
Critical Analysis on Characterization, Systemic Effect, and Therapeutic Potential of Beta-Sitosterol: A Plant-Derived Orphan Phytosterol

 ,
,
Abstract
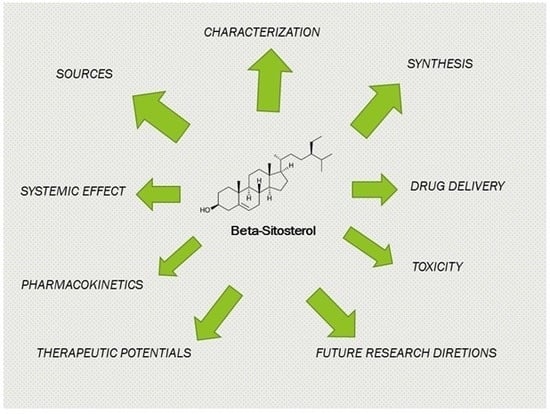
:
1. Introduction
2. Sources
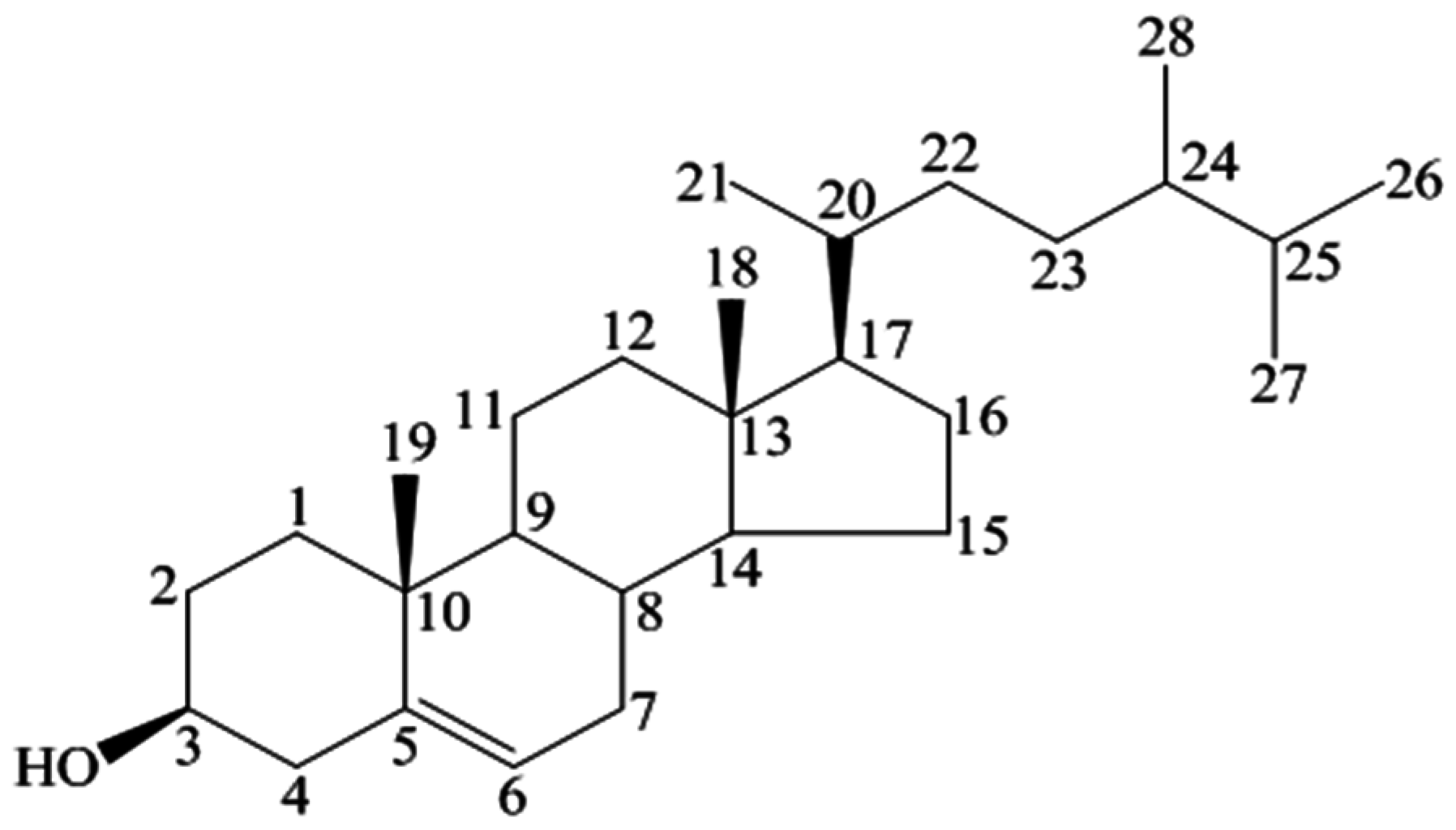
3. Characterization
3.1. Physical and Chemical Properties
3.2. Spectral Analysis
3.2.1. IR Spectral Analysis
3.2.2. NMR Spectral Analysis
3.3. Chromatographic Analysis
3.3.1. Thin Layer Chromatography (TLC)
3.3.2. Gas Layer Chromatography
3.3.3. High-Performance Liquid Chromatography
3.3.4. Gas Chromatography Mass Spectrometry (GCMS)
4. Synthesis
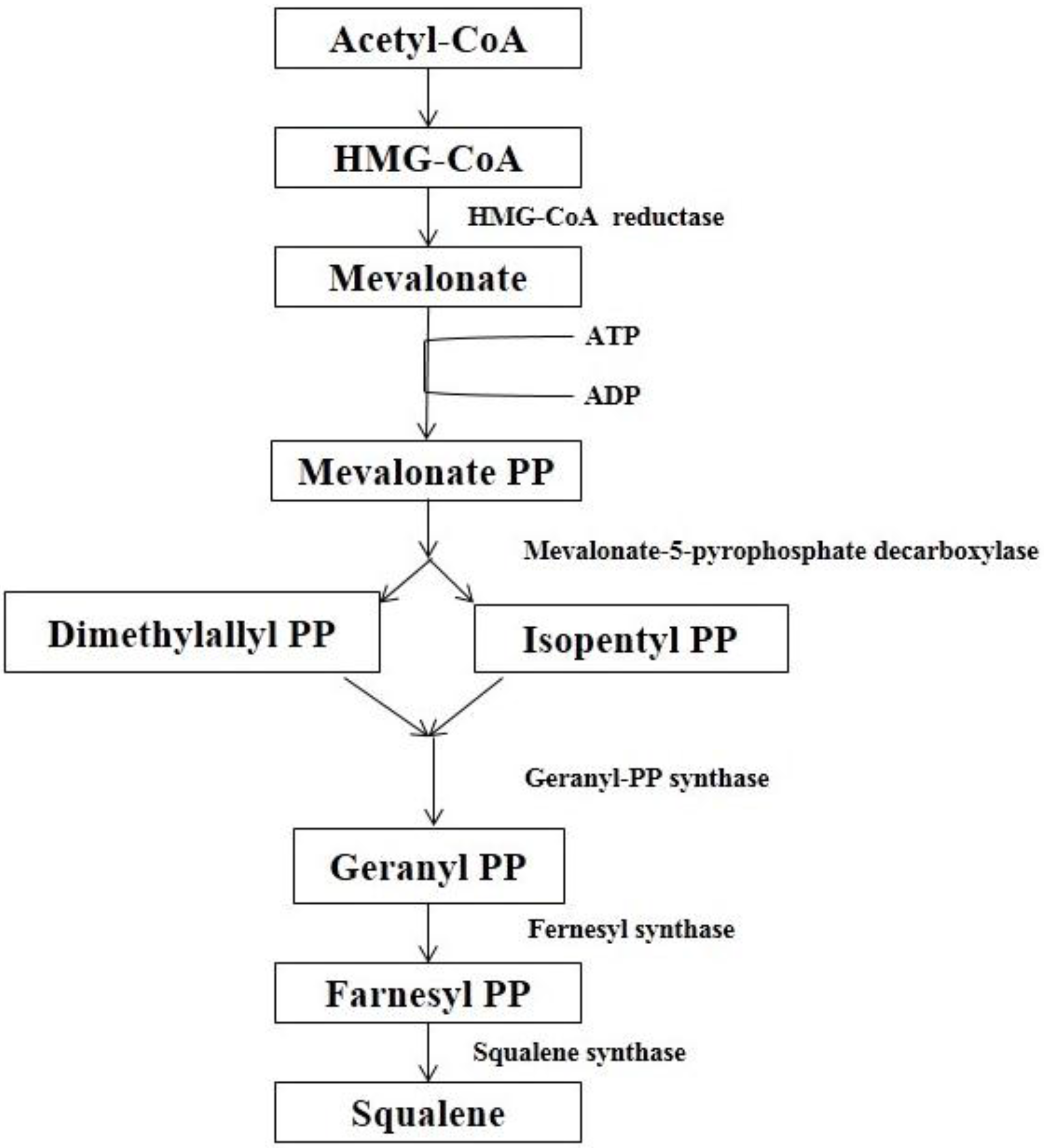
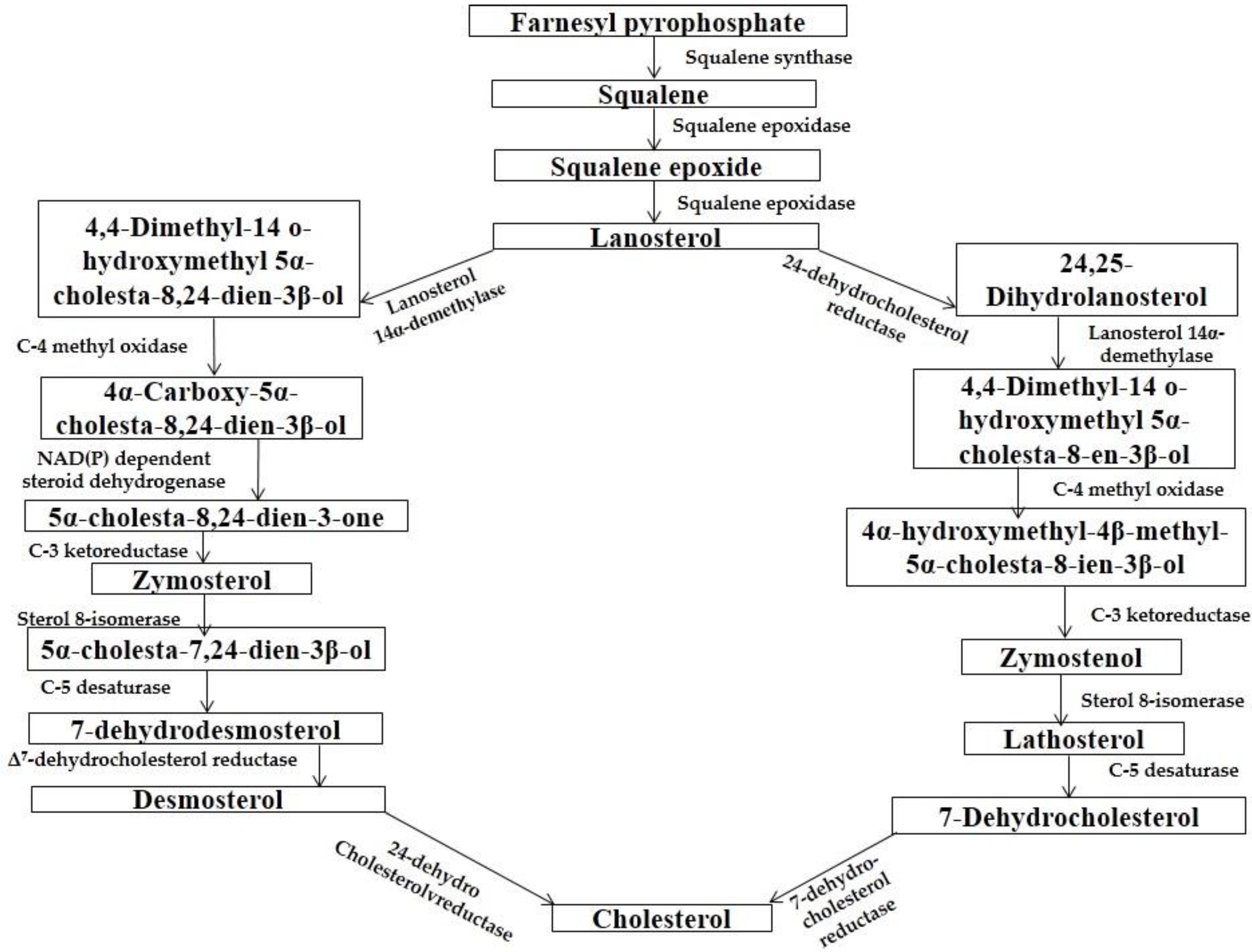
4.1. Biosynthesis
4.2. Comparison of Biosynthesis of BS and Cholesterol
4.3. Chemical Synthesis
5. Systemic Effect
5.1. Central Nervous System
5.2. Skin
5.3. Cardiovascular System
5.4. Liver
5.5. Endocrine System
5.6. Reproductive System
5.7. Wound Healing
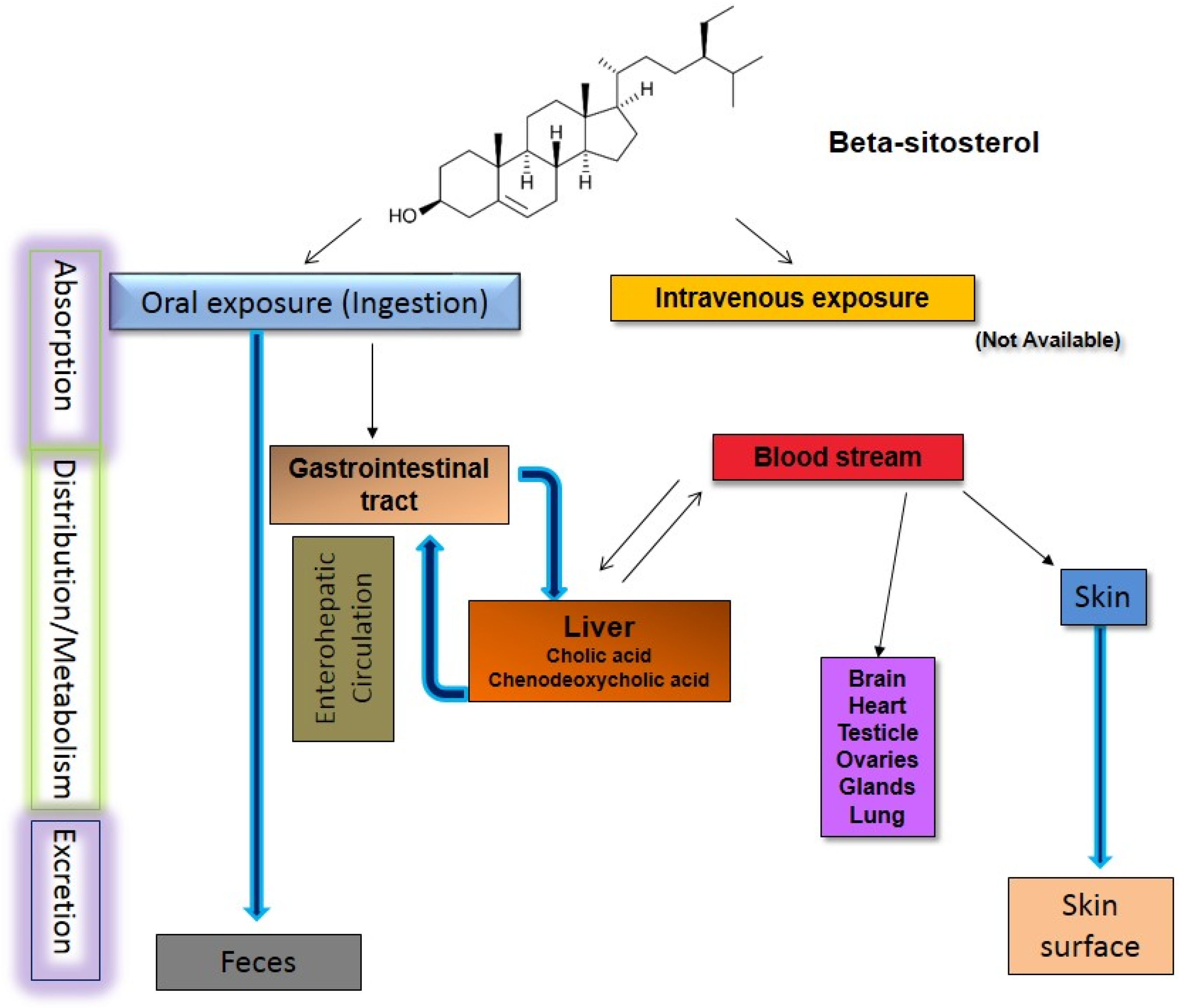
6. Pharmacokinetic Studies
7. Therapeutic Potentials
7.1. Antioxidant Activity
7.2. Angiogenic Effect
7.3. Antihyperlipidemic and Anti-Atherosclerosis Effects
7.4. Antipyretic Activity
7.5. Anti-Inflammatory Activity
7.6. Anti-Arthritic Activity
7.7. Immune Modulation and Anti-HIV Effect
7.8. Anti-Cancer Effect
7.9. Anti-Diabetic Effect
7.10. Anti-Pulmonary Tuberculosis Effect
7.11. Antimicrobial Activity
7.12. Miscellaneous
8. Toxicity
9. Drug Delivery with Beta-Sitosterol
10. Future Research Directions
Acknowledgments
Conflicts of Interest
References
- Bouic, P.; Clark, A.; Lamprecht, J.; Freestone, M.; Pool, E.; Liebenberg, R.; Kotze, D.; van Jaarsveld, P. The effects of B-sitosterol (BSS) and B-sitosterol glucoside (BSSG) mixture on selected immune parameters of marathon runners: Inhibition of post marathon immune suppression and inflammation. Int. J. Sports Med. 1999, 20, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Pegel, K.H. The importance of sitosterol and sitosterolin in human and animal nutrition. S. Afr. J. Sci. 1997, 93, 263–268. [Google Scholar]
- Schneider, K.; DiNovi, M.; Baines, J.; Schlatter, J. Phytosterols, phytostanols and their esters. Saf. Eval. Certain Food Addit. 2009, 60, 117–157. [Google Scholar]
- Raport, E. Consumption of food and beverages with added plant sterols in the European Union. EFSA J. 2008, 133, 1–21. [Google Scholar]
- Paniagua-Pérez, R.; Madrigal-Bujaidar, E.; Reyes-Cadena, S.; Molina-Jasso, D.; Gallaga, J.P.; Silva-Miranda, A.; Velazco, O.; Hernández, N.; Chamorro, G. Genotoxic and cytotoxic studies of beta-sitosterol and pteropodine in mouse. BioMed Res. Int. 2005, 2005, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Wilt, T.J.; Ishani, A.; Rutks, I.; MacDonald, R. Phytotherapy for benign prostatic hyperplasia. Public Health Nutr. 2000, 3, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Shefer, S.; Salen, G.; Nguyen, L.; Batta, A.; Packin, V.; Tint, G.; Hauser, S. Competitive inhibition of bile acid synthesis by endogenous cholestanol and sitosterol in sitosterolemia with xanthomatosis. Effect on cholesterol 7 alpha-hydroxylase. J. Clin. Investig. 1988, 82, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.; Chinnam, M.; Fink, C.; Bradford, P. Β-sitosterol activates fas signaling in human breast cancer cells. Phytomedicine 2007, 14, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Alemany, L.; Barbera, R.; Alegría, A.; Laparra, J. Plant sterols from foods in inflammation and risk of cardiovascular disease: A real threat? Food Chem. Toxicol. 2014, 69, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Vanmierlo, T.; Bogie, J.F.; Mailleux, J.; Vanmol, J.; Lütjohann, D.; Mulder, M.; Hendriks, J.J. Plant sterols: Friend or foe in CNS disorders? Prog. Lipid Res. 2015, 58, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, L.; Guardamagna, O.; Bertucci, P.; Pisciotta, L.; Liberatoscioli, L.; Bertolini, S.; Irace, C.; Gnasso, A.; Federici, G.; Cortese, C. Beta-sitosterolaemia: A new nonsense mutation in the ABCG5 gene. Eur. J. Clin. Investig. 2007, 37, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Mooradian, A.D.; Haas, M.J.; Wong, N.C. The effect of select nutrients on serum high-density lipoprotein cholesterol and apolipoprotein A-I levels. Endocr. Rev. 2006, 27, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Ulbricht, C.E. An evidence-based systematic review of beta-sitosterol, sitosterol (22,23-dihydrostigmasterol, 24-ethylcholesterol) by the natural standard research collaboration. J. Diet. Suppl. 2015, 13, 35–92. [Google Scholar] [PubMed]
- Khan, A.; Rahman, M.; Islam, S. Antipyretic activity of Peperomia pellucida leaves in rabbit. Turk. J. Biol. 2008, 32, 37–41. [Google Scholar]
- Pateh, U.; Haruna, A.; Garba, M.; Iliya, I.; Sule, I.; Abubakar, M.; Ambi, A. Isolation of stigmasterol, β-sitosterol and 2-hydroxyhexadecanoic acid methyl ester from the rhizomes of Stylochiton lancifolius Pyer and Kotchy (Araceae). Niger. J. Pharm. Sci. 2009, 7, 19–25. [Google Scholar]
- Khanam, S.; Sultana, R. Isolation of β-sitosterol & stigmasterol as active immunomodulatory constituents from fruits of Solanum xanthocarpum (Solanaceae). IJPSR 2012, 3, 1057–1060. [Google Scholar]
