Antidiabetic Food-Derived Peptides for Functional Feeding: Production, Functionality and In Vivo Evidences




Abstract
:1. Introduction
1.1. Proteins, Protein Hydrolysates and Peptides
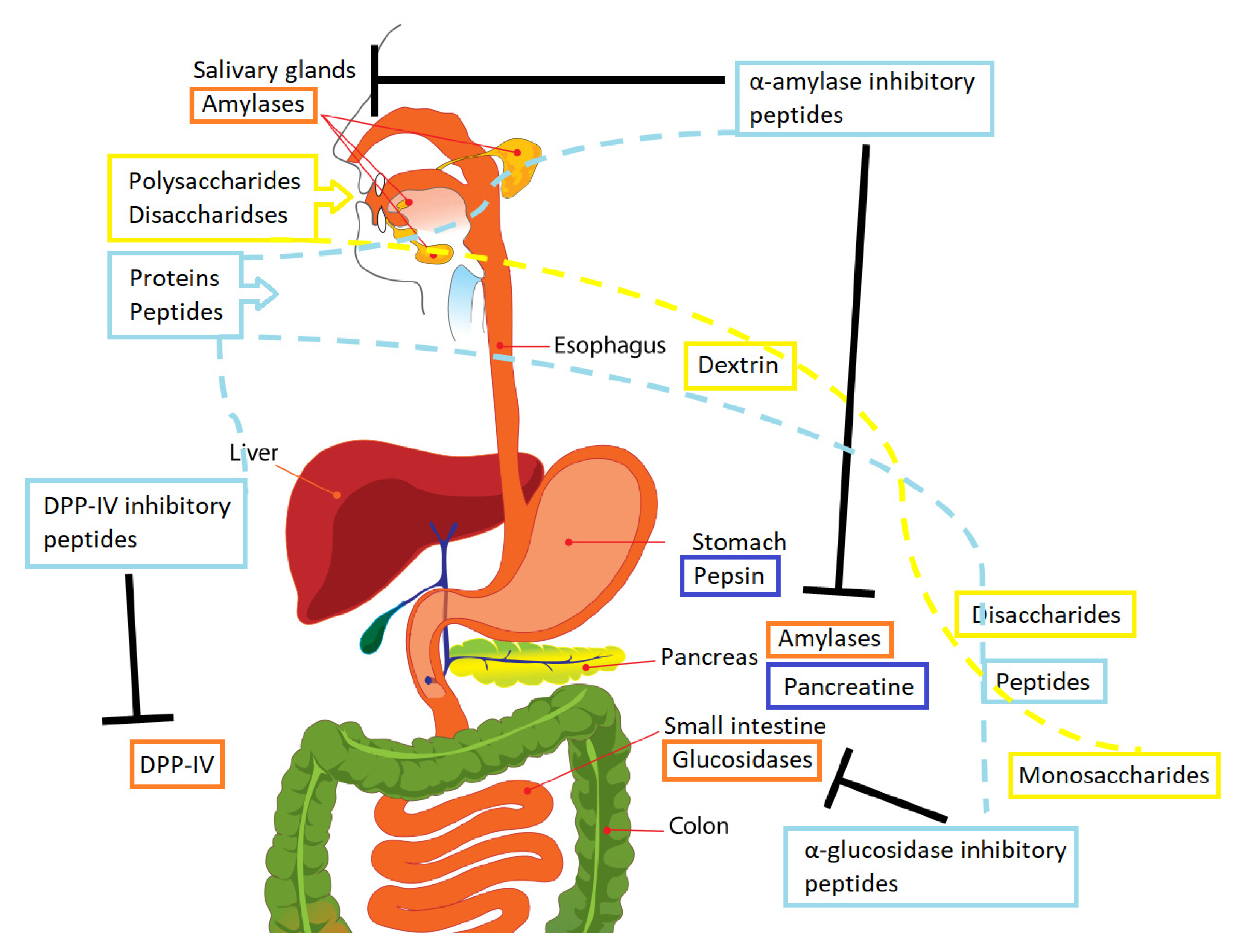
1.2. Carbohydrates Digestion Process and Diabetes
1.3. Diabetes Prevention Strategies
2. Production of Glycemic Index-Regulating Protein Hydrolysates
2.1. Enzymatic Hydrolysis Reaction
2.2. Proteases
2.3. Protein Source
3. Identification of Bioactive Peptides
3.1. Fractionation
3.2. Peptide Sequence Identification
3.3. Bioinformatics Analysis
4. Bioavailability In Vitro
5. Stability and Functionality in Food Matrices
6. Bioactivity Analysis
6.1. Bioactivity Initial Approaches
- -
- BRIN-BD11: insulin-secreting cells (pancreatic B cells) in response to glucose, to analyze the effect of the compounds on insulin secretion [218];
- -
- GLUTag: enteroendocrine cells that allow the secretion of GLP-1 (intestinal hormone regulated by the DPP-IV enzyme) to be measured using the ELISA technique [219];
- -
- 3T3-L1: adipocyte cells that allow the measurement of glucose absorption by fluorimetry [220];
- -
- STC-1: intestinal secretin tumor cell line that expresses and secretes gut hormones in response to physiological stimuli [221].
6.2. In Vivo Analysis
6.2.1. Invertebrates Models
6.2.2. Vertebrates Models
6.2.3. Humans
7. Conclusions
Funding
Conflicts of Interest
References
- Nelson, D.L.; Cox, M.M. Lehninger: Principles of Biochemistry; W H Freeman & Co: New York, NY, USA, 2008; ISBN 9788578110796. [Google Scholar]
- Möller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef]
- McDonald, J.K. An overview of protease specificity and catalytic mechanisms: Aspects related to nomenclature and classification. Histochem. J. 1985, 17, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: London, UK, 1986; ISBN 0-85334-386-1. [Google Scholar]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef] [Green Version]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, S.; Parkin, K. Fennema’s Food Chemistry; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Genot, C.; Kabri, T.H.; Meynier, A. Stabilization of omega-3 oils and enriched foods using emulsifiers. In Food Enrichment with Omega-3 Fatty Acids; Woodhead Publishing: Cambridge, UK, 2013; pp. 151–193. [Google Scholar]
- Lam, R.S.H.; Nickerson, M.T. Food proteins: A review on their emulsifying properties using a structure–function approach. Food Chem. 2013, 141, 975–984. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Zayas, J.F. Water Holding Capacity of Proteins. In Functionality of Proteins in Food; Springer: Berlin/Heidelberg, Germany, 1997; pp. 76–133. [Google Scholar]
- Zielińska, E.; Karaś, M.; Baraniak, B. Comparison of functional properties of edible insects and protein preparations thereof. LWT 2018, 91, 168–174. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Aluko, R.E. Amino Acids, Peptides, and Proteins as Antioxidants for Food Preservation; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9781782420972. [Google Scholar]
- Padial-Domínguez, M.; Espejo-Carpio, F.J.; García-Moreno, P.J.; Jacobsen, C.; Guadix, E.M. Protein derived emulsifiers with antioxidant activity for stabilization of omega-3 emulsions. Food Chem. 2020, 329, 127148. [Google Scholar] [CrossRef]
- Morales-Medina, R.; Tamm, F.; Guadix, A.M.; Guadix, E.M.; Drusch, S. Functional and antioxidant properties of hydrolysates of sardine (S. pilchardus) and horse mackerel (T. mediterraneus) for the microencapsulation of fish oil by spray-drying. Food Chem. 2016, 194, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Liang, J.H. Effect of antioxidants on the oxidative stability of chicken breast meat in a dispersion system. J. Food Sci. 2002, 67, 530–533. [Google Scholar] [CrossRef]
- Hannu, K.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Acquah, C.; Di Stefano, E.; Udenigwe, C.C. Role of hydrophobicity in food peptide functionality and bioactivity. J. Food Bioact. 2018, 4, 88–98. [Google Scholar] [CrossRef] [Green Version]
- García-Moreno, P.J.; Pérez-Gálvez, R.; Espejo-Carpio, F.J.; Ruiz-Quesada, C.; Pérez-Morilla, A.I.; Martínez-Agustín, O.; Guadix, A.; Guadix, E.M. Functional, bioactive and antigenicity properties of blue whiting protein hydrolysates: Effect of enzymatic treatment and degree of hydrolysis. J. Sci. Food Agric. 2017, 97, 299–308. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; O’Cuinn, G. Enzymatic debittering of food protein hydrolysates. Biotechnol. Adv. 2006, 24, 234–237. [Google Scholar] [CrossRef]
- Li, R.; Zhang, P.; Barker, L.E.; Chowdhury, F.M.; Zhang, X. Cost-effectiveness of interventions to prevent and control diabetes mellitus: A systematic review. Diabetes Care 2010, 33, 1872–1894. [Google Scholar] [CrossRef] [Green Version]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef]
- Skeggs, L.T.; Kahn, H.R.; Shumway, N.P. The preparation and function of the hypertension-converting enzyme. J. Exp. Med. 1956, 103, 295–299. [Google Scholar] [CrossRef]
- Mizuno, S.; Matsuura, K.; Gotou, T.; Nishimura, S.; Kajimoto, O.; Yabune, M.; Kajimoto, Y.; Yamamoto, N. Antihypertensive effect of casein hydrolysate in a placebo-controlled study in subjects with high-normal blood pressure and mild hypertension. Br. J. Nutr. 2005, 94, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Hajieva, P.; Behl, C. Antioxidants as a potential therapy against age-related neurodegenerative diseases: Amyloid Beta toxicity and Alzheimer’s disease. Curr. Pharm. Des. 2006, 12, 699–704. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Masci, P.P.; Addepalli, R.; Chen, W.; Gobe, G.C.; Osborne, S.A. In vitro anti-thrombotic and anti-coagulant properties of blacklip abalone (Haliotis rubra) viscera hydrolysate. Anal. Bioanal. Chem. 2017, 409, 4195–4205. [Google Scholar] [CrossRef] [PubMed]
- Meram, C.; Wu, J. Anti-inflammatory effects of egg yolk livetins (α, β, and γ-livetin) fraction and its enzymatic hydrolysates in lipopolysaccharide-induced RAW 264.7 macrophages. Food Res. Int. 2017, 100, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Suwal, S.; Huang, J.-Y.; Liceaga, A.M. Selective separation and characterisation of dual ACE and DPP-IV inhibitory peptides from rainbow trout (Oncorhynchus mykiss) protein hydrolysates. Int. J. Food Sci. Technol. 2018, 1–12. [Google Scholar] [CrossRef]
- Nauck, M.A.; Baller, B.; Meier, J.J. Gastric inhibitory polypeptide and glucagon-like peptide-1 in the pathogenesis of type 2 diabetes. Diabetes 2004, 53. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhao, J.; Yang, R.; Zhao, W. Bioactive peptides with antidiabetic properties: A review. Int. J. Food Sci. Technol. 2019, 54, 1909–1919. [Google Scholar] [CrossRef] [Green Version]
- Tahrani, A.A.; Bailey, C.J.; Del Prato, S.; Barnett, A.H. Management of type 2 diabetes: New and future developments in treatment. Lancet 2011, 378, 182–197. [Google Scholar] [CrossRef]
- Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean-and lupin-derived peptides inhibit DPP-IV activity on in situ human intestinal Caco-2 cells and ex vivo human serum. Nutrients 2018, 10, 1082. [Google Scholar] [CrossRef] [Green Version]
- Caron, J.; Domenger, D.; Dhulster, P.; Ravallec, R.; Cudennec, B. Protein digestion-derived peptides and the peripheral regulation of food intake. Front. Endocrinol. (Lausanne) 2017, 8. [Google Scholar] [CrossRef]
- International Diabetes Federation. Diabetes Atlas de la FID; International Diabetes Federation: Brussels, Belgium, 2017; Volume 8, ISBN 2-930229-80-2. [Google Scholar]
- Li, L.; Hölscher, C. Common pathological processes in Alzheimer disease and type 2 diabetes: A review. Brain Res. Rev. 2007, 56, 384–402. [Google Scholar] [CrossRef]
- Howard-Thompson, A.; Khan, M.; Jones, M.; George, C.M. Type 2 Diabetes Mellitus: Outpatient Insulin Management. Am. Fam. Physician 2018, 97, 29–37. [Google Scholar] [PubMed]
- Sufian, M.K.N.B.; Hira, T.; Miyashita, K.; Nishi, T.; Asano, K.; Hara, H. Pork peptone stimulates cholecystokinin secretion from enteroendocrine cells and suppresses appetite in rats. Biosci. Biotechnol. Biochem. 2006, 70, 1869–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishi, T.; Hara, H.; Asano, K.; Tomita, F. The soybean β-conglycinin β 51-63 fragment suppresses appetite by stimulating cholecystokinin release in rats. Biochem. Mol. Actions Nutr. 2003, 133, 2537–2542. [Google Scholar] [CrossRef] [PubMed]
