; Meade, G.K.; Mena, K.D.; Pillai, S.D. Susceptibility of murine norovirus and hepatitis a virus to electron beam irradiation in oysters and quantifying the reduction in potential infection risks. Appl. Environ. Microbiol. 2013, 79, 3796–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IAEA. Manual of Good Practice in Food Irradiation: Sanitary, Phytosanitary and Other Applications; Technical Reports Series; International Atomic Energy Agency: Vienna, Austria, 2015; ISBN 978-92-0-105215-5. [Google Scholar]
- FDA. Sec. 179.26 Ionizing Radiation for the Treatment of Food 2019; FDA: Washington, DC, USA, 2019.
- Lung, H.-M.; Cheng, Y.-C.; Chang, Y.-H.; Huang, H.-W.; Yang, B.B.; Wang, C.-Y. Microbial decontamination of food by electron beam irradiation. Trends Food Sci. Technol. 2015, 44, 66–78. [Google Scholar] [CrossRef]
- Bhat, R.; Alias, A.K.; Paliyath, G. Use of electron beams in food preservation. In Progress in Food Preservation; John Wiley & Sons, Ltd.: Chichester, UK, 2012; pp. 343–372. ISBN 978-1-119-96204-5. [Google Scholar]
- Shayanfar, S.; Pillai, S.D. Electron beam processing of foods. In Non-Thermal Processing of Foods; Chauhan, O.P., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 315–328. ISBN 978-1-351-86977-5. [Google Scholar]
- Smith, B.; Shayanfar, S.; Walzem, R.; Alvarado, C.Z.; Pillai, S.D. Preserving fresh fruit quality by low-dose electron beam processing for vending distribution channels. Radiat. Phys. Chem. 2020, 168, 108540. [Google Scholar] [CrossRef]
- Gryczka, U.; Migdał, W.; Bułka, S. The effectiveness of the microbiological radiation decontamination process of agricultural products with the use of low energy electron beam. Radiat. Phys. Chem. 2018, 143, 59–62. [Google Scholar] [CrossRef]
- Gryczka, U.; Kameya, H.; Kimura, K.; Todoriki, S.; Migdał, W.; Bułka, S. Efficacy of low energy electron beam on microbial decontamination of spices. Radiat. Phys. Chem. 2020, 170, 108662. [Google Scholar] [CrossRef]
- Ben Salem, I.; Ouesleti, S.; Khammassi, M.A.; Boulila, A.; Mabrouk, Y. Effect of ionizing radiation on the microbiological safety and phytochemical properties of cooked malva sylvestris L. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Kalaiselvan, R.R.; Sugumar, A.; Radhakrishnan, M. Chapter 21—Gamma irradiation usage in fruit juice extraction. In Fruit Juices; Rajauria, G., Tiwari, B.K., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 423–435. ISBN 978-0-12-802230-6. [Google Scholar]
- Obana, H.; Furuta, M.; Tanaka, Y. Detection of 2-alkylcyclobutanones in irradiated meat, poultry and egg after cooking. J. Health Sci. 2006, 52, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Solhaug, K.A.; Song, Y.; Brede, D.A.; Lind, O.C.; Salbu, B.; Tollefsen, K.E. Modes of action and adverse effects of gamma radiation in an aquatic macrophyte Lemna minor. Sci. Total Environ. 2019, 680, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Cerf, O. Rappels des bases théoriques de l’inactivation des microorganismes. In Traitement Ionisants et Hautes Pressions des Aliments; Federighi, M., Tholozan, J.L., Eds.; Economica: Paris, France, 2001; pp. 1–20. ISBN 2-7178-4162-8. [Google Scholar]
- Szczawińska, M.E. Application of ionizing radiation for control of salmonella in food. Curr. Top. Salmonella Salmonellosis 2017. [Google Scholar] [CrossRef] [Green Version]
