
Triple Silencing of HSP27, cFLIP, and CLU Genes Promotes the Sensitivity of Doxazosin-Induced Apoptosis in PC-3 Prostate Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Doxazosin Treatment
2.2. MTT Assay
2.3. Annexin-V Staining
2.4. siRNA Silencing of HSP27, cFLIP, and CLU Genes
2.5. Total RNA Extraction and qRT-PCR
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
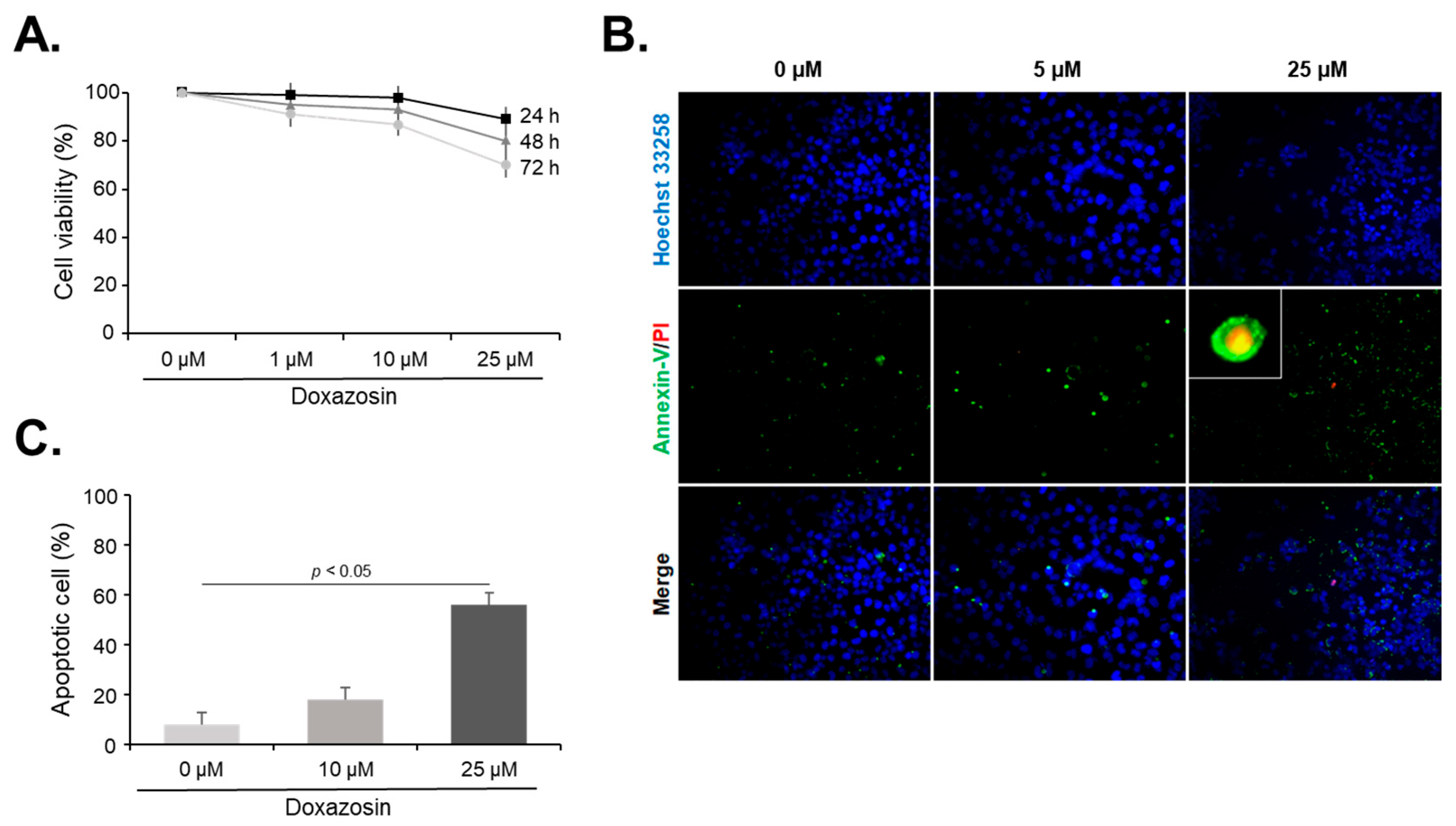
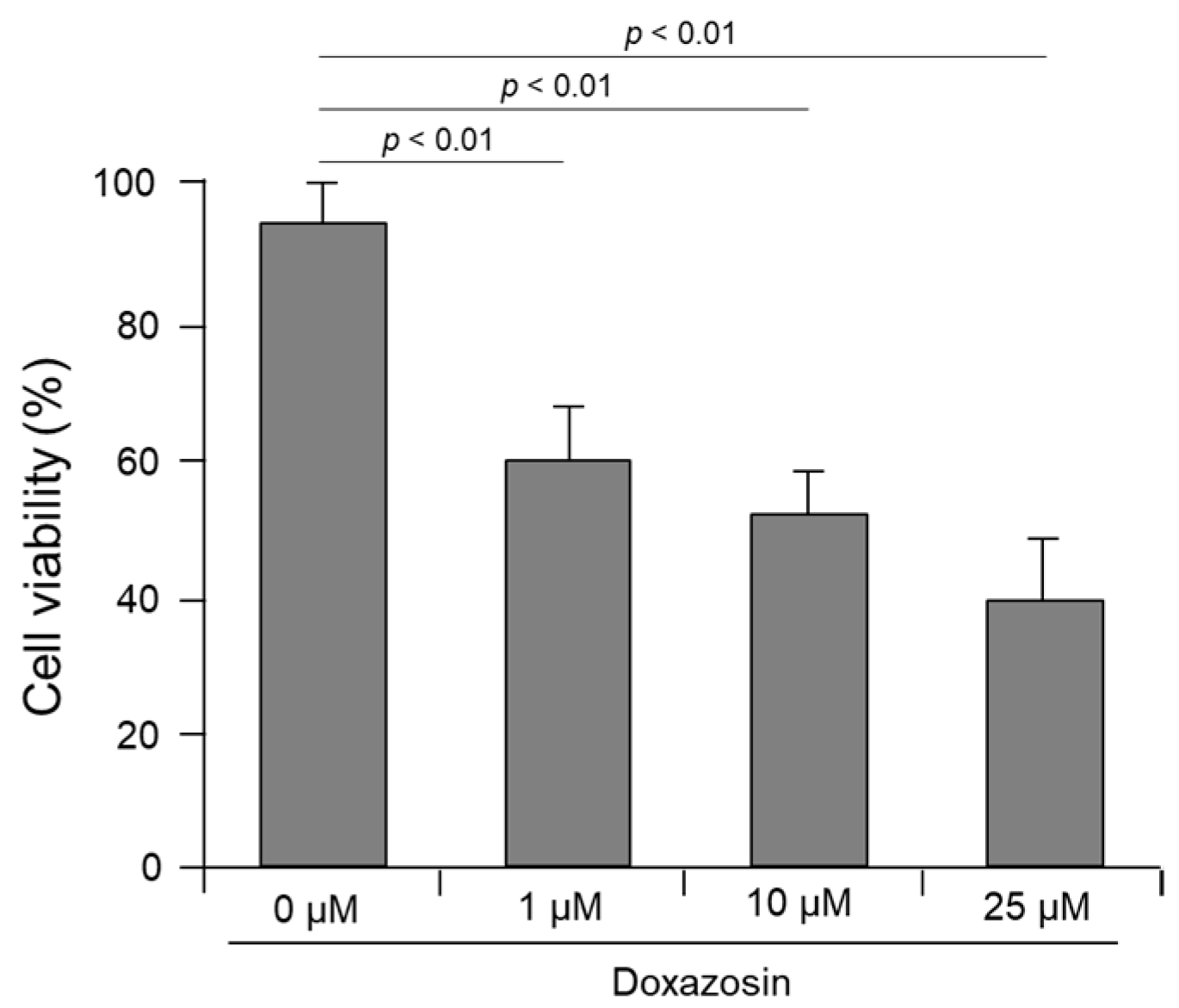
3.1. Cell Viability and Apoptosis after Doxazosin Treatment
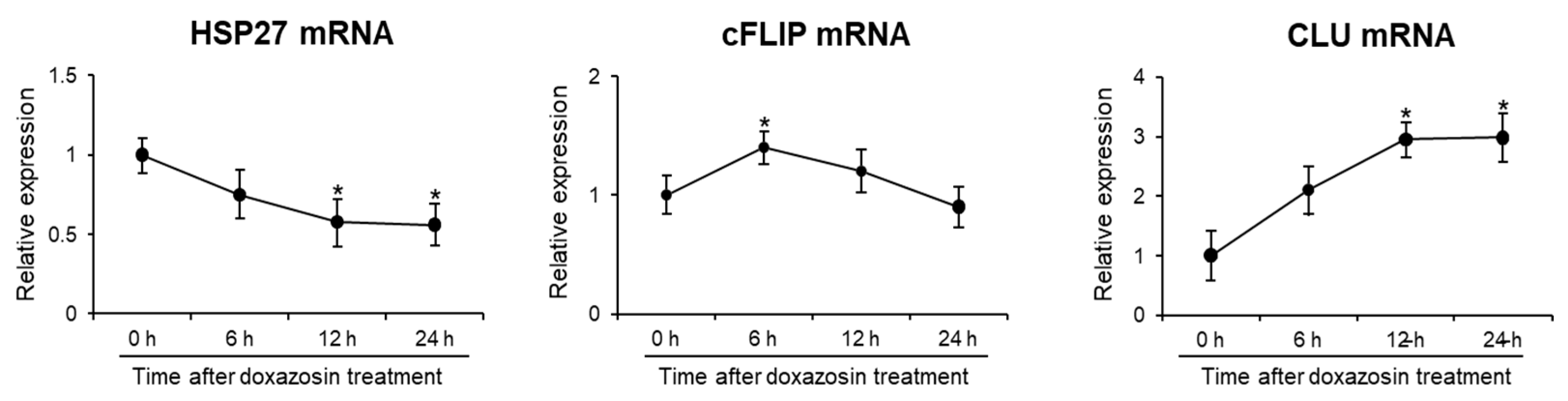
3.2. Expression of Survival-Related Genes after Doxazosin Treatment
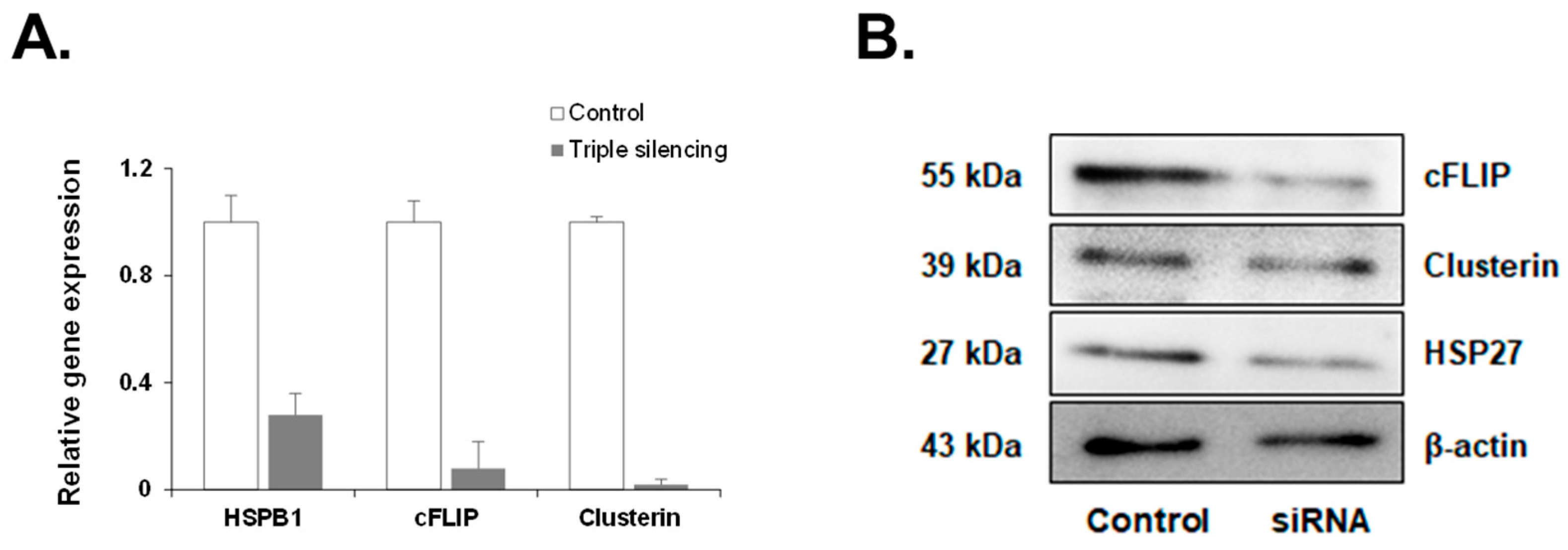
3.3. siRNA Efficiency of HSP27, cFLIP, and CLU Genes
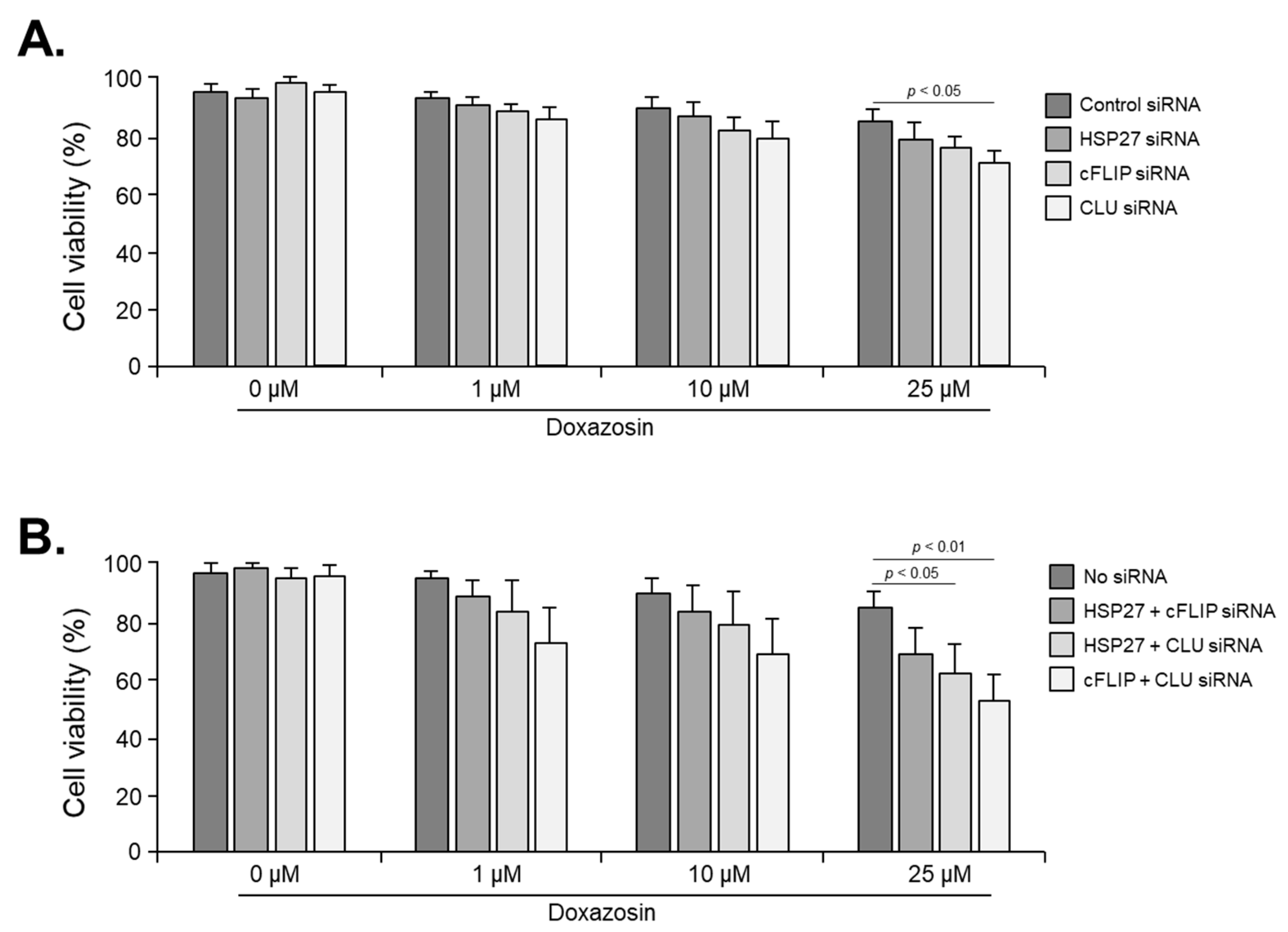
3.4. Cell Viability after siRNA of HSP27, cFLIP, and CLU Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amaral, T.M.; Macedo, D.; Fernandes, I.; Costa, L. Castration-resistant prostate cancer: Mechanisms, targets, and treatment. Prostate Cancer 2012, 2012, 327253. [Google Scholar] [CrossRef]
- Chandrasekar, T.; Yang, J.C.; Gao, A.C.; Evans, C.P. Mechanisms of resistance in castration-resistant prostate cancer (CRPC). Transl. Androl. Urol. 2015, 4, 365–380. [Google Scholar] [PubMed]
- Heidegger, I.; Massoner, P.; Eder, I.E.; Pircher, A.; Pichler, R.; Aigner, F.; Bektic, J.; Horninger, W.; Klocker, H. Novel therapeutic approaches for the treatment of castration-resistant prostate cancer. J. Steroid Biochem. Mol. Biol. 2013, 138, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Wade, C.A.; Goodwin, J.; Preston, D.; Kyprianou, N. Impact of α-adrenoceptor antagonists on prostate cancer development, progression and prevention. Am. J. Clin. Exp. Urol. 2019, 7, 46–60. [Google Scholar]