- Sarin, R.; Bansal, N. Phytosterol from in vivo and in vitro cultures of two medicinal plants viz. Adhatoda Vasica 2011, 2, 927–930. [Google Scholar]
- Ryan, E.; Galvin, K.; O’Connor, T.; Maguire, A.; O’Brien, N. Fatty acid profile, tocopherol, squalene and phytosterol content of brazil, pecan, pine, pistachio and cashew nuts. Int. J. Food Sci. Nutr. 2006, 57, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Kongduang, D.; Wungsintaweekul, J.; de-Eknamkul, W. Biosynthesis of β-sitosterol and stigmasterol proceeds exclusively via the mevalonate pathway in cell suspension cultures of Croton stellatopilosus. Tetrahedron Lett. 2008, 49, 4067–4072. [Google Scholar] [CrossRef]
- De-Eknamkul, W.; Potduang, B. Biosynthesis of β-sitosterol and stigmasterol in Croton sublyratus proceeds via a mixed origin of isoprene units. Phytochemistry 2003, 62, 389–398. [Google Scholar] [CrossRef]
- Klein, R. Phytoecdysteroids. J. Am. Herbalists Guild 2004, 5, 18–28. [Google Scholar]
- Xu, G.; Guan, L.; Sun, J.; Chen, Z.-Y. Oxidation of cholesterol and β-sitosterol and prevention by natural antioxidants. J. Agric. Food Chem. 2009, 57, 9284–9292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Julien-David, D.; Miesch, M.; Geoffroy, P.; Raul, F.; Roussi, S.; Aoudé-Werner, D.; Marchioni, E. Identification and quantitative analysis of β-sitosterol oxides in vegetable oils by capillary gas chromatography–mass spectrometry. Steroids 2005, 70, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Geoffroy, P.; Miesch, M.; Julien-David, D.; Raul, F.; Aoudé-Werner, D.; Marchioni, E. Gram-scale chromatographic purification of β-sitosterol: Synthesis and characterization of β-sitosterol oxides. Steroids 2005, 70, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, L.I.; Rantanen, J.T.; von Bonsdorff, A.K.; Karjalainen, M.A.; Yliruusi, J.K. A novel method of producing a microcrystalline β-sitosterol suspension in oil. Eur. J. Pharm. Sci. 2002, 15, 261–269. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Weingärtner, O.; Pol, S.; Husche, C.; Kerksiek, A.; Friedrichs, S.; Sijbrands, E.; Steinbusch, H.; Grimm, M.; Hartmann, T.; et al. Dietary intake of plant sterols stably increases plant sterol levels in the murine brain. J. Lipid Res. 2012, 53, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, W.; Sheppard, A.; Newkirk, D.; Prosser, A.; Osgood, T. Comparison of various methods for the extraction of total lipids, fatty acids, cholesterol, and other sterols from food products. J. Am. Oil Chem. Soc. 1977, 54, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Careri, M.; Elviri, L.; Mangia, A. Liquid chromatography–UV determination and liquid chromatography–atmospheric pressure chemical ionization mass spectrometric characterization of sitosterol and stigmasterol in soybean oil. J. Chromatogr. A 2001, 935, 249–257. [Google Scholar] [CrossRef]
- Nagy, K.; Jakab, A.; Pollreisz, F.; Bongiorno, D.; Ceraulo, L.; Averna, M.R.; Noto, D.; Vekey, K. Analysis of sterols by high-performance liquid chromatography/mass spectrometry combined with chemometrics. Rapid Commun. Mass Spectrom. 2006, 20, 2433–2440. [Google Scholar] [CrossRef] [PubMed]
- Ahmida, H.M.; Bertucci, P.; Franzò, L.; Massoud, R.; Cortese, C.; Lala, A.; Federici, G. Simultaneous determination of plasmatic phytosterols and cholesterol precursors using gas chromatography–mass spectrometry (GC–MS) with selective ion monitoring (SIM). J. Chromatogr. B 2006, 842, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Nair, V.; Kanfer, I.; Hoogmartens, J. Determination of stigmasterol, β-sitosterol and stigmastanol in oral dosage forms using high performance liquid chromatography with evaporative light scattering detection. J. Pharm. Biomed. Anal. 2006, 41, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.; Jha, S.; Murthy, P.N.; Manik; Sharone, A. Isolation and characterization of stigmast-5-en-3β-ol (β-sitosterol) from the leaves of Hygrophila spinosa. Int. J. Pharm. Sci. Res. 2010, 2, 95–100. [Google Scholar]
- Karan, S.K.; Mishra, S.K.; Pal, D.; Mondal, A. Isolation of β-sitosterol and evaluation of antidiabetic activity of Aristolochia indica in alloxan-induced diabetic mice with a reference to in vitro antioxidant activity. J. Med. Plant Res. 2012, 6, 1219–1223. [Google Scholar]
- Ahmed, K.B.A.; Subramaniam, S.; Veerappan, G.; Hari, N.; Sivasubramanian, A.; Veerappana, A. β-sitosterol-d-glucopyranoside isolated from Desmostachya bipinnata mediate photoinduced rapid green synthesis of silver nanoparticles. RSC Adv. 2014, 4, 59130–59136. [Google Scholar] [CrossRef]
- Chaturvedula, V.S.P.; Prakash, I. Isolation of Stigmasterol and β-Sitosterol from the dichloromethane extract of Rubus suavissimus. Int. Curr. Pharm. J. 2012, 1, 239–242. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, H.; Lu, Z.; Xu, Z. Quantitative determination of steroids in the fruiting bodies and submerged-cultured mycelia of Inonotus obliquus. Chin. J. Chromatogr. 2009, 27, 745–749. [Google Scholar]
- Srividya, N.; Heidorn, D.B.; Lange, B.M. Rapid purification of gram quantities of β-sitosterol from a commercial phytosterol mixture. BMC Res. Notes 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Mackay, D.S.; Jones, P.J.H.; Myrie, S.B.; Plat, J.; Lütjohann, D. Methodological considerations for the harmonization of non-cholesterol sterol bio-analysis. J. Chromatogr. B 2014, 957, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2009; p. 539. [Google Scholar]