- Obodovskiy, I. Radiation sterilization. In Radiation: Fundamentals, Applications, Risks, and Safety; Obodovskiy, I., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 373–378. ISBN 978-0-444-63979-0. [Google Scholar]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: Extremophiles and the limits of life in a planetary context. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Huber, N.; Moeller, R.; Stülke, J.; Dubovcova, B.; Akepsimaidis, G.; Meneses, N.; Drissner, D.; Mathys, A. Role of DNA repair in Bacillus subtilis spore resistance to high energy and low energy electron beam treatments. Food Microbiol. 2020, 87, 103353. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Venkateswaran, A.; Hess, M.; Omelchenko, M.V.; Kostandarithes, H.M.; Makarova, K.S.; et al. Accumulation of Mn(II) in deinococcus radiodurans facilitates gamma-radiation resistance. Science 2004, 306, 1025–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosal, D.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Venkateswaran, A.; Zhai, M.; Kostandarithes, H.M.; Brim, H.; Makarova, K.S.; et al. How radiation kills cells: Survival of Deinococcus radiodurans and Shewanella oneidensis under oxidative stress. Fems. Microbiol. Rev. 2005, 29, 361–375. [Google Scholar] [CrossRef] [Green Version]
- Josephson, P. Preservation of Food by Ionizing Radiation: Volume III; CRC Press: Boca Raton, FL, USA, 2018; ISBN 978-1-351-09289-0. [Google Scholar]
- Sommers, C.H.; Boyd, G. Radiation sensitivity and postirradiation growth of foodborne pathogens on a ready-to-eat frankfurter on a roll product in the presence of modified atmosphere and antimicrobials. J. Food Prot. 2006, 69, 2436–2440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Moeller, R.; Tran, S.; Dubovcova, B.; Akepsimaidis, G.; Meneses, N.; Drissner, D.; Mathys, A. Geobacillus and bacillus spore inactivation by low energy electron beam technology: Resistance and influencing factors. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Xu, A.; Scullen, O.J.; Sheen, S.; Johnson, J.R.; Sommers, C.H. Inactivation of extraintestinal pathogenic E. coli clinical and food isolates suspended in ground chicken meat by gamma radiation. Food Microbiol. 2019, 84, 103264. [Google Scholar] [CrossRef] [PubMed]
- Adu-Gyamfi, A.; Nketsia-Tabiri, J.; Boatin, R. Determination of DB10B values of single and mixed cultures of bacteria after gamma irradiation. J. Appl. Sci. Technol. 2009, 14. [Google Scholar] [CrossRef]
- Gaougaou, G.; Ben-Fadhel, Y.; Déziel, E.; Lacroix, M. Effect of β-lactam antibiotic resistance gene expression on the radio-resistance profile of E. coli O157:H7. Heliyon 2018, 4, e00999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levanduski, L.; Jaczynski, J. Increased resistance of Escherichia coli O157:H7 to electron beam following repetitive irradiation at sub-lethal doses. Int. J. Food Microbiol. 2008, 121, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Tesfai, A.T.; Beamer, S.K.; Matak, K.E.; Jaczynski, J. Radioresistance development of DNA repair deficient Escherichia coli DH5α in ground beef subjected to electron beam at sub-lethal doses. Int. J. Radiat. Biol. 2011, 87, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Grudlewska, K.; Gryń, G.; Skowron, K.J.; Świeca, A.; Paluszak, Z.; Zimek, Z.; Rafalski, A.; Gospodarek-Komkowska, E. Effect of electron beam and gamma radiation on drug-susceptible and drug-resistant Listeria monocytogenes strains in salmon under different temperature. J. Appl. Microbiol. 2018, 125, 828–842. [Google Scholar] [CrossRef]
- Muñoz, A.A. Efectos de Tratamientos no Térmicos y de Bioconservación en Jamón Curado Loncheado Sobre La Expresión Génica de “Listeria Monocytogenes”. Ph.D. Thesis, University of Extremadura, Badajoz, Spain, 2019. [Google Scholar]
- Dutra, M.P.; de Aleixo, G.C.; de Ramos, A.L.S.; Silva, M.H.L.; Pereira, M.T.; Piccoli, R.H.; Ramos, E.M. Use of gamma radiation on control of Clostridium botulinum in mortadella formulated with different nitrite levels. Radiat. Phys. Chem. 2016, 119, 125–129. [Google Scholar] [CrossRef]