- Archer, M.; Dogra, N.; Dovey, Z.; Ganta, T.; Jang, H.S.; Khusid, J.A.; Lantz, A.; Mihalopoulos, M.; Stockert, J.A.; Zahalka, A.; et al. Role of α- and β-adrenergic signaling in phenotypic targeting: Significance in benign and malignant urologic disease. Cell Commun. Signal. 2021, 19, 78. [Google Scholar] [CrossRef]
- Partin, J.V.; Anglin, I.E.; Kyprianou, N. Quinazoline-based alpha 1-adrenoceptor antagonists induce prostate cancer cell apoptosis via TGF-beta signalling and I kappa B alpha induction. Br. J. Cancer 2003, 88, 1615–1621. [Google Scholar] [CrossRef]
- Tahmatzopoulos, A.; Kyprianou, N. Apoptotic impact of alpha1-blockers on prostate cancer growth: A myth or an inviting reality? Prostate 2004, 59, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Anglin, I.E.; Glassman, D.T.; Kyprianou, N. Induction of prostate apoptosis by alpha1-adrenoceptor antagonists: Mechanistic significance of the quinazoline component. Prostate Cancer Prostatic Dis. 2002, 5, 88–95. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Li, A.; Huang, X.; Bai, G.L.; Jiang, Y.X.; Li, R.L.; Liu, C.; Wen, Z.Y.; Wang, P.; Chen, A.J. HSP27 protects skin from ultraviolet B-induced photodamage by regulating autophagy and reactive oxygen species production. Front. Cell Dev. Biol. 2022, 10, 852244. [Google Scholar] [CrossRef]