- Joffrion, T.M.; Cushion, M.T. Sterol biosynthesis and sterol uptake in the fungal pathogen Pneumocystis carinii. FEMS Microbiol. Lett. 2010, 311, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Checa, J.C. Free Cholesterol—A Double-Edge Sword in Alzheimer Disease—Challenges for the Future; Inga, Z., Ed.; Georg August University: Göttingen, Germany, 2015; pp. 141–166. [Google Scholar]
- Geoffroy, P.; Julien-David, D.; Marchioni, E.; Raul, F.; Aoudé-Werner, D.; Miesch, M. Synthesis of highly pure oxyphytosterols and (oxy)phytosterol esters: Part I. Regioselective hydrogenation of stigmasterol: An easy access to oxyphytosterols. Steroids 2008, 73, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Foley, D.A.; O’Callaghan, Y.; O’Brien, N.M.; McCarthy, F.O.; Maguire, A.R. Synthesis and characterization of stigmasterol oxidation products. J. Agric. Food Chem. 2009, 58, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Khripach, V.A.; Zhabinskii, V.N.; Konstantinova, O.V.; Khripach, N.B.; Antonchick, A.V.; Antonchick, A.P.; Schneider, B. Preparation of (25R)-and (25S)-26-functionalized steroids as tools for biosynthetic studies of cholic acids. Steroids 2005, 70, 551–562. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, F.O.; Chopra, J.; Ford, A.; Hogan, S.A.; Kerry, J.P.; O’Brien, N.M.; Ryan, E.; Maguire, A.R. Synthesis, isolation and characterisation of β-sitosterol and β-sitosterol oxide derivatives. Org. Biomol. Chem. 2005, 3, 3059–3065. [Google Scholar] [CrossRef] [PubMed]
- Hang, J.; Dussault, P. A concise synthesis of β-sitosterol and other phytosterols. Steroids 2010, 75, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Niero, R.; Yunes, R.; Pizzolatti, M.; Delle Monache, F.; Calixto, J. Antinociceptive properties of steroids isolated from Phyllanthus corcovadensis in mice. Planta Medica 1995, 61, 329–332. [Google Scholar] [CrossRef] [PubMed]
- López-Rubalcava, C.; Piña-Medina, B.; Estrada-Reyes, R.; Heinze, G.; Martínez-Vázquez, M. Anxiolytic-like actions of the hexane extract from leaves of Annona cherimolia in two anxiety paradigms: Possible involvement of the GABA/benzodiazepine receptor complex. Life Sci. 2006, 78, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Hernández, E.; Rosas-Acevedo, H.; Soto-Hernández, M.; Martínez, A.L.; Moreno, J.; González-Trujano, M.E. Bioactivity-guided isolation of beta-sitosterol and some fatty acids as active compounds in the anxiolytic and sedative effects of Tilia americana var. Mexicana. Planta Medica 2007, 73, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M. CNS active principles from Alangium plantanifolium. Planta Medica 1998, 64, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Brimson, J.M.; Brimson, S.J.; Brimson, C.A.; Rakkhitawatthana, V.; Tencomnao, T. Rhinacanthus nasutus extracts prevent glutamate and amyloid-β neurotoxicity in ht-22 mouse hippocampal cells: Possible active compounds include lupeol, stigmasterol and β-sitosterol. Int. J. Mol. Sci. 2012, 13, 5074–5097. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liu, J.; Wu, F.; Zhu, X.; Yew, D.T.; Xu, J. Β-sitosterol inhibits high cholesterol-induced platelet β-amyloid release. J. Bioenerg. Biomembr. 2011, 43, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wu, F.; Xu, J. Incorporation of β-sitosterol into mitochondrial membrane enhances mitochondrial function by promoting inner mitochondrial membrane fluidity. J. Bioenerg. Biomembr. 2013, 45, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Hamedi, A.; Ghanbari, A.; Saeidi, V.; Razavipour, R.; Azari, H. Effects of β-sitosterol oral administration on the proliferation and differentiation of neural stem cells. J. Funct. Foods 2014, 8, 252–258. [Google Scholar] [CrossRef]
- Hamedi, A.; Ghanbari, A.; Razavipour, R.; Saeidi, V.; Zarshenas, M.M.; Sohrabpour, M.; Azari, H. Alyssum homolocarpum seeds: Phytochemical analysis and effects of the seed oil on neural stem cell proliferation and differentiation. J. Nat. Med. 2015, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Han, N.-R.; Kim, H.-M.; Jeong, H.-J. The β-sitosterol attenuates atopic dermatitis-like skin lesions through down-regulation of TSLP. Exp. Biol. Med. 2014, 239, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, Z.; Yang, H.; Cao, L.; Liu, F.; Bai, X.; Ruan, C. Clinical and molecular genetic analysis of a family with sitosterolemia and co-existing erythrocyte and platelet abnormalities. Haematologica 2006, 91, 1392–1395. [Google Scholar] [PubMed]
- Brufaua, G.; Canelab, M.A.; Rafecas, M. Phytosterols: Physiologic and metabolic aspects related to cholesterol-lowering properties. Nutr. Res. 2008, 28, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Suzuki, H.; Itoh, K. Effect of beta-sitosterol on ultrastructure of liver cells in young and aged mice. Int. J. Vitam. Nutr. Res. 1997, 68, 146–148. [Google Scholar]
- Goswami, S.K.; Frey, C.F. Effect of beta-sitosterol on cholesterol-cholic acid-induced gallstone formation in mice. Am. J. Gastroenterol. 1976, 65, 305–310. [Google Scholar] [PubMed]