- Sukhi, S.S.; Shashidhar, R.; Kumar, S.A.; Bandekar, J.R. Radiation resistance of Deinococcus radiodurans R1 with respect to growth phase. Fems. Microbiol. Lett. 2009, 297, 49–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendonca, A.F.; Romero, M.G.; Lihono, M.A.; Nannapaneni, R.; Johnson, M.G. Radiation resistance and virulence of listeria monocytogenes scott a following starvation in physiological saline. J. Food Prot. 2004, 67, 470–474. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Edelson-Mammel, S.G.; Boyd, G.; Marmer, B.S. Influence of acidulant identity on the effects of pH and acid resistance on the radiation resistance of Escherichia coli O157:H7. Food Microbiol. 2004, 21, 51–57. [Google Scholar] [CrossRef]
- Osaili, T.M.; Al-Nabulsi, A.A.; Abubakar, S.A.; Alaboudi, A.R.; Al-Holy, M.A. Feasibility of using gamma irradiation for inactivation of starvation-, heat-, and cold-stressed salmonella in tahini. J. Food Prot. 2016, 79, 963–969. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Ruppitsch, W.; Pekard-Amenitsch, S.; Forsythe, S.J.; Cormican, M.; Mach, R.L.; Piérard, D.; Allerberger, F. Multicenter study of cronobacter sakazakii infections in humans, Europe, 2017. Emerg. Infect. Dis. 2019, 25, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-W.; Oh, S.-H.; Byun, E.-B.; Kim, J.-H.; Kim, J.-H.; Woon, J.-H.; Byun, M.-W. Inactivation of Enterobacter sakazakii of dehydrated infant formula by gamma-irradiation. Radiat. Phys. Chem. 2007, 76, 1858–1861. [Google Scholar] [CrossRef]
- Osaili, T.M.; Shaker, R.R.; Abu Al-Hasan, A.S.; Ayyash, M.M.; Martin, E.M. Inactivation of enterobacter sakazakii in infant milk formula by gamma irradiation: Determination of D10-Value. J. Food Sci. 2007, 72, M85–M88. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.-H.; Park, J.-Y.; Park, J.-H.; Chung, M.-S.; Kwon, K.-S.; Chung, K.; Won, M.; Song, K.-B. Inactivation of Enterobacter sakazakii, Bacillus cereus, and Salmonella typhimurium in powdered weaning food by electron-beam irradiation. Radiat. Phys. Chem. 2008, 77, 1097–1100. [Google Scholar] [CrossRef]
- Etter, D.; Rupp, A.; Prange, A.; Drissner, D. Inactivation of mould spores in a model system and on raisins by low-energy electron beam. Food Control. 2018, 92, 357–361. [Google Scholar] [CrossRef]
- Kawasaki, S.; Saito, M.; Mochida, M.; Noviyanti, F.; Seito, H.; Todoriki, S. Inactivation of Escherichia coli O157 and Salmonella Enteritidis in raw beef liver by gamma irradiation. Food Microbiol. 2019, 78, 110–113. [Google Scholar] [CrossRef]
- Demonts, M.H.; Strasser, A.; Hasselmann, C.; Marchionie, E.; Hausser, F. Les traitements ionisants des aliments. In Traitement Ionisants et Hautes Pressions des Aliments; Federighi, M., Tholozan, J.L., Eds.; Economica: Paris, France, 2001; pp. 1–20. ISBN 2-7178-4162-8. [Google Scholar]
- Hume, A.J.; Ames, J.; Rennick, L.J.; Duprex, W.P.; Marzi, A.; Tonkiss, J.; Mühlberger, E. Inactivation of RNA viruses by gamma irradiation: A study on mitigating factors. Viruses 2016, 8, 204. [Google Scholar] [CrossRef]
- Bhatia, S.S.; Pillai, S.D. A comparative analysis of the metabolomic response of electron beam inactivated E. coli O26:H11 and salmonella typhimurium ATCC 13311. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemira, B.A.; Fan, X.; Sokorai, K.J.B.; Sommers, C.H. Ionizing radiation sensitivity of listeria monocytogenes ATCC 49594 and listeria innocua ATCC 51742 inoculated on endive (cichorium endiva). J. Food Prot. 2003, 66, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Karagöz, I.; Moreira, R.G.; Castell-Perez, M.E. Radiation D10 values for Salmonella Typhimurium LT2 and an Escherichia coli cocktail in pecan nuts (Kanza cultivar) exposed to different atmospheres. Food Control. 2014, 39, 146–153. [Google Scholar] [CrossRef]
- Yurttas, Z.S.; Moreira, R.G.; Castell-Perez, E. Combined vacuum impregnation and electron-beam irradiation treatment to extend the storage life of sliced white button mushrooms (Agaricus bisporus). J. Food Sci. 2014, 79, E39–E46. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.G.; Puerta-Gomez, A.F.; Kim, J.; Castell-Perez, M.E. Factors affecting radiation D-values (D10) of an escherichia coli cocktail and salmonella typhimurium LT2 inoculated in fresh produce. J. Food Sci. 2012, 77, E104–E111. [Google Scholar] [CrossRef] [PubMed]