- Santos-Hernández, M.; Miralles, B.; Amigo, L.; Recio, I. Intestinal Signaling of Proteins and Digestion-Derived Products Relevant to Satiety. J. Agric. Food Chem. 2018, 66, 10123–10131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konrad, B.; Anna, D.; Marek, S.; Marta, P.; Aleksandra, Z.; Józefa, C. The evaluation of dipeptidyl peptidase (DPP)-IV, α-glucosidase and angiotensin converting enzyme (ACE) inhibitory activities of whey proteins hydrolyzed with serine protease isolated from asian pumpkin (Cucurbita ficifolia). Int. J. Pept. Res. Ther. 2014, 20, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2017, 91, 370–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kshirsagar, A.D.; Aggarwal, A.S.; Harle, U.N.; Deshpande, A.D. DPP IV inhibitors: Successes, failures and future prospects. Diabetes Metab. Syndr. Clin. Res. Rev. 2011, 5, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Marya; Khan, H.; Nabavi, S.M.; Habtemariam, S. Anti-diabetic potential of peptides: Future prospects as therapeutic agents. Life Sci. 2018, 193, 153–158. [Google Scholar] [CrossRef]
- Thoma, R.; Löffler, B.; Stihle, M.; Huber, W.; Ruf, A.; Hennig, M. Structural Basis of Proline-Specific Exopeptidase Activity as Observed in Human Dipeptidyl Peptidase-IV. Structure 2003, 11, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, A.T.; Feinle-Bisset, C.; Fitzgerald, P.C.E.; Standfield, S.; Horowitz, M.; Clifton, P.M.; Luscombe-Marsh, N.D. Comparative effects of intraduodenal whey protein hydrolysate on antropyloroduodenal motility, gut hormones, glycemia, appetite, and energy intake in lean and obese men. Am. J. Clin. Nutr. 2015, 102, 1323–1331. [Google Scholar] [CrossRef]
- Liu, R.; Cheng, J.; Wu, H. Discovery of Food-Derived Dipeptidyl Peptidase IV Inhibitory Peptides: A Review. Int. J. Mol. Sci. 2019, 20, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juillerat-Jeanneret, L. Dipeptidyl peptidase IV and its inhibitors: Therapeutics for type 2 diabetes and what else? J. Med. Chem. 2014, 57, 2197–2212. [Google Scholar] [CrossRef] [PubMed]
- Hsia, D.S.; Grove, O.; Cefalu, W.T. An Update on SGLT2 Inhibitors for the Treatment of Diabetes Mellitus. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Caron, J.; Cudennec, B.; Domenger, D.; Belguesmia, Y.; Flahaut, C.; Kouach, M.; Lesage, J.; Goossens, J.F.; Dhulster, P.; Ravallec, R. Simulated GI digestion of dietary protein: Release of new bioactive peptides involved in gut hormone secretion. Food Res. Int. 2016, 89, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Mata-Cases, M.; Rodríguez-Sánchez, B.; Mauricio, D.; Real, J.; Vlacho, B.; Franch-Nadal, J.; Oliva, J. The association between poor glycemic control and health care costs in people with diabetes: A population-based study. Diabetes Care 2020, 43, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Hewage, S.S.; Wu, S.; Neelakantan, N.; Yoong, J. Systematic review of effectiveness and cost-effectiveness of lifestyle interventions to improve clinical diabetes outcome measures in women with a history of GDM. Clin. Nutr. ESPEN 2020, 35, 20–29. [Google Scholar] [CrossRef]
- Daliri, E.; Oh, D.; Lee, B. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Pérez-Gálvez, R.; Morales-Medina, R.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Modelling of the production of ACE inhibitory hydrolysates of horse mackerel using proteases mixtures. Food Funct. 2016, 7, 3890–3901. [Google Scholar] [CrossRef]
- Espejo-Carpio, F.J.; Pérez-Gálvez, R.; Guadix, A.; Guadix, E.M. Artificial neuronal networks (ANN) to model the hydrolysis of goat milk protein by subtilisin and trypsin. J. Dairy Res. 2018, 85, 339–346. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Cadamuro, C.; Le Gouic, A.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Dipeptidyl peptidase IV (DPP-IV) inhibitory properties of a camel whey protein enriched hydrolysate preparation. Food Chem. 2019, 279, 70–79. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Fraguas, J.; Mirón, J.; Valcárcel, J.; Pérez-Martín, R.I.; Antelo, L.T. Valorisation of fish discards assisted by enzymatic hydrolysis and microbial bioconversion: Lab and pilot plant studies and preliminary sustainability evaluation. J. Clean. Prod. 2020, 246, 119027. [Google Scholar] [CrossRef]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; O’Keeffe, M.B.; Le Gouic, A.V.; Allsopp, P.J.; Mcsorley, E.M.; Sharkey, S.; Whooley, J.; McGovern, B.; O’Harte, F.P.M.; et al. Identification and characterisation of peptides from a boarfish (Capros aper) protein hydrolysate displaying in vitro dipeptidyl peptidase-IV (DPP-IV) inhibitory and insulinotropic activity. Food Res. Int. 2020, 131, 108989. [Google Scholar] [CrossRef] [PubMed]
- Wali, A.; Ma, H.; Shahnawaz, M.; Hayat, K.; Xiaong, J.; Jing, L. Impact of power ultrasound on antihypertensive activity, functional properties, and thermal stability of rapeseed protein hydrolysates. J. Chem. 2017, 2017. [Google Scholar] [CrossRef]
- Knezevic-Jugovic, Z.D.; Stefanović, A.B.; Žuža, M.G.; Milovanović, S.L.; Jakovetić, S.M.; Manojlović, V.B.; Bugarski, B.M. Effects of sonication and high-pressure carbon dioxide processing on enzymatic hydrolysis of egg white proteins. Acta Period. Technol. 2012, 43, 33–41. [Google Scholar] [CrossRef]
- Yu, H.-C.; Tan, F.-J. Effect of ultrasonic pretreatment on the properties of porcine liver protein hydrolysates. Int. J. Food Sci. Technol. 2017, 52, 1392–1399. [Google Scholar] [CrossRef]
- Perreault, V.; Hénaux, L.; Bazinet, L.; Doyen, A. Pretreatment of flaxseed protein isolate by high hydrostatic pressure: Impacts on protein structure, enzymatic hydrolysis and final hydrolysate antioxidant capacities. Food Chem. 2017, 221, 1805–1812. [Google Scholar] [CrossRef]
- Galazka, V.B.; Dickinson, E.; Ledward, D.A. Influence of high pressure processing on protein solutions and emulsions. Curr. Opin. Colloid Interface Sci. 2000, 5, 182–187. [Google Scholar] [CrossRef]
- Muntean, M.-V.; Marian, O.; Barbieru, V.; Cătunescu, G.M.; Ranta, O.; Drocas, I.; Terhes, S. High Pressure Processing in Food Industry–Characteristics and Applications. Agric. Agric. Sci. Procedia 2016, 10, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Marciniak, A.; Suwal, S.; Naderi, N.; Pouliot, Y.; Doyen, A. Enhancing enzymatic hydrolysis of food proteins and production of bioactive peptides using high hydrostatic pressure technology. Trends Food Sci. Technol. 2018, 80, 187–198. [Google Scholar] [CrossRef]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef]
- Tavano, O.L. Protein hydrolysis using proteases: An important tool for food biotechnology. J. Mol. Catal. B Enzym. 2013, 90, 1–11. [Google Scholar] [CrossRef]
- Tavano, O.L.; Berenguer-Murcia, A.; Secundo, F.; Fernandez-Lafuente, R. Biotechnological Applications of Proteases in Food Technology. Compr. Rev. Food Sci. Food Saf. 2018, 17, 412–436. [Google Scholar] [CrossRef] [Green Version]
- Adamson, N.J.; Reynolds, E.C. Characterization of casein phosphopeptides prepared using alcalase: Determination of enzyme specificity. Enzym. Microb. Technol. 1996, 19, 202–207. [Google Scholar] [CrossRef]
- Olsen, J.V.; Ong, S.-E.; Mann, M. Trypsin Cleaves Exclusively C-terminal to Arginine and Lysine Residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura Campos, M.R.; Chel Guerrero, L.A.; Betancur Ancona, D.A. Angiotensin-I converting enzyme inhibitory and antioxidant activities of peptide fractions extracted by ultrafiltration of cowpea Vigna unguiculata hydrolysates. J. Sci. Food Agric. 2010, 90, 2512–2518. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Atlantic salmon (Salmo salar) co-product-derived protein hydrolysates: A source of antidiabetic peptides. Food Res. Int. 2018, 106, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Rivero-Pino, F.; Pérez Gálvez, A.R.; Espejo-Carpio, F.J.; Guadix, E.M. Evaluation of Tenebrio molitor protein as source of peptides modulating physiological processes. Food Funct. 2020, 11, 4376–4386. [Google Scholar] [CrossRef]
- Connolly, A.; Piggott, C.O.; FitzGerald, R.J. In vitro α-glucosidase, angiotensin converting enzyme and dipeptidyl peptidase-IV inhibitory properties of brewers’ spent grain protein hydrolysates. Food Res. Int. 2014, 56, 100–107. [Google Scholar] [CrossRef]
- Arise, R.O.; Idi, J.J.; Mic-Braimoh, I.M.; Korode, E.; Ahmed, R.N.; Osemwegie, O. In vitro Angiotesin-1-converting enzyme, α-amylase and α-glucosidase inhibitory and antioxidant activities of Luffa cylindrical (L.)M. Roem seed protein hydrolysate. Heliyon 2019, 5, e01634. [Google Scholar] [CrossRef] [Green Version]