- Singhal, A.K.; Cohen, B.I.; Finver-Sadowsky, J.; McSherry, C.K.; Mosbach, E.H. Role of hydrophilic bile acids and of sterols on cholelithiasis in the hamster. J. Lipid Res. 1984, 25, 564–570. [Google Scholar] [PubMed]
- Bustos, P.; Duffau, C.; Pacheco, C.; Ulloa, N. Β-sitosterol modulation of monocyte-endothelial cell interaction: A comparison to female hormones. Maturitas 2008, 60, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-X.; Lin, Y.-X.; Ou, M.-R.; Tan, D.-F. An experimental study (II) on the inhibition of prostatic hyperplasia by extract of seeds of Brassica alba. Zhongguo Zhong Yao Za Zhi 2003, 28, 643–646. [Google Scholar] [PubMed]
- Leusch, F.D.; MacLatchy, D.L. In vivo implants of β-sitosterol cause reductions of reactive cholesterol pools in mitochondria isolated from gonads of male goldfish (Carassius auratus). Gen. Comp. Endocrinol. 2003, 134, 255–263. [Google Scholar] [CrossRef]
- Promprom, W.; Kupittayanant, P.; Indrapichate, K.; Wray, S.; Kupittayanant, S. The effects of pomegranate seed extract and β-sitosterol on rat uterine contractions. Reprod. Sci. 2010, 17, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, P.; Pölönen, I.; Mustonen, A.-M. Increased reproductive success in the white American mink (Neovison vison) with chronic dietary β-sitosterol supplement. Anim. Reprod. Sci. 2010, 119, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Malini, T.; Vanithakumari, G. Rat toxicity studies with β-sitosterol. J. Ethnopharmacol. 1990, 28, 221–234. [Google Scholar] [CrossRef]
- Chattopadhyay, D.; Dungdung, S.R.; Mandal, A.B.; Majumder, G.C. A potent sperm motility-inhibiting activity of bioflavonoids from an ethnomedicine of onge, Alstonia macrophylla Wall ex A. Dc, leaf extract. Contraception 2005, 71, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Ryökkynen, A.; Käyhkö, U.-R.; Mustonen, A.-M.; Kukkonen, J.V.; Nieminen, P. Multigenerational exposure to phytosterols in the mouse. Reprod. Toxicol. 2005, 19, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Martel-Estrada, S.A.; Rodríguez-Espinoza, B.; Santos-Rodríguez, E.; Jiménez-Vega, F.; García-Casillas, P.E.; Martínez-Pérez, C.A.; Armendáriz, I.O. Biocompatibility of chitosan/Mimosa tenuiflora scaffolds for tissue engineering. J. Alloy. Compd. 2015, 643, 119–123. [Google Scholar] [CrossRef]
- Ikeda, I.; Sugano, M. Comparison of absorption and metabolism of β-sitosterol and β-sitostanol in rats. Atherosclerosis 1978, 30, 227–237. [Google Scholar] [CrossRef]
- Ritschel, W.; Kastner, U.; Hussain, A.; Koch, H. Pharmacokinetics and bioavailability of beta-sitosterol in the beagle dog. Arzneim.-Forsch. 1990, 40, 463–468. [Google Scholar]
- Relas, H.; Gylling, H.; Miettinen, T.A. Fate of intravenously administered squalene and plant sterols in human subjects. J. Lipid Res. 2001, 42, 988–994. [Google Scholar] [PubMed]
- Duchateau, G.; Cochrane, B.; Windebank, S.; Rudzinska, J.; Sanghera, D.; Burien, A.M.; Müller, M.; Zeitlinger, M.; Lappin, G. Absolute oral bioavailability and metabolic turnover of β-sitosterol in healthy subjects. Drug Metab. Dispos. 2012, 40, 2026–2030. [Google Scholar] [CrossRef] [PubMed]
- Sudhop, T.; Sahin, Y.; Lindenthal, B.; Hahn, C.; Lüers, C.; Berthold, H.; von Bergmann, K. Comparison of the hepatic clearances of campesterol, sitosterol, and cholesterol in healthy subjects suggests that efflux transporters controlling intestinal sterol absorption also regulate biliary secretion. Gut 2002, 51, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Gilman, C.I.; Leusch, F.D.; Breckenridge, W.C.; MacLatchy, D.L. Effects of a phytosterol mixture on male fish plasma lipoprotein fractions and testis P450scc activity. Gen. Comp. Endocrinol. 2003, 130, 172–184. [Google Scholar] [CrossRef]
- Ikeda, I.; Tanaka, K.; Sugano, M.; Vahouny, G.; Gallo, L. Discrimination between cholesterol and sitosterol for absorption in rats. J. Lipid Res. 1988, 29, 1583–1591. [Google Scholar] [PubMed]
- Salen, G.; Shore, V.; Tint, G.; Forte, T.; Shefer, S.; Horak, I.; Horak, E.; Dayal, B.; Nguyen, L.; Batta, A. Increased sitosterol absorption, decreased removal, and expanded body pools compensate for reduced cholesterol synthesis in sitosterolemia with xanthomatosis. J. Lipid Res. 1989, 30, 1319–1330. [Google Scholar] [PubMed]
- Salen, G.; Ahrens, E., Jr.; Grundy, S.M. Metabolism of β-sitosterol in man. J. Clin. Investig. 1970, 49, 952. [Google Scholar] [CrossRef] [PubMed]
- Sirtori, C.; Manzoni, C.; Lovati, M. Mechanisms of lipid-lowering agents. Cardiology 1991, 78, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Hauser, H.; Dyer, J.H.; Nandy, A.; Vega, M.A.; Werder, M.; Bieliauskaite, E.; Weber, F.E.; Compassi, S.; Gemperli, A.; Boffelli, D. Identification of a receptor mediating absorption of dietary cholesterol in the intestine. Biochemistry 1998, 37, 17843–17850. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Mensink, R.P. Plant stanol and sterol esters in the control of blood cholesterol levels: Mechanism and safety aspects. Am. J. Cardiol. 2005, 96, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Nichols, J.A.; Mensink, R.P. Plant sterols and stanols: Effects on mixed micellar composition and LXR (target gene) activation. J. Lipid Res. 2005, 46, 2468–2476. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, W.; Lei, L.; Liu, Y.; Ma, K.Y.; Li, Y.M.; Wang, L.; Huang, Y.; Chen, Z.-Y. Blockage of hydroxyl group partially abolishes the cholesterol-lowering activity of β-sitosterol. J. Funct. Foods 2015, 12, 199–207. [Google Scholar] [CrossRef]