- Kudra, L.L.; Sebranek, J.G.; Dickson, J.S.; Larson, E.M.; Mendonca, A.F.; Prusa, K.J.; Cordray, J.C.; Jackson-Davis, A.; Lu, Z. Control of listeria monocytogenes on frankfurters and cooked pork chops by irradiation combined with modified atmosphere packaging. J. Food Prot. 2012, 75, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, C.P. High-dose irradiated food: Current progress, applications, and prospects. Radiat. Phys. Chem. 2018, 144, 34–36. [Google Scholar] [CrossRef]
- Kortei, N.K.; Odamtten, G.T.; Obodai, M.; Wiafe-Kwagyan, M. Mycofloral profile and the radiation sensitivity (D10 values) of solar dried and gamma irradiated Pleurotus ostreatus (Jacq.Ex. Fr.) Kummer fruitbodies stored in two different packaging materials. Food Sci. Nutr. 2017, 6, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Kortei, N.K.; Odamtten, G.T.; Appiah, V.; Obodai, M.; Adu-Gyamfi, A.; Annan, T.A.; Akonor, P.T.; Annan, S.N.Y.; Acquah, S.A.; Armah, J.O.; et al. Microbiological quality assessment of gamma irradiated fresh and dried mushrooms (Pleurotus ostreatus) and determination of D10 values of Bacillus cereus in storage packs. Eur. J. Biotechnol. Biosci. 2014, 2, 28–34. [Google Scholar]
- Haji-Saeid, M.; Sampa, M.H.O.; Chmielewski, A.G. Radiation treatment for sterilization of packaging materials. Radiat. Phys. Chem. 2007, 76, 1535–1541. [Google Scholar] [CrossRef]
- Li, S.; Kundu, D.; Holley, R.A. Use of lactic acid with electron beam irradiation for control of Escherichia coli O157:H7, non-O157 VTEC E. coli, and Salmonella serovars on fresh and frozen beef. Food Microbiol. 2015, 46, 34–39. [Google Scholar] [CrossRef]
- Huq, T.; Vu, K.D.; Riedl, B.; Bouchard, J.; Lacroix, M. Synergistic effect of gamma (γ)-irradiation and microencapsulated antimicrobials against Listeria monocytogenes on ready-to-eat (RTE) meat. Food Microbiol. 2015, 46, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Modi, N.K.; Rose, S.A.; Tranter, H.S. The effects of irradiation and temperature on the immunological activity of staphylococcal enterotoxin A. Int. J. Food Microbiol. 1990, 11, 85–92. [Google Scholar] [CrossRef]
- Rose, S.A.; Modi, N.K.; Tranter, H.S.; Bailey, N.E.; Stringer, M.F.; Hambleton, P. Studies on the irradiation of toxins of Clostridium botulinum and Staphylococcus aureus. J. Appl. Bacteriol. 1988, 65, 223–229. [Google Scholar] [CrossRef]
- Jo, C.; Lee, N.Y.; Kang, H.; Hong, S.; Kim, Y.; Kim, H.J.; Byun, M.W. Radio-sensitivity of pathogens in inoculated prepared foods of animal origin. Food Microbiol. 2005, 22, 329–336. [Google Scholar] [CrossRef]
- Umar, S.; Younus, M.; Rehman, M.U.; Aslam, A.; Shah, M.A.A.; Munir, M.T.; Hussain, S.; Iqbal, F.; Fiaz, M.; Ullah, S. Role of aflatoxin toxicity on transmissibility and pathogenicity of H9N2 avian influenza virus in turkeys. Avian Pathol. 2015, 44, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Umar, S.; Munir, M.T.; Muhammad, A.S.; Shahzad, M.; Khan, R.A.; Sohoo, M.; Khan, A.U.; Ameen, K.; Rafia-Munir, A. Farrukh Saleem Outbreak of aflatoxicosis on a local cattle farm in Pakistan. Veterinaria 2015, 3, 13–17. [Google Scholar]
- Munir, M.T.; Rehman, Z.U.; Shah, M.A.; Umar, S. Interactions of Aspergillus fumigatus with the respiratory system in poultry. World’s Poult. Sci. J. 2017, 73, 321–336. [Google Scholar] [CrossRef]
- Kaboudi, K.; Rejeb, A.; Bouzouaia, M.; Munir, M.; Sajid, U. Outbreak of respiratory aspergillosis in backyard duck flock in Tunisia. Int. J. Livest. Res. 2018, 8, 361. [Google Scholar] [CrossRef]