- Horner, K.; Drummond, E.; Brennan, L. Bioavailability of milk protein-derived bioactive peptides: A glycaemic management perspective. Nutr. Res. Rev. 2016, 29, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Miralles, B.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Amigo, L.; Recio, I. Health-related functional value of dairy proteins and peptides. In Proteins in Food Processing; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; pp. 523–568. ISBN 9780081007297. [Google Scholar]
- Montesano, D.; Gallo, M.; Blasi, F.; Cossignani, L. Biopeptides from vegetable proteins: New scientific evidences. Curr. Opin. Food Sci. 2020, 31, 31–37. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Dipeptidyl peptidase-IV inhibitory activity of dairy protein hydrolysates. Int. Dairy J. 2012, 25, 97–102. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Blue whiting (Micromesistius poutassou) muscle protein hydrolysate with in vitro and in vivo antidiabetic properties. J. Funct. Foods 2018, 40, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.L.; Jao, C.L.; Ho, K.P.; Hsu, K.C. Dipeptidyl-peptidase IV inhibitory activity of peptides derived from tuna cooking juice hydrolysates. Peptides 2012, 35, 114–121. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Bioactive peptides from Atlantic salmon (Salmo salar) with angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory, and antioxidant activities. Food Chem. 2017, 218, 396–405. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Production and identification of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from discarded Sardine pilchardus protein. Food Chem. 2020, 328, 127096. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M. Global diets link environmental sustainability and human health. Nature 2014, 515, 518–522. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Unlocking the biological potential of proteins from edible insects through enzymatic hydrolysis: A review. Innov. Food Sci. Emerg. Technol. 2017, 43, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Bioactive Peptides Derived from Seaweed Protein and Their Health Benefits: Antihypertensive, Antioxidant, and Antidiabetic Properties. J. Food Sci. 2018, 83, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Harnedy, P.A.; FitzGerald, R.J. In vitro assessment of the cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Escuredo, O.; González Martín, M.I.; Wells Moncada, G.; Fischer, S.; Hernández Hierro, J.M. Amino acid profile of the quinoa (Chenopodium quinoa Willd.) using near infrared spectroscopy and chemometric techniques. J. Cereal Sci. 2014, 60, 67–74. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Boye, J.I. Comparative Study of the Effects of Processing on the Nutritional, Physicochemical and Functional Properties of Lentil. J. Food Process. Preserv. 2017, 41, 24–30. [Google Scholar] [CrossRef]
- Connolly, A.; Piggott, C.O.; FitzGerald, R.J. Characterisation of protein-rich isolates and antioxidative phenolic extracts from pale and black brewers’ spent grain. Int. J. Food Sci. Technol. 2013, 48, 1670–1681. [Google Scholar] [CrossRef]
- Azagoh, C.; Ducept, F.; Garcia, R.; Rakotozafy, L.; Cuvelier, M.-E.; Keller, S.; Lewandowski, R.; Mezdour, S. Extraction and physicochemical characterization of Tenebrio molitor proteins. Food Res. Int. 2016, 88, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Breternitz, N.R.; Bolini, H.M.A.; Hubinger, M.D. Sensory acceptance evaluation of a new food flavoring produced by microencapsulation of a mussel (Perna perna) protein hydrolysate. LWT-Food Sci. Technol. 2017, 83, 141–149. [Google Scholar] [CrossRef]
- Sathivel, S.; Bechtel, P.J.; Babbitt, J.; Smiley, S.; Crapo, C.; Reppond, K.D.; Prinyawiwatkul, W. Biochemical and Functional Properties of Herring (Clupea harengus) Byproduct Hydrolysates. J. Food Sci. 2003, 68, 2196–2200. [Google Scholar] [CrossRef]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef]
- Gonzalez-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martinez-Villaluenga, C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, a-amylase, and a-glucosidase enzymes. Int. J. Mol. Sci. 2018, 19, 2883. [Google Scholar] [CrossRef] [Green Version]
- Parthsarathy, V.; McLaughlin, C.M.; Harnedy, P.A.; Allsopp, P.J.; Crowe, W.; McSorley, E.M.; FitzGerald, R.J.; O’Harte, F.P.M.M. Boarfish (Capros aper) protein hydrolysate has potent insulinotropic and GLP-1 secretory activity in vitro and acute glucose lowering effects in mice. Int. J. Food Sci. Technol. 2018, 54, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Lamoureux, C.; Fitzgerald, R.J. Generation of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides during the enzymatic hydrolysis of tropical banded cricket (Gryllodes sigillatus) proteins. Food Funct. 2018, 9, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammi, C.; Bollati, C.; Gelain, F.; Arnoldi, A.; Pugliese, R. Enhancement of the stability and anti-DPPIV activity of hempseed hydrolysates through self-assembling peptide-based hydrogels. Front. Chem. 2019, 7, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.-L.; Hussain, N.; Joy Ujiroghene, O.; Pang, X.-Y.; Zhang, S.-W.; Lu, J.; Liu, L.; Lv, J.-P. Generation and characterization of dipeptidyl peptidase-IV inhibitory peptides from trypsin-hydrolyzed α-lactalbumin-rich whey proteins. Food Chem. 2020, 318, 126333. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.X.; Wang, B.K.; Wu, Y.C.; Li, Q.Y.; Qin, B.W.; Li, H.J. Release of antidiabetic peptides from Stichopus japonicas by simulated gastrointestinal digestion. Food Chem. 2020, 315, 126273. [Google Scholar] [CrossRef] [PubMed]
- Megrous, S.; Al-Dalali, S.; Zhao, X.; Chen, C.; Cao, Y.; Bourouis, I.; Mekkaoui, A.; Yang, Z.; Yang, Z. Evaluation of Antidiabetic Activities of Casein Hydrolysates by a Bacillus Metalloendopeptidase. Int. J. Pept. Res. Ther. 2020, 1–9. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, L.; Liu, C.; Liu, X.; Li, H.; Shi, J.; Li, M.; Min, W. Anti-diabetic effect by walnut (Juglans mandshurica Maxim.)-derived peptide LPLLR through inhibiting α-glucosidase and α-amylase, and alleviating insulin resistance of hepatic HepG2 cells. J. Funct. Foods 2020, 69, 103944. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Baraniak, B.; Jakubczyk, A. Evaluation of ACE, α-glucosidase, and lipase inhibitory activities of peptides obtained by in vitro digestion of selected species of edible insects. Eur. Food Res. Technol. 2020, 246, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Karimi, A.; Azizi, M.H.; Ahmadi Gavlighi, H. Frationation of hydrolysate from corn germ protein by ultrafiltration: In vitro antidiabetic and antioxidant activity. Food Sci. Nutr. 2020, 8, 2395–2405. [Google Scholar] [CrossRef] [Green Version]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Jęderka, K.; Lewicki, S.; Złotek, U. Different temperature treatments of millet grains affect the biological activity of protein hydrolyzates and peptide fractions. Nutrients 2019, 11, 550. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Pérez, E.; Jiménez-Morales, K.; Quintal-Novelo, C.; Moo-Puc, R.; Chel-Guerrero, L.; Betancur-Ancona, D. Enzymatic protein hydrolysates and ultrafiltered peptide fractions from Cowpea Vigna unguiculata L bean with in vitro antidiabetic potential. J. Iran. Chem. Soc. 2019, 16, 1773–1781. [Google Scholar] [CrossRef]
- Valencia-Mejía, E.; Batista, K.A.; Fernández, J.J.A.; Fernandes, K.F. Antihyperglycemic and hypoglycemic activity of naturally occurring peptides and protein hydrolysates from easy-to-cook and hard-to-cook beans (Phaseolus vulgaris L.). Food Res. Int. 2019, 121, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Karthiraj, T.; Harish Babu, B.; Senthil Kumar, R. Task-specific deep eutectic solvent based extraction coupled cascade chromatography quantification of α-glucosidase inhibitory peptide from Ocimum tenuriflorum seeds. Microchem. J. 2020, 157, 104883. [Google Scholar] [CrossRef]
- Cermeño, M.; Connolly, A.; O’Keeffe, M.B.; Flynn, C.; Alashi, A.M.; Aluko, R.E.; FitzGerald, R.J. Identification of bioactive peptides from brewers’ spent grain and contribution of Leu/Ile to bioactive potency. J. Funct. Foods 2019, 60, 103455. [Google Scholar] [CrossRef]
- Cermeño, M.; Stack, J.; Tobin, P.R.; O’Keeffe, M.B.; Harnedy, P.A.; Stengel, D.B.; Fitzgerald, R.J. Peptide identification from a: Porphyra dioica protein hydrolysate with antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activities. Food Funct. 2019, 10, 3421–3429. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Identification of Bioactive Peptides with α-Amylase Inhibitory Potential from Enzymatic Protein Hydrolysates of Red Seaweed (Porphyra spp.). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef]