- Calpe-Berdiel, L.; Escolà-Gil, J.C.; Blanco-Vaca, F. New insights into the molecular actions of plant sterols and stanols in cholesterol metabolism. Atherosclerosis 2009, 203, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.; Sugano, M. Some aspects of mechanism of inhibition of cholesterol absorption by β-sitosterol. Biochim. Biophys. Acta (BBA)-Biomembr. 1983, 732, 651–658. [Google Scholar] [CrossRef]
- Field, F.J.; Mathur, S.N. Beta-sitosterol: Esterification by intestinal acylcoenzyme A: Cholesterol acyltransferase (ACAT) and its effect on cholesterol esterification. J. Lipid Res. 1983, 24, 409–417. [Google Scholar] [PubMed]
- Sanders, D.; Minter, H.; Howes, D.; Hepburn, P. The safety evaluation of phytosterol esters. Part 6. The comparative absorption and tissue distribution of phytosterols in the rat. Food Chem. Toxicol. 2000, 38, 485–491. [Google Scholar] [CrossRef]
- Sehayek, E.; Duncan, E.M.; Lutjohann, D.; von Bergmann, K.; Ono, J.G.; Batta, A.K.; Salen, G.; Breslow, J.L. Loci on chromosomes 14 and 2, distinct from ABCG5/ABCG8, regulate plasma plant sterol levels in a C57bl/6J× CASA/Rk intercross. Proc. Natl. Acad. Sci. USA 2002, 99, 16215–16219. [Google Scholar] [CrossRef] [PubMed]
- Jansen, P.J.; Lütjohann, D.; Abildayeva, K.; Vanmierlo, T.; Plösch, T.; Plat, J.; von Bergmann, K.; Groen, A.K.; Ramaekers, F.C.; Kuipers, F. Dietary plant sterols accumulate in the brain. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2006, 1761, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Jin, C.; Park, E.-H. Identification of metabolites of phytosterols in rat feces using GC/MS. Arch. Pharm. Res. 2000, 23, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Aringer, L.; Eneroth, P. Formation and metabolism in vitro of 5,6-epoxides of cholesterol and β-sitosterol. J. Lipid Res. 1974, 15, 389–398. [Google Scholar] [PubMed]
- Aringer, L.; Eneroth, P.; Nordström, L. Side chain hydroxylation of cholesterol, campesterol and beta-sitosterol in rat liver mitochondria. J. Lipid Res. 1976, 17, 263–272. [Google Scholar] [PubMed]
- Miettinen, T.A.; Vanhanen, H. Dietary-sitostanol related to absorption, synthesis and serum level of cholesterol in different apolipoprotein E phenotypes. Atherosclerosis 1994, 105, 217–226. [Google Scholar] [CrossRef]
- Grundy, S.M.; Ahrens, E.H., Jr.; Salen, G. Dietary β-sitosterol as an internal standard to correct for cholesterol losses in sterol balance studies. J. Lipid Res. 1968, 9, 374–387. [Google Scholar] [PubMed]
- Bhattacharyya, A.K.; Connor, W.E.; Lin, D.S. The origin of plant sterols in the skin surface lipids in humans: From diet to plasma to skin. J. Investig. Dermatol. 1983, 80, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, T.; Tarpila, S. Fecal β-sitosterol in patients with diverticular disease of the colon and in vegetarians. Scand. J. Gastroenterol. 1978, 13, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, W.; Nye, E.; Macfarlane, D.; Williamson, S.; Robertson, M. Cholesterol metabolism in distance runners. Clin. Physiol. 1992, 12, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Vuoristo, M.; Tilvis, R.; Miettinen, T. Serum plant sterols and lathosterol related to cholesterol absorption in coeliac disease. Clin. Chim. Acta 1988, 174, 213–224. [Google Scholar] [CrossRef]
- Gupta, R.; Sharma, A.K.; Dobhal, M.; Sharma, M.; Gupta, R. Antidiabetic and antioxidant potential of β-sitosterol in streptozotocin-induced experimental hyperglycemia. J. Diabetes 2011, 3, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.J.; Mitjavila, M.T. The degree of unsaturation of dietary fatty acids and the development of atherosclerosis (review). J. Nutr. Biochem. 2003, 14, 182–195. [Google Scholar] [CrossRef]
- Vivancos, M.; Moreno, J.J. Β-sitosterol modulates antioxidant enzyme response in raw 264.7 macrophages. Free Radic. Biol. Med. 2005, 39, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Moon, E.-J.; Lee, Y.M.; Lee, O.-H.; Lee, M.-J.; Lee, S.-K.; Chung, M.-H.; Park, Y.-I.; Sung, C.-K.; Choi, J.-S.; Kim, K.-W. A ncovel angiogenic factor derived from aloe vera gel: Β-sitosterol, a plant sterol. Angiogenesis 1999, 3, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Kong, K.R.; Kim, Y.; Jung, K.-O.; Kil, J.-H.; Rhee, S.-H.; Park, K.-Y. Induction of bax and activation of caspases during β-sitosterol-mediated apoptosis in human colon cancer cells. Int. J. Oncol. 2003, 23, 1657–1662. [Google Scholar] [CrossRef] [PubMed]
- Pollak, O.J.; Kritchevsky, D. Sitosterol; Karger: Basel, Switzerland, 1981. [Google Scholar]
- Hu, F.B. Plant-based foods and prevention of cardiovascular disease: An overview. Am. J. Clin. Nutr. 2003, 78, 544S–551S. [Google Scholar] [PubMed]
- Retelny, V.S.; Neuendorf, A.; Roth, J.L. Nutrition protocols for the prevention of cardiovascular disease. Nutr. Clin. Pract. 2008, 23, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Yeshurun, D.; Gotto, A. Drug treatment of hyperlipidemia. Am. J. Med. 1976, 60, 379–396. [Google Scholar] [CrossRef]