- Bi, X.; Jiang, B.; Zhou, J.; Luo, L.; Yin, Z. Phosphorylated Hsp27 prevents LPS-induced excessive inflammation in THP-1 cells via suppressing ROS-mediated upregulation of, C.B.P. Cell Biol. Int. 2020, 44, 253–267. [Google Scholar] [CrossRef]
- Garrido, C.; Schmitt, E.; Candé, C.; Vahsen, N.; Parcellier, A.; Kroemer, G. HSP27 and HSP70, potentially oncogenic apoptosis inhibitors. Cell Cycle 2003, 2, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Bruey, J.M.; Ducasse, C.; Bonniaud, P.; Ravagnan, L.; Susin, S.A.; Diaz-Latoud, C.; Gurbuxani, S.; Arrigo, A.P.; Kroemer, G.; Solary, E.; et al. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nat. Cell Biol. 2000, 2, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Moon, S.P.; Cutolo, G.; Javed, A.; Ahn, B.S.; Ryu, A.H.; Pratt, M.R. HSP27 inhibitory activity against caspase-3 cleavage and activation by caspase-9 is enhanced by chaperone O-GlcNAc modification in vitro. ACS Chem. Biol. 2023, 18, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, P.; Jugpal, P.; So, A.; Sinneman, S.; Ettinger, S.; Fazli, L.; Nelson, C.; Gleave, M. Small interference RNA targeting heat-shock protein 27 inhibits the growth of prostatic cell lines and induces apoptosis via caspase-3 activation in vitro. BJU Int. 2006, 98, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Ottavi, P.; Fromentin, A.; Hammann, A.; Arrigo, A.P.; Chauffert, B.; Mehlen, P. HSP27 as a mediator of confluence-dependent resistance to cell death induced by anticancer drugs. Cancer Res. 1997, 57, 2661–2667. [Google Scholar]