- Jiang, M.; Yan, H.; He, R.; Ma, Y. Purification and a molecular docking study of α-glucosidase-inhibitory peptides from a soybean protein hydrolysate with ultrasonic pretreatment. Eur. Food Res. Technol. 2018, 244, 1995–2005. [Google Scholar] [CrossRef]
- Ina, S.; Hamada, A.; Nakamura, H.; Yamaguchi, Y.; Kumagai, H.; Kumagai, H. Rice (Oryza sativa japonica) albumin hydrolysates suppress postprandial blood glucose elevation by adsorbing glucose and inhibiting Na+-d-glucose cotransporter SGLT1 expression. J. Funct. Foods 2020, 64, 103603. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R.M. Tuber Storage Proteins as Potential Precursors of Bioactive Peptides: An In Silico Analysis. Int. J. Pept. Res. Ther. 2019, 25, 437–446. [Google Scholar] [CrossRef]
- Evaristus, N.A.; Wan Abdullah, W.N.; Gan, C.Y. Extraction and identification of α-amylase inhibitor peptides from Nephelium lappacheum and Nephelium mutabile seed protein using gastro-digestive enzymes. Peptides 2018, 102, 61–67. [Google Scholar] [CrossRef]
- Ngoh, Y.Y.; Gan, C.Y. Identification of Pinto bean peptides with inhibitory effects on α-amylase and angiotensin converting enzyme (ACE) activities using an integrated bioinformatics-assisted approach. Food Chem. 2018, 267, 124–131. [Google Scholar] [CrossRef]
- Mohd Salim, M.A.S.; Gan, C.Y. Dual-function peptides derived from egg white ovalbumin: Bioinformatics identification with validation using in vitro assay. J. Funct. Foods 2020, 64, 103618. [Google Scholar] [CrossRef]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of novel DPP–IV inhibitory peptides from Atlantic salmon (Salmo salar) skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Richel, A.; Hao, Y.; Fan, X.; Everaert, N.; Yang, X.; Ren, G. Novel dipeptidyl peptidase-IV and angiotensin-I-converting enzyme inhibitory peptides released from quinoa protein by in silico proteolysis. Food Sci. Nutr. 2020, 8, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Aiello, G.; Bollati, C.; Bartolomei, M.; Arnoldi, A.; Lammi, C. Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity. Nutrients 2020, 12, 794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartal, C.; Kaplan Türköz, B.; Otles, S. Prediction, identification and evaluation of bioactive peptides from tomato seed proteins using in silico approach. J. Food Meas. Charact. 2020, 1–19. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, D.; Yu, Z.; Ding, L.; Liu, J. Novel membrane peptidase inhibitory peptides with activity against angiotensin converting enzyme and dipeptidyl peptidase IV identified from hen eggs. J. Funct. Foods 2020, 64, 103649. [Google Scholar] [CrossRef]
- Zhang, C.; Gao, S.; Zhang, C.; Zhang, Y.; Liu, H.; Hong, H.; Luo, Y. Evaluating in vitro dipeptidyl peptidase IV inhibition by peptides from common carp (Cyprinus carpio) roe in cell culture models. Eur. Food Res. Technol. 2020, 246, 179–191. [Google Scholar] [CrossRef]
- Aiello, G.; Li, Y.; Boschin, G.; Bollati, C.; Arnoldi, A.; Lammi, C. Chemical and biological characterization of spirulina protein hydrolysates: Focus on ACE and DPP-IV activities modulation. J. Funct. Foods 2019, 63, 103592. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, R.; Fang, L.; Qin, X.; Cai, M.; Gu, R.; Lu, J.; Wang, Y. Hypoglycemic effects and biochemical mechanisms of Pea oligopeptide on high-fat diet and streptozotocin-induced diabetic mice. J. Food Biochem. 2019, 43. [Google Scholar] [CrossRef]
- Ashok, A.; Brijesha, N.; Aparna, H.S. Discovery, synthesis, and in vitro evaluation of a novel bioactive peptide for ACE and DPP-IV inhibitory activity. Eur. J. Med. Chem. 2019, 180, 99–110. [Google Scholar] [CrossRef]
- Casanova-Martí, À.; Bravo, F.I.; Serrano, J.; Ardévol, A.; Pinent, M.; Muguerza, B. Antihyperglycemic effect of a chicken feet hydrolysate: Via the incretin system: DPP-IV-inhibitory activity and GLP-1 release stimulation. Food Funct. 2019, 10, 4062–4070. [Google Scholar] [CrossRef] [PubMed]
- Gomez, H.L.R.; Peralta, J.P.; Tejano, L.A.; Chang, Y.-W. In Silico and In Vitro Assessment of Portuguese Oyster (Crassostrea angulata) Proteins as Precursor of Bioactive Peptides. Int. J. Mol. Sci. 2019, 20, 5191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Xu, Q.; Lin, L.; Zeng, X.A.; Sun, B.; Zhao, M. In Vitro Metabolic Stability of a Casein-Derived Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Peptide VPYPQ and Its Controlled Release from Casein by Enzymatic Hydrolysis. J. Agric. Food Chem. 2019, 67, 10604–10613. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, J.; O’Sullivan, M.; Casey, E. Production of Whey-Derived DPP-IV Inhibitory Peptides Using an Enzymatic Membrane Reactor. Food Bioprocess Technol. 2019, 12, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Du, Z.; Yu, Z.; Liu, J. Hydrolysis and Transport of Egg White-Derived Peptides in Caco-2 Cell Monolayers and Everted Rat Sacs. J. Agric. Food Chem. 2019, 67, 4839–4848. [Google Scholar] [CrossRef]
- Xu, F.; Yao, Y.; Xu, X.; Wang, M.; Pan, M.; Ji, S.; Wu, J.; Jiang, D.; Ju, X.; Wang, L. Identification and Quantification of DPP-IV-Inhibitory Peptides from Hydrolyzed-Rapeseed-Protein-Derived Napin with Analysis of the Interactions between Key Residues and Protein Domains. J. Agric. Food Chem. 2019, 67, 3679–3690. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Dávalos Terán, I.; Fogliano, V.; Wichers, H.J. Investigation into the potential of commercially available lesser mealworm (A. diaperinus) protein to serve as sources of peptides with DPP-IV inhibitory activity. Int. J. Food Sci. Technol. 2019, 54, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Mudgil, P.; Jobe, B.; Kamal, H.; Alameri, M.; Al Ahbabi, N.; Maqsood, S. Dipeptidyl peptidase-IV, α-amylase, and angiotensin I converting enzyme inhibitory properties of novel camel skin gelatin hydrolysates. LWT 2019, 101, 251–258. [Google Scholar] [CrossRef]
- Lima, R.D.C.L.; Berg, R.S.; Rønning, S.B.; Afseth, N.K.; Knutsen, S.H.; Staerk, D.; Wubshet, S.G. Peptides from chicken processing by-product inhibit DPP-IV and promote cellular glucose uptake: Potential ingredients for T2D management. Food Funct. 2019, 10, 1619–1628. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Miralles, B.; Hernández-Ledesma, B. Release of multifunctional peptides from kiwicha (Amaranthus caudatus) protein under in vitro gastrointestinal digestion. J. Sci. Food Agric. 2019, 99, 1225–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, H.; Hong, H.; Luo, Y. Purification and identification of dipeptidyl peptidase IV and angiotensin-converting enzyme inhibitory peptides from silver carp (Hypophthalmichthys molitrix) muscle hydrolysate. Eur. Food Res. Technol. 2019, 245, 243–255. [Google Scholar] [CrossRef]
- Han, R.; Maycock, J.; Murray, B.S.; Boesch, C. Identification of angiotensin converting enzyme and dipeptidyl peptidase-IV inhibitory peptides derived from oilseed proteins using two integrated bioinformatic approaches. Food Res. Int. 2019, 115, 283–291. [Google Scholar] [CrossRef]
- Mune Mune, M.A.; Minka, S.R.; Henle, T. Investigation on antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activity of Bambara bean protein hydrolysates. Food Chem. 2018, 250, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Dávalos Terán, I.; Imai, K.; Lacroix, I.M.E.; Fogliano, V.; Udenigwe, C.C. Bioinformatics of edible yellow mealworm (Tenebrio molitor) proteome reveal the cuticular proteins as promising precursors of dipeptidyl peptidase-IV inhibitors. J. Food Biochem. 2019, 44, e13121. [Google Scholar] [CrossRef] [PubMed]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Drummond, E.; Flynn, S.; Whelan, H.; Nongonierma, A.B.; Holton, T.A.; Robinson, A.; Egan, T.; Cagney, G.; Shields, D.C.; Gibney, E.R.; et al. Casein Hydrolysate with Glycemic Control Properties: Evidence from Cells, Animal Models, and Humans. J. Agric. Food Chem. 2018, 66, 4352–4363. [Google Scholar] [CrossRef]