- Jesch, E.D.; Carr, T.P. Sitosterol reduces micellar cholesterol solubility in model bile. Nutr. Res. 2006, 26, 579–584. [Google Scholar] [CrossRef]
- Sugano, M.; Morioka, H.; Ikeda, I. A comparison of hypocholesterolemic activity of beta-sitosterol and beta-sitostanol in rats. J. Nutr. 1977, 107, 2011–2019. [Google Scholar] [PubMed]
- Zak, A.; Zeman, M.; Vitkova, D.; Hrabak, P.; Tvrzicka, E. Beta-sitosterol in the treatment of hypercholesterolemia. Casopis Lekaru Ceskych 1990, 129, 1320–1323. [Google Scholar] [PubMed]
- Martins, S.; Silva, H.F.; Novaes, M.; Ito, M.K. Therapeutic effects of phytosterols and phytostanols in cholesterolemia. Arch. Latinoam. Nutr. 2004, 54, 257–263. [Google Scholar] [PubMed]
- Ellegard, L.H.; Andersson, S.W.; Normén, A.L.; Andersson, H.A. Dietary plant sterols and cholesterol metabolism. Nutr. Rev. 2007, 65, 39. [Google Scholar] [CrossRef] [PubMed]
- Richter, W.O.; Geiss, H.C.; Sönnichsen, A.C.; Schwandt, P. Treatment of severe hypercholesterolemia with a combination of beta-sitosterol and lovastatin. Curr. Ther. Res. 1996, 57, 497–505. [Google Scholar] [CrossRef]
- Weingärtner, O.; Böhm, M.; Laufs, U. Controversial role of plant sterol esters in the management of hypercholesterolaemia. Eur. Heart J. 2009, 30, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Volkova, N.; Aviram, M. Pomegranate phytosterol (β-sitosterol) and polyphenolic antioxidant (punicalagin) addition to statin, significantly protected against macrophage foam cells formation. Atherosclerosis 2013, 226, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Wilund, K.R.; Yu, L.; Xu, F.; Vega, G.L.; Grundy, S.M.; Cohen, J.C.; Hobbs, H.H. No association between plasma levels of plant sterols and atherosclerosis in mice and men. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2326–2332. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Nath, R.; Srivastava, N.; Shanker, K.; Kishor, K.; Bhargava, K. Anti-inflammatory and antipyretic activities of beta-sitosterol (Cyperus). Planta Medica 1980, 39, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Begum, T.N.; Ilyas, M.H.M.; Anand, A.V. Antipyretic activity of Azima tetracantha in experimental animals. Int. J. Curr. Biomed. Pharm. Res. 2011, 1, 41–44. [Google Scholar]
- Loizou, S.; Lekakis, I.; Chrousos, G.P.; Moutsatsou, P. Β-sitosterol exhibits anti-inflammatory activity in human aortic endothelial cells. Mol. Nutr. Food Res. 2010, 54, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, S.A.; Pal, S.C.; Mandal, S.C.; Patil, A.N. Analgesic and anti-inflammatory activity of β-sitosterol isolated from Nyctanthes arbortristis leaves. Inflammopharmacology 2012, 20, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Garcia, J.; Recio, M.C.; Giner, R.M. Anti-inflammatory activity of beta-sitosterol in a model of oxazolone-induced contact delayed type hypersensitivity. Bol. Latinoam. Caribe Plantas Med. Aromát. 2006, 6, 57–62. [Google Scholar]
- Bouic, P.; Etsebeth, S.; Liebenberg, R.; Albrecht, C.; Pegel, K.; Van Jaarsveld, P. Beta-sitosterol and beta-sitosterol glucoside stimulate human peripheral blood lymphocyte proliferation: Implications for their use as an immunomodulatory vitamin combination. Int. J. Immunopharmacol. 1996, 18, 693–700. [Google Scholar] [CrossRef]
- Bouic, P.; Lamprecht, J.H. Plant sterols and sterolins: A review of their immune-modulating properties. Altern. Med. Rev. 1999, 4, 170–177. [Google Scholar] [PubMed]
- Valerio, M.; Awad, A.B. Β-sitosterol down-regulates some pro-inflammatory signal transduction pathways by increasing the activity of tyrosine phosphatase SHP-1 in J774A. 1 murine macrophages. Int. Immunopharmacol. 2011, 11, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.; Saenz, M.; Garcia, M.; Fernandez, M. Study of the topical anti-inflammatory activity of Achillea ageratum on chronic and acute inflammation models. Z. Naturforschung C 1999, 54, 937–941. [Google Scholar] [CrossRef]
- Navarro, A.; De Las Heras, B.; Villar, A. Anti-inflammatory and immunomodulating properties of a sterol fraction from Sideritis foetens Clem. Biol. Pharm. Bull. 2001, 24, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Breu, W.; Hagenlocher, M.; Redl, K.; Stadler, F.; Wagner, H. Anti-inflammatory activity of sabal fruit extracts prepared with supercritical carbon dioxide. In vitro antagonists of cyclooxygenase and 5-lipoxygenase metabolism. Arzneim.-Forsch. 1992, 42, 547–551. [Google Scholar]
- Liz, R.; Zanatta, L.; dos Reis, G.O.; Horst, H.; Pizzolatti, M.G.; Silva, F.R.M.B.; Fröde, T.S. Acute effect of β-sitosterol on calcium uptake mediates anti-inflammatory effect in murine activated neutrophils. J. Pharm. Pharmacol. 2013, 65, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.G.; Mehta, A.A. Suppression of ovalbumin-induced Th2-driven airway inflammation by β-sitosterol in a guinea pig model of asthma. Eur. J. Pharmacol. 2011, 650, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Tatiya, A.U.; Saluja, A.K. Further studies on membrane stabilizing, anti-inflammatory and FCA induced arthritic activity of various fractions of bark of Machilus macrantha in rats. Rev. Bras. Farmacogn. 