- Arts, H.J.; Hollema, H.; Lemstra, W.; Willemse, P.H.; De Vries, E.G.; Kampinga, H.H.; Van der Zee, A.G. Heat-shock-protein-27 (HSP27) expression in ovarian carcinoma: Relation in response to chemotherapy and prognosis. Int. J. Cancer 1999, 84, 234–238. [Google Scholar] [CrossRef]
- Safa, A.R. cFLIP, a master anti-apoptotic regulator. Exp. Oncol. 2012, 34, 176–184. [Google Scholar]
- Oztürk, S.; Schleich, K.; Lavrik, I.N. Cellular FLICE-like inhibitory proteins (cFLIPs): Fine-tuners of life and death decisions. Exp. Cell Res. 2012, 318, 1324–1331. [Google Scholar] [CrossRef]
- Hwang, E.Y.; Jeong, M.S.; Park, S.Y.; Jang, S.B. Evidence of complex formation between FADD and cFLIP death effector domains for the death inducing signaling complex. BMB Rep. 2014, 47, 488–493. [Google Scholar] [CrossRef]
- Haag, C.; Stadel, D.; Zhou, S.; Bachem, M.G.; Möller, P.; Debatin, K.M.; Fulda, S. Identification of cFLIP(L) and cFLIP(S) as critical regulators of death receptor-induced apoptosis in pancreatic cancer cells. Gut 2011, 60, 225–237. [Google Scholar] [CrossRef]
- Shannan, B.; Seifert, M.; Boothman, D.A.; Tilgen, W.; Reichrath, J. Clusterin and DNA repair: A new function in cancer for a key player in apoptosis and cell cycle control. J. Mol. Histol. 2006, 37, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Viard, I.; Wehrli, P.; Jornot, L.; Bullani, R.; Vechietti, J.L.; Schifferli, J.A.; Tschopp, J.; French, L.E. Clusterin gene expression mediates resistance to apoptotic cell death induced by heat shock and oxidative stress. J. Investig. Dermatol. 1999, 112, 290–296. [Google Scholar] [CrossRef] [PubMed]