- Sila, A.; Alvarez, O.M.; Haddar, A.; Frikha, F.; Dhulster, P.; Nedjar-Arroume, N.; Bougatef, A. Purification, identification and structural modelling of DPP-IV inhibiting peptides from barbel protein hydrolysate. J. Chromatogr. B 2016, 1008, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Paolella, S.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Identification of novel dipeptidyl peptidase IV (DPP-IV) inhibitory peptides in camel milk protein hydrolysates. Food Chem. 2018, 244, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Food-derived dipeptidyl-peptidase IV inhibitors as a potential approach for glycemic regulation–Current knowledge and future research considerations. Trends Food Sci. Technol. 2016, 54, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Oseguera-Toledo, M.E.; Gonzalez de Mejia, E.; Amaya-Llano, S.L. Hard-to-cook bean (Phaseolus vulgaris L.) proteins hydrolyzed by alcalase and bromelain produced bioactive peptide fractions that inhibit targets of type-2 diabetes and oxidative stress. Food Res. Int. 2015, 76, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Inhibition of dipeptidyl peptidase (DPP)-IV and α-glucosidase activities by pepsin-treated whey proteins. J. Agric. Food Chem. 2013, 61, 7500–7506. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Matsui, T.; Oki, T.; Osajima, Y. Isolation and identification of peptidic α-glucosidase inhibitors derived from sardine muscle hydrolyzate. Zeitschrift fur Naturforsch.-Sect. C J. Biosci. 1999, 54, 259–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Yin, Y.; Zhao, W.; Liu, J.; Chen, F. Anti-diabetic activity peptides from albumin against α-glucosidase and α-amylase. Food Chem. 2012, 135, 2078–2085. [Google Scholar] [CrossRef] [PubMed]
- Akkarachiyasit, S.; Yibchok-Anun, S.; Wacharasindhu, S.; Adisakwattana, S. In vitro inhibitory effects of cyandin-3-rutinoside on pancreatic α-amylase and its combined effect with acarbose. Molecules 2011, 16, 2075–2083. [Google Scholar] [CrossRef]
- Espejo-Carpio, F.J.; Pérez-Gálvez, R.; Del Carmen Almécija, M.; Guadix, A.; Guadix, E.M. Production of goat milk protein hydrolysate enriched in ACE-inhibitory peptides by ultrafiltration. J. Dairy Res. 2014, 81, 385–393. [Google Scholar] [CrossRef]
- Ngoh, Y.-Y.; Gan, C.-Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef]
- Liu, R.; Zheng, W.; Li, J.; Wang, L.; Wu, H.; Wang, X.; Shi, L. Rapid identification of bioactive peptides with antioxidant activity from the enzymatic hydrolysate of Mactra veneriformis by UHPLC–Q-TOF mass spectrometry. Food Chem. 2015, 167, 484–489. [Google Scholar] [CrossRef]
- Perutka, Z.; Šebela, M. Basis of Mass Spectrometry: Technical Variants. In The Use of Mass Spectrometry Technology (MALDI-TOF) in Clinical Microbiology; Academic Press: Cambridge, MA, USA, 2018; pp. 19–45. ISBN 9780128144510. [Google Scholar]
- Ngoh, Y.-Y.; Tye, G.J.; Gan, C.-Y. The investigation of α-amylase inhibitory activity of selected Pinto bean peptides via preclinical study using AR42J cell. J. Funct. Foods 2017, 35, 641–647. [Google Scholar] [CrossRef]
- Siow, H.-L.; Gan, C.-Y. Extraction, identification, and structure–activity relationship of antioxidative and α-amylase inhibitory peptides from cumin seeds (Cuminum cyminum). J. Funct. Foods 2016, 22, 1–12. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Fitzgerald, R.J. An in silico model to predict the potential of dietary proteins as sources of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.-H.; Wang, T.-Y.; Hung, C.-C.; Jao, C.-L.; Hsieh, Y.-L.; Wu, S.-X.; Hsu, K.-C. In silico, in vitro and in vivo analyses of dipeptidyl peptidase IV inhibitory activity and the antidiabetic effect of sodium caseinate hydrolysate. Food Funct. 2016, 7, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- De Cicco, M.; Mamone, G.; Di Stasio, L.; Ferranti, P.; Addeo, F.; Picariello, G. Hidden “digestome”: Current analytical approaches provide incomplete peptide inventories of food digests. J. Agric. Food Chem. 2019, 67, 7775–7782. [Google Scholar] [CrossRef]
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. TrAC-Trends Anal. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- De Vries, S.J.; Rey, J.; Schindler, C.E.M.; Zacharias, M.; Tuffery, P. The pepATTRACT web server for blind, large-scale peptide-protein docking. Nucleic Acids Res. 2017, 45, W361–W364. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Fitzgerald, R.J. Learnings from quantitative structure-activity relationship (QSAR) studies with respect to food protein-derived bioactive peptides: A review. RSC Adv. 2016, 6, 75400–75413. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; Cermeño, M.; Khalesi, M.; Kleekayai, T.; Amigo-Benavent, M. Application of in silico approaches for the generation of milk protein-derived bioactive peptides. J. Funct. Foods 2020, 64, 103636. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J. Structure–activity relationships study on biological activity of peptides as dipeptidyl peptidase IV inhibitors by chemometric modeling. Chem. Biol. Drug Des. 2020, 95, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R.M. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, L.; González-Rogel, D.; Heres, A.; Toldrá, F. Iberian dry-cured ham as a potential source of α-glucosidase-inhibitory peptides. J. Funct. Foods 2020, 67, 103840. [Google Scholar] [CrossRef]
- Karaś, M. Influence of physiological and chemical factors on the absorption of bioactive peptides. Int. J. Food Sci. Technol. 2019, 54, 1486–1496. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; Fitzgerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Segura-Campos, M.; Chel-Guerrero, L.; Betancur-Ancona, D.; Hernandez-Escalante, V.M. Bioavailability of bioactive peptides. Food Rev. Int. 2011, 27, 213–226. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Xie, N.; Li, B. Influence of peptide characteristics on their stability, intestinal transport, and in vitro bioavailability: A review. J. Food Biochem. 2019, 43, e12571. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.L.; Mindell, J.A.; Newstead, S. Thermodynamic evidence for a dual transport mechanism in a POT peptide transporter. eLife 2014, 3, e04273. [Google Scholar] [CrossRef]
- Salamat-Miller, N.; Johnston, T.P. Current strategies used to enhance the paracellular transport of therapeutic polypeptides across the intestinal epithelium. Int. J. Pharm. 2005, 294, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Caron, J.; Domenger, D.; Dhulster, P.; Ravallec, R.; Cudennec, B. Using Caco-2 cells as novel identification tool for food-derived DPP-IV inhibitors. Food Res. Int. 2017, 92, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Erickson, R.H.; Matsumoto, H.; And, E.G.; Kim, Y.S. Expression of dipeptidyl aminopeptidase IV during enterocytic differentiation of human colon cancer (Caco-2) cells. Int. J. Cancer 1991, 47, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, H.; Chen, S.; Luo, Y. Evaluating the effects of IADHFL on inhibiting DPP-IV activity and expression in Caco-2 cells and contributing to the amount of insulin released from INS-1 cells: In vitro. Food Funct. 2018, 9, 2240–2250. [Google Scholar] [CrossRef]
- Gianfranceschi, G.L.; Gianfranceschi, G.; Quassinti, L.; Bramucci, M. Biochemical requirements of bioactive peptides for nutraceutical efficacy. J. Funct. Foods 2018, 47, 252–263. [Google Scholar] [CrossRef]
- Thanou, M.; Verhoef, J.; Junginger, H. Oral drug absorption enhancement by chitosan and its derivatives. Adv. Drug Deliv. Rev. 2001, 52, 117–126. [Google Scholar] [CrossRef]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; Crowe, W.; Allsopp, P.J.; McSorley, E.M.; Devaney, M.; Whooley, J.; McGovern, B.; Parthsarathy, V.; O’Harte, F.P.M.; et al. Stability to thermal treatment of dipeptidyl peptidase IV (DPP-IV) inhibitory activity of a boarfish (Capros aper) protein hydrolysate when incorporated into tomato-based products. Int. J. Food Sci. Technol. 2020, 1–8. [Google Scholar] [CrossRef]
- Sun, X.; Acquah, C.; Aluko, R.E.; Udenigwe, C.C. Considering food matrix and gastrointestinal effects in enhancing bioactive peptide absorption and bioavailability. J. Funct. Foods 2020, 64, 103680. [Google Scholar] [CrossRef]
- Mouécoucou, J.; Frémont, S.; Sanchez, C.; Villaume, C.; Méjean, L. In vitro allergenicity of peanut after hydrolysis in the presence of polysaccharides. Clin. Exp. Allergy 2004, 34, 1429–1437. [Google Scholar] [CrossRef]
- Mandalari, G.; Rigby, N.M.; Bisignano, C.; Lo Curto, R.B.; Mulholland, F.; Su, M.; Venkatachalam, M.; Robotham, J.M.; Willison, L.N.; Lapsley, K.; et al. Effect of food matrix and processing on release of almond protein during simulated digestion. LWT-Food Sci. Technol. 2014, 59, 439–447. [Google Scholar] [CrossRef]
- Seiquer, I.; Díaz-Alguacil, J.; Delgado-Andrade, C.; López-Frías, M.; Muñoz-Hoyos, A.; Galdó, G.; Navarro, M.P. Diets rich in Maillard reaction products affect protein digestibility in adolescent males aged 11–14 y. Am. J. Clin. Nutr. 2006, 83, 1082–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar-Villanea, S.; Butré, C.I.; Wierenga, P.A.; Bruininx, E.M.A.M.; Gruppen, H.; Hendriks, W.H.; van der Poel, A.F.B. Apparent ileal digestibility of Maillard reaction products in growing pigs. PLoS ONE 2018, 13, e0199499. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Parmar, P.; Tiwari, B.K. Stability of Proteins During Processing and Storage. In Proteins: Sustainable Source, Processing and Applications; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 295–330. ISBN 9780128166956. [Google Scholar]