2011, 21, 1052–1064. [Google Scholar] [CrossRef]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Fraile, L.; Crisci, E.; Córdoba, L.; Navarro, M.A.; Osada, J.; Montoya, M. Immunomodulatory properties of beta-sitosterol in pig immune responses. Int. Immunopharmacol. 2012, 13, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Bouic, P.J. The role of phytosterols and phytosterolins in immune modulation: A review of the past 10 years. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Breytenbach, U.; Clark, A.; Lamprecht, J.; Bouic, P. Flow cytometric analysis of the Th1–Th2 balance in healthy individuals and patients infected with the human immunodeficiency virus (HIV) receiving a plant sterol/sterolin mixture. Cell Biol. Int. 2001, 25, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Bouic, P.J. Immunomodulation in HIV/AIDS: The Tygerberg/Stellenbosch University Experience. AIDS Bull. 1997, 6, 18–20. [Google Scholar]
- Neurath, A.R.; Strick, N.; Li, Y.Y.; Debnath, A.K. Punica granatum (Pomegranate) juice provides an HIV-1 entry inhibitor and candidate topical microbicide. Ann. N. Y. Acad. Sci. 2005, 1056, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Kotwal, G.J. Genetic diversity-independent neutralization of pandemic viruses (eg HIV), potentially pandemic (eg H5N1 strain of influenza) and carcinogenic (eg HBV and HCV) viruses and possible agents of bioterrorism (variola) by enveloped virus neutralizing compounds (EVNCS). Vaccine 2008, 26, 3055–3058. [Google Scholar] [PubMed]
- Kotwal, G.J.; Kulkarni, A.P. Antigenic variation in microbial evasion of immune responses. eLS 2013. [Google Scholar] [CrossRef]
- Bin Sayeed, M.S.; Ameen, S.S. Beta-sitosterol: A promising but orphan nutraceutical to fight against cancer. Nutr. Cancer 2015, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.; Downie, A.C.; Fink, C.S. Inhibition of growth and stimulation of apoptosis by beta-sitosterol treatment of MDA-MB-231 human breast cancer cells in culture. Int. J. Mol. Med. 2000, 5, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Moon, D.-O.; Rhu, C.-H.; Choi, B.T.; Lee, W.H.; Kim, G.-Y.; Choi, Y.H. Beta-sitosterol induces anti-proliferation and apoptosis in human leukemic U937 cells through activation of caspase-3 and induction of Bax/Bcl-2 ratio. Biol. Pharm. Bull. 2007, 30, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Kim, M.-O.; Choi, Y.H.; Kim, G.-Y. Β-sitosterol induces G 2/M arrest, endoreduplication, and apoptosis through the Bcl-2 and PI3K/Akt signaling pathways. Cancer Lett. 2008, 264, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Tasyriq, M.; Najmuldeen, I.A.; In, L.L.; Mohamad, K.; Awang, K.; Hasima, N. 7α-hydroxy-β-sitosterol from Chisocheton tomentosus induces apoptosis via dysregulation of cellular Bax/Bcl-2 ratio and cell cycle arrest by downregulating Erk1/2 activation. eCAM 2012, 12, 155–237. [Google Scholar]
- Awad, A.; Chen, Y.; Fink, C.; Hennessey, T. Beta-sitosterol inhibits ht-29 human colon cancer cell growth and alters membrane lipids. Anticancer Res. 1995, 16, 2797–2804. [Google Scholar]
- Awad, A.; Von Holtz, R.; Cone, J.; Fink, C.; Chen, Y. Beta-sitosterol inhibits growth of HT-29 human colon cancer cells by activating the sphingomyelin cycle. Anticancer Res. 1997, 18, 471–473. [Google Scholar]
- Baskar, A.A.; Ignacimuthu, S.; Paulraj, G.M.; Al Numair, K.S. Chemopreventive potential of β-sitosterol in experimental colon cancer model-an in vitro and in vivo study. BMC Complement. Altern. Med. 2010, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Normén, A.L.; Brants, H.A.; Voorrips, L.E.; Andersson, H.A.; van den Brandt, P.A.; Goldbohm, R.A. Plant sterol intakes and colorectal cancer risk in the netherlands cohort study on diet and cancer. Am. J. Clin. Nutr. 2001, 74, 141–148. [Google Scholar] [PubMed]
- Awad, A.B.; Burr, A.T.; Fink, C.S. Effect of resveratrol and β-sitosterol in combination on reactive oxygen species and prostaglandin release by PC-3 cells. Prostaglandins Leukot. Essent. Fatty Acids 2005, 72, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Jourdain, C.; Tenca, G.; Deguercy, A.; Troplin, P.; Poelman, D. In Vitro effects of polyphenols from cocoa and β-sitosterol on the growth of human prostate cancer and normal cells. Eur. J. Cancer Prev. 2006, 15, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasha, G.; Mandadi, K.; Poulose, S.M.; Jadegoud, Y.; Gowda, G.N.; Patil, B.S. Inhibition of colon cancer cell growth and antioxidant activity of bioactive compounds from Poncirus trifoliata (L.) Raf. Bioorg. Med. Chem. 2007, 15, 4923–4932. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, Y.; Chen, S.; Ge, X.; Xu, C.; Gui, S. Effects of beta-sitosterol on microtubular systems in cervical cancer cells. Zhonghua Yi Xue Za Zhi 2006, 86, 2771–2775. [Google Scholar] [PubMed]
- Matos, M.; Leite, L.; Brustolim, D.; De Siqueira, J.; Carollo, C.; Hellmann, A.; Pereira, N.; da Silva, D. Antineoplastic activity of selected constituents of Duguetia glabriuscula. Fitoterapia 2006, 77, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.P.; Turjman, N.; Kessie, G.; Calkins, B.; Goodman, G.T.; Davidovitz, H.; Nimmagadda, G. Diet, nutrition intake, and metabolism in populations at high and low risk for colon cancer. Dietary cholesterol, beta-sitosterol, and stigmasterol. Am. J. Clin. Nutr. 1984, 40, 927–930. [Google Scholar] [PubMed]