- García-Aranda, M.; Téllez, T.; Muñoz, M.; Redondo, M. Clusterin inhibition mediates sensitivity to chemotherapy and radiotherapy in human cancer. Anticancer Drugs 2017, 28, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, T.; Kiyama, S.; Chi, K.; Miyake, H.; Adomat, H.; Skov, K.; Gleave, M.E. Overexpression of the cytoprotective protein clusterin decreases radiosensitivity in the human LNCaP prostate tumour model. BJU Int. 2003, 92, 463–469. [Google Scholar] [CrossRef]
- Shannan, B.; Seifert, M.; Leskov, K.; Willis, J.; Boothman, D.; Tilgen, W.; Reichrath, J. Challenge and promise: Roles for clusterin in pathogenesis, progression and therapy of cancer. Cell Death Differ. 2006, 13, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Kumano, M.; Beraldi, E.; Fazli, L.; Du, C.; Moore, S.; Sorensen, P.; Zoubeidi, A.; Gleave, M.E. Clusterin facilitates stress-induced lipidation of LC3 and autophagosome biogenesis to enhance cancer cell survival. Nat. Commun. 2014, 5, 5775. [Google Scholar] [CrossRef] [PubMed]
- Scaltriti, M.; Bettuzzi, S.; Sharrard, R.M.; Caporali, A.; Caccamo, A.E.; Maitland, N.J. Clusterin overexpression in both malignant and nonmalignant prostate epithelial cells induces cell cycle arrest and apoptosis. Br. J. Cancer 2004, 91, 1842–1850. [Google Scholar] [CrossRef]
- Benning, C.M.; Kyprianou, N. Quinazoline-derived alpha1-adrenoceptor antagonists induce prostate cancer cell apoptosis via an alpha1-adrenoceptor-independent action. Cancer Res. 2002, 62, 597–602. [Google Scholar]