- Van Lancker, F.; Adams, A.; De Kimpe, N. Chemical modifications of peptides and their impact on food properties. Chem. Rev. 2011, 111, 7876–7903. [Google Scholar] [CrossRef]
- Ten Have, G.A.M.; Van Der Pijl, P.C.; Kies, A.K.; Deutz, N.E.P.P. Enhanced lacto-tri-peptide bio-availability by co-ingestion of macronutrients. PLoS ONE 2015, 10, e0130638. [Google Scholar] [CrossRef] [PubMed]
- Graves, A.M.; Hettiarachchy, N.; Rayaprolu, S.; Li, R.; Horax, R.; Seo, H.S. Bioactivity of a Rice Bran-Derived Peptide and Its Sensory Evaluation and Storage Stability in Orange Juice. J. Food Sci. 2016, 81, H1010–H1015. [Google Scholar] [CrossRef]
- Contreras, M.; del, M.; Sevilla, M.A.; Monroy-Ruiz, J.; Amigo, L.; Gómez-Sala, B.; Molina, E.; Ramos, M.; Recio, I. Food-grade production of an antihypertensive casein hydrolysate and resistance of active peptides to drying and storage. Int. Dairy J. 2011, 21, 470–476. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Bioactive fish hydrolysates resistance to food processing. LWT-Food Sci. Technol. 2020, 117. [Google Scholar] [CrossRef]
- Li, X.; Farid, M. A review on recent development in non-conventional food sterilization technologies. J. Food Eng. 2016, 182, 33–45. [Google Scholar] [CrossRef]
- Chizoba Ekezie, F.-G.; Cheng, J.-H.; Sun, D.-W. Effects of nonthermal food processing technologies on food allergens: A review of recent research advances. Trends Food Sci. Technol. 2018, 74, 12–25. [Google Scholar] [CrossRef]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of processing on conformational changes of food proteins related to allergenicity. Trends Food Sci. Technol. 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto-Leppälä, A.; Rantamäki, P.; Tupasela, T. Impact of processing on bioactive proteins and peptides. Trends Food Sci. Technol. 1998, 9, 307–319. [Google Scholar] [CrossRef]
- Ustunol, Z. Physical, chemical, and processing-induced changes in proteins. In Applied Food Protein Chemistry; Willey Blackwell: Hoboken, NJ, USA, 2018; pp. 24–46. [Google Scholar]
- García-Moreno, P.J.; Batista, I.; Pires, C.; Bandarra, N.M.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Antioxidant activity of protein hydrolysates obtained from discarded Mediterranean fish species. Food Res. Int. 2014, 65, 469–476. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Agyei, D.; Udenigwe, C.C. Impact of processing on the chemistry and functionality of food proteins. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 27–45. ISBN 9780081007228. [Google Scholar]
- Ratti, C. Hot air and freeze-drying of high-value foods: A review. J. Food Eng. 2001, 49, 311–319. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, Y.; Soladoye, O.P.; Aluko, R.E. Maillard reaction products derived from food protein-derived peptides: Insights into flavor and bioactivity. Crit. Rev. Food Sci. Nutr. 2019, 8398. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.J.; Jiménez-Pérez, S. Free radical scavenging capacity of Maillard reaction products as related to colour and fluorescence. Food Chem. 2001, 72, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wu, C.; Fan, G.; Li, T.; Sun, Y. Improvement of antioxidant activity of Morchella esculenta protein hydrolysate by optimized glycosylation reaction. CYTA-J. Food 2018, 16, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Salam, M.H.; El-Shibiny, S. Glycation of whey proteins: Technological and nutritional implications. Int. J. Biol. Macromol. 2018, 112, 83–92. [Google Scholar] [CrossRef]
- Hong, X.; Meng, J.; Lu, R.R. Improvement of ACE inhibitory activity of casein hydrolysate by Maillard reaction with xylose. J. Sci. Food Agric. 2015, 95, 66–71. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Q.; Wang, S.; Qiu, C.; Liu, Y.; Su, G.; Zhao, M. Effects of Maillard reaction on bioactivities promotion of anchovy protein hydrolysate: The key role of MRPs and newly formed peptides with basic and aromatic amino acids. LWT-Food Sci. Technol. 2018, 97, 245–253. [Google Scholar] [CrossRef]
- Mohan, A.; Rajendran, S.R.C.K.; He, Q.S.; Bazinet, L.; Udenigwe, C.C. Encapsulation of food protein hydrolysates and peptides: A review. RSC Adv. 2015, 5, 79270–79278. [Google Scholar] [CrossRef]
- Wang, K.; Sun, D.W.; Pu, H.; Wei, Q. Principles and applications of spectroscopic techniques for evaluating food protein conformational changes: A review. Trends Food Sci. Technol. 2017, 67, 207–219. [Google Scholar] [CrossRef]
- Capuano, E.; Oliviero, T.; van Boekel, M.A.J. Modelling food matrix effects on chemical reactivity: Challenges and perspectives. Crit. Rev. Food Sci. Nutr. 2017, 58, 2814–2828. [Google Scholar] [CrossRef] [PubMed]
- Guyomarc’h, F.; Warin, F.; Donald Muir, D.; Leaver, J. Lactosylation of milk proteins during the manufacture and storage of skim milk powders. Int. Dairy J. 2000, 10, 863–872. [Google Scholar] [CrossRef]
- Albalá-Hurtado, S.; Veciana-Nogués, M.T.; Mariné-Font, A.; Vidal-Carou, M.C. Progress of browning reactions during storage of liquid infant milks. J. Agric. Food Chem. 1999, 47, 4033–4037. [Google Scholar] [CrossRef]
- McClenaghan, N.H.; Barnett, C.R.; Ah-Sing, E.; Abdel-Wahab, Y.H.A.; O’Harte, F.P.M.; Yoon, T.W.; Swanston-Flatt, S.K.; Flatt, P.R. Characterization of a novel glucose-responsive insulin-secreting cell line, BRIN-BD11, produced by electrofusion. Diabetes 1996, 45, 1132–1140. [Google Scholar] [CrossRef]
- Drucker, D.J.; Jin, T.; Asa, S.L.; Young, T.A.; Brubaker, P.L. Activation of proglucagon gene transcription by protein kinase-A in a novel mouse enteroendocrine cell line. Mol. Endocrinol. 1994, 8, 1646–1655. [Google Scholar] [CrossRef] [Green Version]
- Green, H.; Kehinde, O. An established preadipose cell line and its differentiation in culture II. Factors affecting the adipose conversion. Cell 1975, 5, 19–27. [Google Scholar] [CrossRef]
- McCarthy, T.; Green, B.D.; Calderwood, D.; Gillespie, A.; Cryan, J.F.; Giblin, L. STC-1 Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer International Publishing: Cham, Switzerland, 2015; pp. 211–220. ISBN 9783319161044. [Google Scholar]
- Nongonierma, A.B.; FitzGerald, R.J. Strategies for the discovery and identification of food protein-derived biologically active peptides. Trends Food Sci. Technol. 2017, 69, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S. The genetics of Caenorabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, Y.; Qin, C.; Liang, M.; Mao, X.; Li, S.; Zou, Y.; Jia, W.; Li, H.; Ma, C.W.; et al. Bioactive Peptides from Angelica sinensis Protein Hydrolyzate Delay Senescence in Caenorhabditis elegans through Antioxidant Activities. Oxid. Med. Cell. Longev. 2016, 2016, 8956981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Xu, Q.; Zhou, X.; Song, S.; Zhu, B. Stress resistance and lifespan extension of: Caenorhabditis elegans enhanced by peptides from mussel (Mytilus edulis) protein hydrolyzate. Food Funct. 2018, 9, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Yin, F.; Wang, L.; Wei, W.; Jiang, L.; Qin, J. Modeling type 2 diabetes-like hyperglycemia in C. elegans on a microdevice. Integr. Biol. (UK) 2016, 8, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Schlotterer, A.; Kukudov, G.; Bozorgmehr, F.; Hutter, H.; Du, X.; Oikonomou, D.; Ibrahim, Y.; Pfisterer, F.; Rabbani, N.; Thornalley, P.; et al. C. elegans as model for the study of high glucose-mediated life span reduction. Diabetes 2009, 58. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.; Vilcinskas, A. The Fruit Fly Drosophila melanogaster as a Model for Aging Research. In Yellow Biotechnology I: Insect Biotechnologie in Drug Discovery and Preclinical Research; Vilcinskas, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 63–77. ISBN 978-3-642-39863-6. [Google Scholar]
- Chen, S.; Yang, Q.; Chen, X.; Tian, Y.; Liu, Z.; Wang, S. Bioactive peptides derived from crimson snapper and in vivo anti-aging effects on fat diet-induced high fat Drosophila melanogaster. Food Funct. 2020, 11, 524–533. [Google Scholar] [CrossRef]
- Suárez, S.; Aphalo, P.; Rinaldi, G.; Añón, M.C.; Quiroga, A. Effect of amaranth proteins on the RAS system. In vitro, in vivo and ex vivo assays. Food Chem. 2020, 308, 125601. [Google Scholar] [CrossRef]
- King, A.J.F. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Kim, Y.Y.; Kim, B.; Nam, H.; Suh, J.G. Improving glycemic control in model mice with type 2 diabetes by increasing superoxide dismutase (SOD) activity using silk fibroin hydrolysate (SFH). Biochem. Biophys. Res. Commun. 2017, 493, 115–119. [Google Scholar] [CrossRef]