- Calado, T.; Fernández-Cruz, M.L.; Cabo Verde, S.; Venâncio, A.; Abrunhosa, L. Gamma irradiation effects on ochratoxin A: Degradation, cytotoxicity and application in food. Food Chem. 2018, 240, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Domijan, A.-M.; Pleadin, J.; Mihaljević, B.; Vahčić, N.; Frece, J.; Markov, K. Reduction of ochratoxin A in dry-cured meat products using gamma-irradiation. Food Addit. Contam. Part A 2015, 32, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Hassanpour, M.; Rezaie, M.R.; Baghizadeh, A. Practical analysis of aflatoxin M1 reduction in pasteurized Milk using low dose gamma irradiation. J. Environ. Health Sci. Eng. 2019, 17, 863–872. [Google Scholar] [CrossRef]
- Kottapalli, B.; Wolf-Hall, C.E.; Schwarz, P. Effect of electron-beam irradiation on the safety and quality of Fusarium-infected malting barley. Int. J. Food Microbiol. 2006, 110, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Domijan, A.-M.; Čermak, A.M.M.; Vulić, A.; Bujak, I.T.; Pavičić, I.; Pleadin, J.; Markov, K.; Mihaljević, B. Cytotoxicity of gamma irradiated aflatoxin B1 and ochratoxin A. J. Environ. Sci. Health Part B 2019, 54, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuzaudi, M.; Eerikäinen, T.; Turunen, O.; Ghelawi, M.; Assad, M.E.H.; Tawalbeh, M.; Bedade, D.; Shamekh, S. Effect of gamma irradiation and heat treatment on the artificial contamination of maize grains by Aspergillus flavus Link NRRL 5906. J. Stored Prod. Res. 2017, 71, 57–63. [Google Scholar] [CrossRef]
- Nurtjahja, K.; Dharmaputra, O.S.; Rahayu, W.P.; Syarief, R. Gamma irradiation of Aspergillus flavus strains associated with Indonesian nutmeg (Myristica fragrans). Food Sci. Biotechnol. 2017, 26, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Al-Abdalall, A.H.A. Inhibitory effect of gamma radiation in degrading and preventing fungal toxins. J. Food Agric. Environ. 2014, 12, 77–81. [Google Scholar]
- DiCaprio, E.; Phantkankum, N.; Culbertson, D.; Ma, Y.; Hughes, J.H.; Kingsley, D.; Uribe, R.M.; Li, J. Inactivation of human norovirus and Tulane virus in simple media and fresh whole strawberries by ionizing radiation. Int. J. Food Microbiol. 2016, 232, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, A.I.; Margaça, F.M.A.; Cabo Verde, S. Virucidal activity of gamma radiation on strawberries and raspberries. Int. J. Food Microbiol. 2019, 304, 89–96. [Google Scholar] [CrossRef]
- Miekka, S.I.; Forng, R.-Y.; Rohwer, R.G.; MacAuley, C.; Stafford, R.E.; Flack, S.L.; MacPhee, M.; Kent, R.S.; Drohan, W.N. Inactivation of viral and prion pathogens by γ-irradiation under conditions that maintain the integrity of human albumin. Vox Sang. 2003, 84, 36–44. [Google Scholar] [CrossRef]
- Bidawid, S.; Farber, J.M.; Sattar, S.A. Inactivation of hepatitis A virus (HAV) in fruits and vegetables by gamma irradiation. Int. J. Food Microbiol. 2000, 57, 91–97. [Google Scholar] [CrossRef]
- Smolko, E.E.; Lombardo, J.H. Virus inactivation studies using ion beams, electron and gamma irradiation. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2005, 236, 249–253. [Google Scholar] [CrossRef]
- Feng, K.; Divers, E.; Ma, Y.; Li, J. Inactivation of a human norovirus surrogate, human norovirus virus-like particles, and vesicular stomatitis virus by gamma irradiation. Appl. Environ. Microbiol. 2011, 77, 3507–3517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, F.; Shupert, W.L.; Haddock, E.; Twardoski, B.; Feldmann, H. Gamma irradiation as an effective method for inactivation of emerging viral pathogens. Am. J. Trop. Med. Hyg. 2019, 100, 1275–1277. [Google Scholar] [CrossRef] [PubMed]
- Gominet, M.; Vadrot, C.; Austruy, G.; Darbord, J.C. Inactivation of prion infectivity by ionizing rays. Radiat. Phys. Chem. 2007, 76, 1760–1762. [Google Scholar] [CrossRef]
- Sullivan, R.; Fassolitis, A.C.; Larkin, E.P.; Read, R.B.; Peeler, J.T. Inactivation of thirty viruses by gamma radiation. Appl. Environ. Microbiol. 1971, 22, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Kaboudi, K.; Umar, S.; Munir, M.T. Prevalence of coccidiosis in free-range chicken in Sidi Thabet, Tunisia. Scientifica 2016, 7075195. [Google Scholar] [CrossRef] [Green Version]