- Garrison, J.B.; Kyprianou, N. Docetaxel induces apoptosis of benign and malignant prostate cells via a death receptor-mediated pathway. Cancer Res. 2006, 66, 464–472. [Google Scholar] [CrossRef]
- Karaca, B.; Bakır, E.; Yerer, M.B.; Cumaoğlu, A.; Hamurcu, Z.; Eken, A. Doxazosin and erlotinib have anticancer effects in the endometrial cancer cell and important roles in ERα and Wnt/β-catenin signaling pathways. J. Biochem. Mol. Toxicol. 2021, 35, e22905. [Google Scholar] [CrossRef] [PubMed]
- Karademir, D.; Özgür, A. Small molecule heat shock protein 27 inhibitor J2 decreases ovarian cancer cell proliferation via induction of apoptotic pathways. Med. Oncol. 2023, 40, 250. [Google Scholar] [CrossRef]
- Lampros, M.; Vlachos, N.; Voulgaris, S.; Alexiou, G.A. The Role of Hsp27 in Chemotherapy Resistance. Biomedicines 2022, 10, 897. [Google Scholar] [CrossRef]
- Singh, M.K.; Sharma, B.; Tiwari, P.K. The small heat shock protein Hsp27, Present understanding and future prospects. J. Therm. Biol. 2017, 69, 149–154. [Google Scholar] [CrossRef]
- Davidovich, P.; Higgins, C.A.; Najda, Z.; Longley, D.B.; Martin, S.J. cFLIPL acts as a suppressor of TRAIL- and Fas-initiated inflammation by inhibiting assembly of caspase-8/FADD/RIPK1 NF-κB-activating complexes. Cell Rep. 2023, 42, 113476. [Google Scholar] [CrossRef]
- Ranjan, K.; Surolia, A.; Pathak, C. Apoptotic potential of Fas-associated death domain on regulation of cell death regulatory protein cFLIP and death receptor mediated apoptosis in HEK 293T cells. J. Cell Commun. Signal. 2012, 6, 155–168. [Google Scholar] [CrossRef]