- Mochida, T.; Hira, T.; Hara, H. The corn protein, zein hydrolysate, administered into the ileum attenuates hyperglycemia via its dual action on glucagon-like peptide-1 secretion and dipeptidyl peptidase-IV activity in rats. Endocrinology 2010, 151, 3095–3104. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y.; Hira, T.; Inoue, D.; Harada, Y.; Hashimoto, H.; Fujii, M.; Kadowaki, M.; Hara, H. Rice protein hydrolysates stimulate GLP-1 secretion, reduce GLP-1 degradation, and lower the glycemic response in rats. Food Funct. 2015, 6, 2525–2534. [Google Scholar] [CrossRef]
- D’Souza, R.; Pandeya, D.R.; Rahman, M.; Lee, H.S.; Jung, J.K.; Hong, S.T. Genetic engineering of Lactococcus lactis to produce an amylase inhibitor for development of an anti-diabetes biodrug. New Microbiol. 2012, 35, 35–42. [Google Scholar] [PubMed]
- Wang, T.Y.; Hsieh, C.H.; Hung, C.C.; Jao, C.L.; Chen, M.C.; Hsu, K.C. Fish skin gelatin hydrolysates as dipeptidyl peptidase IV inhibitors and glucagon-like peptide-1 stimulators improve glycaemic control in diabetic rats: A comparison between warm-and cold-water fish. J. Funct. Foods 2015, 19, 330–340. [Google Scholar] [CrossRef]
- Drotningsvik, A.; Mjos, S.A.; Pampanin, D.M.; Slizyte, R.; Carvajal, A.; Remman, T.; Hogoy, I.; Gudbrandsen, O.A. Dietary fish protein hydrolysates containing bioactive motifs affect serum and adipose tissue fatty acid compositions, serum lipids, postprandial glucose regulation and growth in obese Zucker fa/fa rats. Br. J. Nutr. 2016, 116, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, D.A.; Merricks, E.P.; Nichols, T.C. Swine models of type 2 diabetes mellitus: Insulin resistance, glucose tolerance, and cardiovascular complications. ILAR J. 2006, 47, 243–258. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.Y.; Chung, H.J.; Kim, K.W.; Cho, K.H.; Choi, I.; Lee, H.T. Potential use of transgenic domestic pigs expressing recombinant human erythropoietin in diabetes translation research. Anim. Cells Syst. 2019, 23, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Ribel, U.; Larsen, M.O.; Rolin, B.; Carr, R.D.; Wilken, M.; Sturis, J.; Westergaard, L.; Deacon, C.F.; Knudsen, L.B. NN2211: A long-acting glucagon-like peptide-1 derivative with anti-diabetic effects in glucose-intolerant pigs. Eur. J. Pharmacol. 2002, 451, 217–225. [Google Scholar] [CrossRef]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Chalamaiah, M.; Keskin Ulug, S.; Hong, H.; Wu, J. Regulatory requirements of bioactive peptides (protein hydrolysates) from food proteins. J. Funct. Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Manders, R.J.F.; Hansen, D.; Zorenc, A.H.G.; Dendale, P.; Kloek, J.; Saris, W.H.M.; Van Loon, L.J.C. Protein co-ingestion strongly increases postprandial insulin secretion in type 2 diabetes patients. J. Med. Food 2014, 17, 758–763. [Google Scholar] [CrossRef]
- Manders, R.J.; Koopman, R.; Sluijsmans, W.E.; van den Berg, R.; Verbeek, K.; Saris, W.H.; Wagenmakers, A.J.; van Loon, L.J. Co-Ingestion of a Protein Hydrolysate with or without Additional Leucine Effectively Reduces Postprandial Blood Glucose Excursions in Type 2 Diabetic Men. J. Nutr. 2006, 136, 1294–1299. [Google Scholar] [CrossRef] [Green Version]
- Jonker, J.T.; Wijngaarden, M.A.; Kloek, J.; Groeneveld, Y.; Gerhardt, C.; Brand, R.; Kies, A.K.; Romijn, J.A.; Smit, J.W.A. Effects of low doses of casein hydrolysate on post-challenge glucose and insulin levels. Eur. J. Intern. Med. 2011, 22, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Geerts, B.F.; Van Dongen, M.G.J.; Flameling, B.; Moerland, M.M.; Kam, M.L.D.; Cohen, A.F.; Romijn, J.A.; Gerhardt, C.C.; Kloek, J.; Burggraaf, J. Hydrolyzed casein decreases postprandial glucose concentrations in T2DM patients irrespective of leucine content. J. Diet. Suppl. 2011, 8, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Crombach, N.; Gijsen, A.P.; Walrand, S.; Fauquant, J.; Kies, A.K.; Lemosquet, S.; Saris, W.H.; Boirie, Y.; van Loon, L.J. Ingestion of a protein hydrolysate is accompanied by an accelerated in vivo digestion and absorption rate when compared with its intact protein. Am. J. Clin. Nutr. 2009, 90, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, L.; Schrier, N.L.; Bruins, M.J.; Steegers, E.A.P.; van den Meiracker, A.H.; Visser, W. Effect of oral protein hydrolysate on glucose control in patients with gestational diabetes. Clin. Nutr. 2018, 37, 878–883. [Google Scholar] [CrossRef]
- Petersen, B.L.; Ward, L.S.; Bastian, E.D.; Jenkins, A.L.; Campbell, J.; Vuksan, V. A whey protein supplement decreases post-prandial glycemia. Nutr. J. 2009, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Frid, A.H.; Nilsson, M.; Holst, J.J.; Björck, I.M.E. Effect of whey on blood glucose and insulin responses to composite breakfast and lunch meals in type 2 diabetic subjects. Am. J. Clin. Nutr. 2005, 82, 69–75. [Google Scholar] [CrossRef]
- Sartorius, T.; Weidner, A.; Dharsono, T.; Boulier, A.; Wilhelm, M.; Schön, C. Postprandial Effects of a Proprietary Milk Protein Hydrolysate Containing Bioactive Peptides in Prediabetic Subjects. Nutrients 2019, 11, 1700. [Google Scholar] [CrossRef] [Green Version]
- Goudarzi, M.; Madadlou, A. Influence of whey protein and its hydrolysate on prehypertension andpostprandial hyperglycaemia in adult men. Int. Dairy J. 2013, 33, 62–66. [Google Scholar] [CrossRef]
- Plat, J.; Severins, N.; Mensink, R.P. Improvement of pulse wave velocity and metabolic cardiovascular risk parameters through egg protein hydrolysate intake: A randomized trial in overweight or obese subjects with impaired glucose tolerance or type 2 diabetes. J. Funct. Foods 2019, 52, 418–423. [Google Scholar] [CrossRef]
- Calbet, J.A.L.; Holst, J.J. Gastric emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur. J. Nutr. 2004, 43, 127–139. [Google Scholar] [CrossRef]
- Zhu, C.F.; Li, G.Z.; Peng, H.-B.; Zhang, F.; Chen, Y.; Li, Y. Treatment with marine collagen peptides modulates glucose and lipid metabolism in chinese patients with type 2 diabetes mellitus. Appl. Physiol. Nutr. Metab. 2010, 35, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Hovland, I.H.; Leikanger, I.S.; Stokkeland, O.; Waage, K.H.; Mjøs, S.A.; Brokstad, K.A.; McCann, A.; Ueland, P.M.; Slizyte, R.; Carvajal, A.; et al. Effects of low doses of fish and milk proteins on glucose regulation and markers of insulin sensitivity in overweight adults: A randomised, double blind study. Eur. J. Nutr. 2020, 59, 1013–1029. [Google Scholar] [CrossRef] [PubMed]
- Dale, H.F.; Jensen, C.; Hausken, T.; Lied, E.; Hatlebakk, J.G.; Brønstad, I.; Lihaug Hoff, D.A.; Lied, G.A. Effect of a cod protein hydrolysate on postprandial glucose metabolism in healthy subjects: A double-blind cross-over trial. J. Nutr. Sci. 2018, 7, e33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowe, W.; McLaughlin, C.M.; Allsopp, P.J.; Slevin, M.M.; Harnedy, P.A.; Cassidy, Y.; Baird, J.; Devaney, M.; Fitzgerald, R.J.; O’Harte, F.P.M.; et al. The effect of boarfish protein hydrolysate on postprandial glycaemic response and satiety in healthy adults. Proc. Nutr. Soc. 2018, 77. [Google Scholar] [CrossRef] [Green Version]
- Curran, A.M.; Horner, K.; O’Sullivan, V.; Nongonierma, A.B.; Le Maux, S.; Murphy, E.; Kelly, P.; Fitzgerald, R.J.; Brennan, L. Variable Glycemic Responses to Intact and Hydrolyzed Milk Proteins in Overweight and Obese Adults Reveal the Need for Precision Nutrition. J. Nutr. 2019, 149, 88–97. [Google Scholar] [CrossRef]
- Lee, S.K.; Posthauer, M.E.; Dorner, B.; Redovian, V.; Maloney, M.J. Pressure ulcer healing with a concentrated, fortified, collagen protein hydrolysate supplement: A randomized controlled trial. Adv. Skin Wound Care 2006, 19, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Hillesheim, E.; Brennan, L. Metabotyping and its role in nutrition research. Nutr. Res. Rev. 2020, 33, 33–42. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Amino Acid | Quinoa | Lentil Protein Isolate | Brewer Spent Grain | Mealworm Larvae Meal | Silkworm Pupae Meal | Mussel Meal | Herring |
|---|---|---|---|---|---|---|---|
| Essential | |||||||
| H | 2.2 | 2.0 | 3.6 | 2.9 | 2.6 | 1.9 | 2.1 |
| I | 0.8 | 3.1 | 4.2 | 4.7 | 5.1 | 4.5 | 3.3 |
| L | 2.5 | 6.9 | 7.2 | 8.0 | 7.5 | 7.2 | 7.9 |
| K | 2.3 | 5.6 | 3.1 | 6.3 | 7 | 8.3 | 10.1 |
| M | 0.3 | 0.6 | 1.4 | 1.4 | 3.5 | 2.6 | 3.3 |
| F + Y | 2.8 | 7.1 | 9.7 | 9.5 | 11.1 | 8.7 | 5.9 |
| T | 5.7 | 3.0 | 3.2 | 4.3 | 5.1 | 5.3 | 4.0 |
| W | 1.0 | - | 1.2 | 0.9 | 1.0 | - | |