- Pohle, S.; Ernst, R.; MacKenzie, C.; Spicher, M.; Romig, T.; Hemphill, A.; Gripp, S. Echinococcus multilocularis: The impact of ionizing radiation on metacestodes. Exp. Parasitol. 2011, 127, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.R.; Park, W.Y. The effect of γ-irradiation on the viability of cryptosporidium parvum. J. Parasitol. 2003, 89, 639–642. [Google Scholar] [CrossRef]
- Franssen, F.; Gerard, C.; Cozma-Petruţ, A.; Vieira-Pinto, M.; Jambrak, A.R.; Rowan, N.; Paulsen, P.; Rozycki, M.; Tysnes, K.; Rodriguez-Lazaro, D.; et al. Inactivation of parasite transmission stages: Efficacy of treatments on food of animal origin. Trends Food Sci. Technol. 2019, 83, 114–128. [Google Scholar] [CrossRef]
- Hallman, G.J. Control of stored product pests by ionizing radiation. J. Stored Prod. Res. 2013, 52, 36–41. [Google Scholar] [CrossRef]
- Codex Alimentarius. Codex Alimentarius Code of Practice for Radiation Processing of Food (CAC/RCP 19–1979, Rev.2–2003b); Editorial Correction 2011; Food Standards Program; Secretariat of the Codex Alimentarius Commission; FAO/WHO: Rome, Italy, 2011. [Google Scholar]
- Codex Alimentarius. Codex Alimentarius General Standard for Irradiated Foods (Codex Standard 106–1983, Rev.1–2003a); Editorial Correction 2011; Food Standards Program; Secretariat of the Codex Alimentarius Commission; FAO/WHO: Rome, Italy, 2011. [Google Scholar]
- European Parliament and of the Council. Directive 1999/3/EC of the European Parliament and of the Council of 22 February 1999 on the Establishment of a Community List of Foods and Food Ingredients Treated with Ionising Radiation; European Parliament and of the Council: Brussels, Belgium, 1999; L 66/24. [Google Scholar]
- Junqueira-Gonçalves, M.P.; Galotto, M.J.; Valenzuela, X.; Dinten, C.M.; Aguirre, P.; Miltz, J. Perception and view of consumers on food irradiation and the Radura symbol. Radiat. Phys. Chem. 2011, 80, 119–122. [Google Scholar] [CrossRef]
- Rahman, M.H.; Islam, M.S.; Begum, S.; Ali, M.L.; Sutradhar, B.C.; O’neil, V.; Hossain, A.; Nandwa, C. Scientific opinion on standards and regulations of irradiated food. J. Nutr. Food Sci. 2018, 8, 2. [Google Scholar] [CrossRef]
- Aymerich, T.; Picouet, P.A.; Monfort, J.M. Decontamination technologies for meat products. Meat Sci. 2008, 78, 114–129. [Google Scholar] [CrossRef] [PubMed]
- Pedreschi, F.; Mariotti-Celis, M.S. Irradiation kills microbes. In Genetically Modified and Irradiated Food; Elsevier: Amsterdam, The Netherlands, 2020; pp. 233–242. ISBN 978-0-12-817240-7. [Google Scholar]
- Bearth, A.; Siegrist, M. “As long as it is not irradiated”—Influencing factors of US consumers’ acceptance of food irradiation. Food Qual. Prefer. 2019, 71, 141–148. [Google Scholar] [CrossRef]
- Yamakage, K.; Sui, H.; Ohta, R.; Toyoizumi, T.; Kawakami, K.; Matsumoto, H.; Takahashi, T.; Sasaki, K.; Ikezumi, M.; Negishi, S.; et al. Genotoxic potential and in vitro tumour-promoting potential of 2-dodecylcyclobutanone and 2-tetradecylcyclobutanone, two radiolytic products of fatty acids. Mutat. Res. Toxicol. Environ. Mutagen. 2014, 770, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.B. Food irradiation is safe: Half a century of studies. Radiat. Phys. Chem. 2014, 105, 78–82. [Google Scholar] [CrossRef]
- Maherani, B.; Hossain, F.; Criado, P.; Ben-Fadhel, Y.; Salmieri, S.; Lacroix, M. World market development and consumer acceptance of irradiation technology. Foods 2016, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Galati, A.; Moavero, P.; Crescimanno, M. Consumer awareness and acceptance of irradiated foods: The case of Italian consumers. Br. Food J. 2019, 121, 1398–1412. [Google Scholar] [CrossRef]