- Zou, W.; Chen, S.; Liu, X.; Yue, P.; Sporn, M.B.; Khuri, F.R.; Sun, S.Y. cFLIP downregulation contributes to apoptosis induction by the novel synthetic triterpenoid methyl-2-cyano-3, 12-dioxooleana-1, 9-dien-28-oate (CDDO-Me) in human lung cancer cells. Cancer Biol. Ther. 2007, 6, 1614–1620. [Google Scholar] [CrossRef]
- Zhang, S.; Shen, H.M.; Ong, C.N. Down-regulation of cFLIP contributes to the sensitization effect of 3,3′-diindolylmethane on TRAIL-induced apoptosis in cancer cells. Mol. Cancer Ther. 2005, 4, 1972–1981. [Google Scholar] [CrossRef] [PubMed]
- Cunin, P.; Beauvillain, C.; Miot, C.; Augusto, J.F.; Preisser, L.; Blanchard, S.; Pignon, P.; Scotet, M.; Garo, E.; Fremaux, I.; et al. Clusterin facilitates apoptotic cell clearance and prevents apoptotic cell-induced autoimmune responses. Cell Death Dis. 2016, 7, e2215. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, F.; Bettuzzi, S. The clusterin paradigm in prostate and breast carcinogenesis. Endocr. Relat. Cancer 2010, 17, R1–R17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cao, W.; Su, Q.; Liu, Z.; Zhang, L. Clusterin silencing inhibits proliferation and reduces invasion in human laryngeal squamous carcinoma cells. World J. Surg. Oncol. 2014, 12, 124. [Google Scholar] [CrossRef] [PubMed]
- Qiao, P.; Tian, Z. Atractylenolide I inhibits EMT and enhances the antitumor effect of cabozantinib in prostate cancer via targeting Hsp27. Front. Oncol. 2023, 12, 1084884. [Google Scholar] [CrossRef]