| V | 1.0 | 3.5 | 6.0 | 8.5 | 5.5 | 4.6 | 4.6 |
| C | 0.1 | 0.5 | 1.4 | 0.8 | 1 | 1.0 | 1.1 |
| Non-essential | |||||||
| R | 3.0 | 7.4 | 5.9 | 5.4 | 5.6 | 7.6 | 7.5 |
| G | 3.0 | 3.1 | 3.8 | 5.5 | 4.8 | 6.6 | 7.6 |
| E | 8.7 | 15.5 | 24.8 | 10.6 | 13.9 | 14.0 | 17.1 |
| D | 3.7 | 10.5 | 6.6 | 7.8 | 10.4 | 11.3 | 9.3 |
| P | 1.8 | 2.9 | 9.7 | 6.0 | 5.2 | 4.2 | 4.7 |
| S | 1.7 | 5.2 | 4.1 | 4.6 | 5.0 | 5.4 | 4.3 |
| A | 2.2 | 3.4 | 4.3 | 8.4 | 5.8 | 5.1 | 7.1 |
| Ref. | [89] | [90] | [91] | [92] | [93] | [94] | [95] |
| In Vitro | Cellular Assay | In Vivo | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Substrate | Enzymatic Treatment | ID | Verification | B-A | DPP-IV | GIA | AMY | Cell line | Model | Ref |
| Rainbow trout (Oncorhynchus mykiss) | Alcalase | No | No | No | Yes | No | No | No | No | [30] |
| Camel whey protein | PTN 6.0S | Yes | Yes | Yes | Yes | No | No | No | No | [57] |
| Boarfish (Capros aper) | Alcalase 2.4 L, Flavourzyme 500 L; Simulated digestion | Yes | Yes | No | Yes | No | No | Caco-2; BRIN-BD11 | No | [59] |
| Blue whiting (Micromesistius poutassou) | Alcalase 2.4 L and Flavourzyme 500 L Simulated digestion | Yes | No | No | Yes | No | No | BRIN-BD11, GLUTag, 3T3-L1 | NIH Swiss mice | [81] |
| Cricket (G. sigillatus) | Alcalase Simulated digestion | No | No | No | Yes | No | No | No | No | [96] |
| Soybean (Glycine max) | Simulated digestion | Yes | No | No | Yes | Yes | Yes | No | No | [97] |
| Luffa cylindrical seed | Alcalase, trypsin | No | No | No | No | Yes | Yes | No | No | [76] |
| Salmon (Salmo salar) | Alcalase 2.4 L, Alcalase 2.4 L and Flavourzyme 500 L, and Promod 144 MG | Yes | No | No | Yes | No | No | BRIN-BD11; GLUTag | No | [73] |
| Boarfish (Capros aper) | Alcalase 2.4 L, Flavourzyme 500 L Simulated digestion | No | No | No | Yes | No | No | Caco-2; BRIN-BD11; GLUTag; 3T3-L1 | Mice | [98] |
| Mealworm (Tenebrio molitor) | Alcalase, trypsin, ficin, flavourzyme | No | No | No | Yes | No | No | No | No | [74] |
| Tropical banded crickets (Gryllodes sigillatus) | Protamex Simulated digestion | No | No | No | Yes | No | No | No | No | [99] |
| Hempseed (Cannabis sativa) | Pepsin, trypsin | No | No | No | Yes | No | No | Yes | Ex-vivo | [100] |
| Bovine whey | Trypsin | Yes | Yes | No | Yes | No | No | No | No | [101] |
| Sea cucumber (Stichopus japonicus) | Simulated gastrointestinal digestion | Yes | No | Yes | Yes | No | No | 3T3-L1, HepG2 | No | [102] |
| Casein | PROTIN SD-NY10 | No | No | No | No | Yes | Yes | No | No | [103] |
| Walnut (Juglans mandshurica) | Alcalase 2.4 L | Yes | Yes | Yes | No | Yes | Yes | HepG2 cells | No | [104] |
| Mealworm (T. molitor), locust (Schistocerca gregaria), cricket (G. sigilatus) | Simulated digestion | Yes | Yes | No | No | Yes | No | No | No | [105] |
| Corn germ | Alcalase, flavourzyme, trypsin | No | No | No | Yes | Yes | Yes | No | No | [106] |
| Millet grains (Panicum miliaceum) | Simulated digestion | Yes | No | No | No | Yes | Yes | No | No | [107] |
| Cowpea (Vigna unguiculata) | Alcalase + Flavourzyme | No | No | No | Yes | Yes | Yes | No | No | [108] |
| Beans (Phaseolus vulgaris) | Simulated digestion | No | No | No | No | Yes | Yes | No | Male wistar rats | [109] |
| Basil seeds (Ocimum tenuriflorum) | Pepsin | No | No | No | No | No | Yes | No | No | [110] |
| Brewers’ spent grain | Alcalase + Flavourzme Simulated digestion | Yes | Yes | No | Yes | No | No | No | No | [111] |
| Porphyra dioica extracted protein | Alcalase + Flavourzyme | Yes | Yes | No | Yes | No | No | No | No | [112] |
| Red Seaweed (Porphyra spp) | Alcalase, neutrase, pepsin, and trypsin | Yes | Yes | No | No | No | Yes | No | No | [113] |
| Soybean (Glycine max) | Trypsin | Yes | Yes | Yes | No | Yes | No | No | Mice | [114] |
| Rice albumin (Oryza sativa japonica) | Trypsin | No | No | No | No | No | No | STC-1 | Wistar rats | [115] |
| Tuber storage proteins | Simulated digestion (In silico) | Yes | No | Yes | No | No | No | No | No | [116] |
| Rambutan (Nephelium lappaceum), pulasan (N. mutabile) | Simulated digestion | Yes | No | Yes | No | No | Yes | No | No | [117] |
| Pinto beans (P. vulgaris) | Protamex | Yes | Yes | Yes | No | No | Yes | No | No | [118] |
| Egg white ovoalbumin | Simulated digestion (In silico) | Yes | Yes | Yes | Yes | No | No | No | No | [119] |
| Salmon skin collagen (Salmo salar) | Pepsin, trypsin, papain, Alcalase 2.4 L | Yes | Yes | Yes | Yes | No | No | No | No | [120] |
| Quinoa (Chenopodium quinoa) | Papain, ficin, bromelain (In silico) | Yes | Yes | Yes | Yes | No | No | No | No | [121] |
| Spirulina (Arthrospira platensis) | Tryspin | Yes | No | No | Yes | No | No | Caco-2 | No | [122] |
| Tomato seed proteins (Solanum lycopersicum) | 15 enzymes (In silico) | Yes | No | No | No | No | No | No | No | [123] |
| Egg | Pepsin, trypsin (in silico) | Yes | Yes | Yes | Yes | No | No | No | No | [124] |
| Common carp (Cyprinus carpio) | Papain, neutrase, trypsin, pepsin; Simulated digestion | Yes | Yes | Yes | Yes | No | No | Caco-2 HepG2 | No | [125] |
| Spirulina (Spirulina platensis) | Trypsin, pepsin | Yes | No | No | Yes | No | No | Caco-2 | No | [126] |
| Pea (Pisum sativum) | Alcalase, neutrase | Yes | No | No | No | No | No | No | Male Kunming mice | [127] |
| Buffalo colostrum (Bubalus bubalis) | Simulated digestion | Yes | Yes | Yes | Yes | No | No | No | No | [128] |
| Chicken feet (Gallus gallus domesticus) | Neutrase, Protamex | No | No | No | Yes | No | No | STC-1 | Wistar Rats | [129] |
| Portuguese Oyster (Crassostrea angulata) | Pepsin, bromelain, papain | Yes | No | Yes | Yes | No | No | No | No | [130] |
| Casein | Alcalase, protamex, neutrase, bromelain, and papain | Yes | Yes | Yes | Yes | No | No | No | Male Kun Ming mice | [131] |
| Whey | Corolase 2TS, Protamex | No | No | No | Yes | No | No | No | No | [132] |
| Soy (Glycine max) | Alkaline proteinase, papain, trypsin; Simulated digestion | Yes | Yes | No | Yes | Yes | No | No | No | [133] |
| Egg | Simulated digestion | Yes | Yes | Yes | Yes | No | No | Caco-2 | Wistar rats | [134] |
| Rapeseed (Brassica napus) | Alcalase, trypsin pepsin, flavourzyme, papain | Yes | Yes | Yes | Yes | No | No | No | No | [135] |
| Lesser mealworm (A. diaperinus) | Simulated digestion; alcalase, Flavourzyme, papain, and thermolysin | Yes | No | No | Yes | No | No | No | No | [136] |
| Camel skin gelatin (Camelius dromedarius) | Alcalase, protease from S. Griseus | No | No | No | Yes | No | Yes | No | No | [137] |
| Chicken (Gallus gallus) | Corolase, Flavourzyme | Yes | No | No | Yes | No | No | Skeletal muscle | No | [138] |
| Kiwicha (Amaranthus caudatus) | Simulated digestion | Yes | No | No | Yes | No | Yes | Caco-2 | No | [139] |
| Silver carp (Hypophthalmichthys molitrix) | Alcalase 2.4 L, neutrase, pepsin, trypsin, Flavourzyme | Yes | Yes | Yes | Yes | No | No | No | No | [140] |
| Flaxseed (Linum usitatissimum), rapeseed (Brassica napus), sunflower (Helianthus annuus), sesame (Sesamum indicum), soybean (Glycine max) | Subtilisin, pepsin, pepsin (In silico) | Yes | No | Yes | No | No | No | No | No | [141] |
| Bambara bean (Vigna subterranean) | Alcalase, thermolysin, trypsin Simulated digestion | Yes | No | Yes | Yes | No | No | No | No | [142] |
| Mealworm (T. molitor) | Pepsin, papain (In silico and experimental) | Yes | No | No | Yes | No | No | No | No | [143] |
| Yellow field pea (Pisum sativum) | Alcalase, chymotrypsin, pepsin, trypsin | Yes | No | No | No | Yes | Yes | No | No | [144] |
| Sardine (Sardine pilchardus) | Alcalase, Trypsin, Flavourzyme | Yes | No | Yes | Yes | No | No | No | No | [84] |
| Sodium caseinate | Simulated digestion | Yes | Yes | No | No | No | No | BRIN-BD11, 3T3-L1 | Mice | [145] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Antidiabetic Food-Derived Peptides for Functional Feeding: Production, Functionality and In Vivo Evidences. Foods 2020, 9, 983. https://doi.org/10.3390/foods9080983
Rivero-Pino F, Espejo-Carpio FJ, Guadix EM. Antidiabetic Food-Derived Peptides for Functional Feeding: Production, Functionality and In Vivo Evidences. Foods. 2020; 9(8):983. https://doi.org/10.3390/foods9080983
Chicago/Turabian StyleRivero-Pino, Fernando, F. Javier Espejo-Carpio, and Emilia M. Guadix. 2020. "Antidiabetic Food-Derived Peptides for Functional Feeding: Production, Functionality and In Vivo Evidences" Foods 9, no. 8: 983. https://doi.org/10.3390/foods9080983
APA StyleRivero-Pino, F., Espejo-Carpio, F. J., & Guadix, E. M. (2020). Antidiabetic Food-Derived Peptides for Functional Feeding: Production, Functionality and In Vivo Evidences. Foods, 9(8), 983. https://doi.org/10.3390/foods9080983