- Galati, A.; Tulone, A.; Moavero, P.; Crescimanno, M. Consumer interest in information regarding novel food technologies in Italy: The case of irradiated foods. Food Res. Int. 2019, 119, 291–296. [Google Scholar] [CrossRef]
- Shahbaz, H.M.; Akram, K.; Ahn, J.-J.; Kwon, J.-H. Worldwide status of fresh fruits irradiation and concerns about quality, safety, and consumer acceptance. Crit. Rev. Food Sci. Nutr. 2016, 56, 1790–1807. [Google Scholar] [CrossRef]




| Species | Genome Size (Mb) | Number of Cytochrome C | Intracellular Mn/Fe Ratio | DRD (kGy) |
|---|---|---|---|---|
| Deinococcus radiodurans | 3.28 | 7 | 0.24 | 10–12 |
| D. geothermalis | 3.23 | 7 | 0.46 | 10 |
| Escherichia coli | 4.64 | 6 | 0.007 | 0.7 |
| Pseudomonas putida | 6.18 | 17 | <0.001 | 0.25 |
| Shewanella odeinensis | 5.13 | 39 | <0.001 | 0.07 |
| Species | Single Culture | Mix Culture |
|---|---|---|
| Escherichia coli | 0.27 | 0.24 |
| Staphylococcus aureus | 0.33 | 0.28 |
| Salmonella Paratyphi B | 0.44 | 0.32 |
| Medium | Bone Marrow | Corned Beef | Whole Egg | Meat | Kaolin Clay | Phosphate Buffer |
|---|---|---|---|---|---|---|
| DRD | 0.91 | 0.8 | 0.632 | 0.558 | 0.21 | 0.208 |
| Treatment Media | ||
|---|---|---|
| Numeration Media | Phosphate Buffer | Chicken Meat |
| Listeria selective agar | 0.324 | 0.424 |
| Tryptone soya levure agar | 0.404 | 0.481 |
| DRD Values (kGy) | ||
|---|---|---|
| Treatment Media | At Room Temperature | At Frozen Temperature (−15 °C) |
| Phosphate buffer | 0.208 | 0.391 |
| Meat | 0.558 | 0.963 |
| Dehydrated coconut | 1.58 | - |
| Vacuum-Packed | Nitrogen-Packed | Oxygen-Packed | Air-Packed | |
|---|---|---|---|---|
| Salmonella Typhimurium | 0.44 ± 0.06 | 0.38 ± 0.05 | 0.34 ± 0.04 | 0.36 ± 0.01 |
| Escherichia coli | 0.46 ± 0.05 | 0.40 ± 0.04 | 0.36 ± 0.05 | 0.38 ± 0.005 |
| Yeasts | DRD (kGy) | Dose 1 (kGy) | Treatment Media |
|---|---|---|---|
| Candida krusei | - | 5.5 | Buffer |
| Candida spp | 1.25 | - | Nutrient broth |
| Candida tropicales | - | 10 | Buffer |
| Candida zelyanoides | 0.7 | - | Chicken skin |
| Cryptococcus albidus | - | 10 | Grape juice |
| Debaryomyces klöeckeri | - | 7.5 | Grape juice |
| Rhodotorula glutinis | - | 10 | Grape juice |
| Molds | DRD (kGy) | Treatment Media | Dose 1 (kGy) | Treatment Media |
|---|---|---|---|---|
| Alternaria spp | 6 | Nutrient agar | ||
| Alternaria citri | 1.2 | Lemon | ||
| Aspergillus flavus | - | - | 5 | Peanuts |
| A. flavus (spores) | 0.4 | Water | 1.7 | Nutrient broth |
| A. flavus | - | - | 6 | Sesam seed |
| Aspergillus candidus | 0.3 | - | - | - |
| Aspergillus niger | 0.45 | Water | 2.5 | Nutrient agar |
| Botrytis cinerea | 0.6 | Plum | 5 | Nutrient agar |
| Cladosporium herbarum | 1 | Plum | 6 | Nutrient agar |
| Geotrichum candidum | 0.4 | Lemon | - | - |
| Penicillium camenbertii | 0.2 | Water | - | - |
| Penicillium expansum | 0.4 | Plum | - | - |
| Penicillium roquefortii | 0.4 | - | - | - |
| Penicillium viridicatum | - | - | 1.4 | Nutrient agar |
| Rhizopus nigricans | 0.7 | Plum | 2.5 | Nutrient agar |
| Viruses | DRD (kGy) | Treatment Media | Temperature | References |
|---|---|---|---|---|
| FMD 1 virus types A and C | 6 | Calf kidney | NI | [49] |
| FMD virus type D | 5 | Calf kidney | NI | [49] |
| FMD virus type O | 6 | Calf kidney | −60 °C | [49] |
| FMD virus (A, C, O) | 2.7 | Buffer | <0 °C | [82] |
| TESHEN disease virus | 4.3 | 20% peptone solution | NI | [49] |
| Rinderpest virus | <6 | NI 2 | −20 °C | [49] |
| Swine fever virus | <6 | NI | −20 °C | [49] |
| Hepatitis A virus | 2.72 | Lettuce | +4 °C | [81] |
| Hepatitis A virus | 2.92 | Strawberry | +4 °C | [81] |
| Hepatitis A virus | 4.83 | Oysters | 0 °C | [8] |
| Hepatitis A virus | 5.74 | Oyster homogenate | 0 °C | [8] |
| Human norovirus | 4.05 | Oysters | 0 °C | [8] |
| Human norovirus | 4.97 | Oyster homogenate | 0 °C | [8] |