- Keledjian, K.; Kyprianou, N. Anoikis induction by quinazoline based alpha 1-adrenoceptor antagonists in prostate cancer cells: Antagonistic effect of bcl-2. J. Urol. 2003, 169, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Min, K.J.; Jang, J.H.; Lee, J.T.; Choi, K.S.; Kwon, T.K. Glucocorticoid receptor antagonist sensitizes TRAIL-induced apoptosis in renal carcinoma cells through up-regulation of DR5 and down-regulation of c-FLIP(L) and Bcl-2. J. Mol. Med. 2012, 90, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Koltai, T. Clusterin: A key player in cancer chemoresistance and its inhibition. OncoTargets Ther. 2014, 7, 447–456. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.M.; Sun, S.; Im, E.; Yang, H.; Yoo, T.K. Triple Silencing of HSP27, cFLIP, and CLU Genes Promotes the Sensitivity of Doxazosin-Induced Apoptosis in PC-3 Prostate Cancer Cells. Medicines 2024, 11, 7. https://doi.org/10.3390/medicines11030007
Cho JM, Sun S, Im E, Yang H, Yoo TK. Triple Silencing of HSP27, cFLIP, and CLU Genes Promotes the Sensitivity of Doxazosin-Induced Apoptosis in PC-3 Prostate Cancer Cells. Medicines. 2024; 11(3):7. https://doi.org/10.3390/medicines11030007
Chicago/Turabian StyleCho, Jeong Man, Sojung Sun, Eunji Im, Hyunwon Yang, and Tag Keun Yoo. 2024. "Triple Silencing of HSP27, cFLIP, and CLU Genes Promotes the Sensitivity of Doxazosin-Induced Apoptosis in PC-3 Prostate Cancer Cells" Medicines 11, no. 3: 7. https://doi.org/10.3390/medicines11030007
APA StyleCho, J. M., Sun, S., Im, E., Yang, H., & Yoo, T. K. (2024). Triple Silencing of HSP27, cFLIP, and CLU Genes Promotes the Sensitivity of Doxazosin-Induced Apoptosis in PC-3 Prostate Cancer Cells. Medicines, 11(3), 7. https://doi.org/10.3390/medicines11030007