| Human norovirus | 2.55 | PBS 3 | 0 °C | [8] |
| Adenovirus | 4.2–5 | MEM 4 + FBS 5 | 0 °C | [86] |
| Coxsackievirus | 4.2–5 | MEM 4 + FBS 5 | 0 °C | [86] |
| Echovirus | 4.3–5.5 | MEM + FBS | 0 °C | [86] |
| Poliovirus | 4.3–5.2 | MEM 4 + FBS 5 | 0 °C | [86] |
| Herpes simplex virus | 2.92 (β) | Buffer | <0 °C | [82] |
| Herpes simplex virus | 1.1–1.3 (γ) | Buffer | <0 °C | [82] |
| Rauscher Leukemia Virus | 0.6–2.1 | Buffer | <0 °C | [82] |
| Influenza A virus | 4.7 | MEM 4 + FBS 5 | 0 °C | [86] |
| Rabies virus | 1 | 10% brain emulsion | NI | [49] |
| Vesicular stomatitis virus | 2.7 | DMEM 6 | −78 °C | [50] |
| La Crosse virus | 2.6 | DMEM | −78 °C | [50] |
| Measles virus | 2.5 | DMEM | −78 °C | [50] |
| Measles virus | 2.7 | DMEM | −78 °C | [50] |
| Parasite | Foods | Parasitic Form | Sterility Dose |
|---|---|---|---|
| Ascoris lumbricoïdes | Raw vegetables | Egg | 1 to 48 |
| Anisakis marina | Mackerel | Larvae | 3 to 6 |
| Cysticercus bovis | Beef | Larvae | 0.2 to 0.6 |
| Cysticercus cellulosae | Pork | Destruction | 2.8 to 10 |
| Fasciola hepatica | Water, vegetables | Cysts forms | 0.03 |
| Fasciola hepatica | Larvae | 1.9 | |
| Fasciola hepatica | Egg | 4.8 | |
| Hymenolepsis nana | Cereals | Larvae | 0.3 to 0.4 |
| Trichinella spiralis | Pork | Larvae | 0.03 to 0.09 |
| Trichinella spiralis | Destruction | 1.4 to 6.3 |
| Insects Species | Sterilization Dose for Insects | References | |
|---|---|---|---|
| Male (kGy) | Female (kGy) | ||
| Attagenus unicolor (Brahm) | 0.175 | 0.175 | |
| Callosobruchus maculatus | 0.07 | 0.07 | |
| Lasioderma serricorne | 0.25 | 0.175 | |
| Latheticus oryzae | 0.2 | 0.1 | |
| Sitophilus granarius | 0.1 | 0.1 | |
| Sitrotoga cereallela | 0.45 | 0.45 | |
| Tenebrio molitor | 0.15 | 0.05 | |
| Tribolium castaneum | 0.2 | 0.2 | |
| Trogoderma glabrum | 0.25 | 0.132 | |
| Dose (kGy) | Usage for Food Products |
|---|---|
| 0.05–0.15 | Anti-germination for potatoes, onions, garlic, shallots, etc. |
| 0.15–3 | Pesticide and insecticide properties (by the destruction of embryos and larvae, and sterilization of adults) for cereals, fruits, and pulses. In this dose range, some meat parasitic worms, e.g., Trichinella in pork, can be eliminated, and also the ripening of fresh fruits and vegetables is delayed. |
| 2–10 | Treatment of meat and meat products, ready meals, fish products (fish, shellfish, frog’s legs), fresh fruit and vegetables, spices and herbs, and various ingredients (gums, additives) to improve and guarantee a hygienic quality by eliminating pathogenic microorganisms and/or extend their shelf life by reducing the microbial population of spoilage. |
| 10–60 | Food is sterilized and then can be stored for up to 2 years at room temperature in sealed plastic packaging |
| Foods | Dose (kGy) | Temperature |
|---|---|---|
| Bacon | 25.2 | Room |
| Beef | 41.2 | −30 °C |
| Corned beef | 26.9 | −30 °C |
| Duck with orange sauce | 21 | −40 °C |
| Turkey | 28 | −40 °C |
| Ham | 31.4 | −30 °C |
| Pork | 43.7 | −30 °C |
| Pork sausages | 25.5 | −30 °C |
| Chicken | 42.7 | −30 °C |
| Mashed carrots and sweet potatoes | 28 | −40 °C |
| Madras rice | 28 | −40 °C |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munir, M.T.; Federighi, M. Control of Foodborne Biological Hazards by Ionizing Radiations. Foods 2020, 9, 878. https://doi.org/10.3390/foods9070878
Munir MT, Federighi M. Control of Foodborne Biological Hazards by Ionizing Radiations. Foods. 2020; 9(7):878. https://doi.org/10.3390/foods9070878
Chicago/Turabian StyleMunir, Muhammad Tanveer, and Michel Federighi. 2020. "Control of Foodborne Biological Hazards by Ionizing Radiations" Foods 9, no. 7: 878. https://doi.org/10.3390/foods9070878
APA StyleMunir, M. T., & Federighi, M. (2020). Control of Foodborne Biological Hazards by Ionizing Radiations. Foods, 9(7), 878. https://doi.org/10.3390/foods9070878