- Carilla, E.; Briley, M.; Fauran, F.; Sultan, C.; Duvilliers, C. Binding of permixon, a new treatment for prostatic benign hyperplasia, to the cytosolic androgen receptor in the rat prostate. J. Steroid Biochem. 1984, 20, 521–523. [Google Scholar] [CrossRef]
- Von Holtz, R.L.; Fink, C.S.; Awad, A.B. Β-sitosterol activates the sphingomyelin cycle and induces apoptosis in LNCaP human prostate cancer cells. Nutr. Cancer 1998, 32, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Fink, C.S. Phytosterols as anticancer dietary components: Evidence and mechanism of action. J. Nutr. 2000, 130, 2127–2130. [Google Scholar] [PubMed]
- Awad, A.B.; Williams, H.; Fink, C.S. Phytosterols reduce in vitro metastatic ability of MDA-MB-231 human breast cancer cells. Nutr. Cancer 2001, 40, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Roy, R.; Fink, C.S. Β-sitosterol, a plant sterol, induces apoptosis and activates key caspases in MDA-MB-231 human breast cancer cells. Oncol. Rep. 2003, 10, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Lee, K.-J.; Choi, Y.H.; Kim, G.-Y. Β-sitosterol-induced-apoptosis is mediated by the activation of ERK and the downregulation of Akt in MCA-102 murine fibrosarcoma cells. Int. Immunopharmacol. 2007, 7, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.J. Effect of olive oil minor components on oxidative stress and arachidonic acid mobilization and metabolism by macrophages RAW 264.7. Free Radic. Biol. Med. 2003, 35, 1073–1081. [Google Scholar] [CrossRef]
- Hueber, A. Role of membrane microdomain rafts in TNFR-mediated signal transduction. Cell Death Differ. 2003, 10, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Scheel-Toellner, D.; Wang, K.; Assi, L.; Webb, P.; Craddock, R.; Salmon, M.; Lord, J. Clustering of death receptors in lipid rafts initiates neutrophil spontaneous apoptosis. Biochem. Soc. Trans. 2004, 32, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.; Chinnam, M.; Fink, C.; Bradford, P. Targeting ceramide by dietary means to stimulate apoptosis in tumor cells. Curr. Top. Nutraceutical Res. 2004, 2, 93–100. [Google Scholar]
- Ivorra, M.; D’ocon, M.; Paya, M.; Villar, A. Antihyperglycemic and insulin-releasing effects of beta-sitosterol 3-beta-d-glucoside and its aglycone, beta-sitosterol. Arch. Int. Pharmacodyn. Thér. 1987, 296, 224–231. [Google Scholar]
- Chai, J.-W.; Lim, S.-L.; Kanthimathi, M.; Kuppusamy, U.R. Gene regulation in β-sitosterol-mediated stimulation of adipogenesis, glucose uptake, and lipid mobilization in rat primary adipocytes. Genes Nutr. 2011, 6, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Donald, P.; Lamprecht, J.; Freestone, M.; Albrecht, C.; Bouic, P.; Kotze, D.; Van Jaarsveld, P. A randomised placebo-controlled trial of the efficacy of beta-sitosterol and its glucoside as adjuvants in the treatment of pulmonary tuberculosis. Int. J. Tuberc. Lung Dis. 1997, 1, 518–522. [Google Scholar] [PubMed]
- Kiprono, P.C.; Kaberia, F.; Keriko, J.M.; Karanja, J.N. The in vitro anti-fungal and anti-bacterial activities of β-sitosterol from Senecio lyratus (Asteraceae). Z. Naturforschung C 2000, 55, 485–488. [Google Scholar]
- Ajaiyeoba, E.; Onocha, P.; Nwozo, S.; Sama, W. Antimicrobial and cytotoxicity evaluation of Buchholzia coriacea stem bark. Fitoterapia 2003, 74, 706–709. [Google Scholar] [CrossRef]
- Mishra, G.; Singh, P.; Verma, R.; Kumar, S.; Srivastav, S.; Jha, K.; Khosa, R. Traditional uses, phytochemistry and pharmacological properties of Moringa oleifera plant: An overview. Der Pham. Lett. 2011, 3, 141–164. [Google Scholar]
- Rahuman, A.A.; Gopalakrishnan, G.; Venkatesan, P.; Geetha, K. Isolation and identification of mosquito larvicidal compound from Abutilon indicum (Linn.) sweet. Parasitol. Res. 2008, 102, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Nweze, N.E. In vitro anti-trypanosomal activity of Morinda lucida leaves. Afr. J. Biotechnol. 2011, 11, 1812–1817. [Google Scholar]
- Bumrela, S.; Naik, S. Hepato protective activity of methanolic extract of Dipteracanthus patulus (Jacq) Nees: Possible involvement of antioxidant and membrane stabilization property. Int. J. Pharm. Pharm. Sci. 2012, 4, 685–690. [Google Scholar]
- Kanokmedhakul, K.; Kanokmedhakul, S.; Phatchana, R. Biological activity of anthraquinones and triterpenoids from Prismatomeris fragrans. J. Ethnopharmacol. 2005, 100, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.-B.; Oh, M.-N.; Kim, J.-G.; Shin, D.-S.; Shin, J. Inhibition of sortase-mediated Staphylococcus aureus adhesion to fibronectin via fibronectin-binding protein by sortase inhibitors. Appl. Microbiol. Biotechnol. 2006, 70, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Villaseñor, I.M.; Angelada, J.; Canlas, A.P.; Echegoyen, D. Bioactivity studies on β-sitosterol and its glucoside. Phytother. Res. 2002, 16, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Deepak, M.; Dipankar, G.; Prashanth, D.; Asha, M.; Amit, A.; Venkataraman, B. Tribulosin and β-sitosterol-d-glucoside, the anthelmintic principles of Tribulus terrestris. Phytomedicine 2002, 9, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Saha, A.; Chatterjee, I.; Chakravarty, A. Viper and cobra venom neutralization by β-sitosterol and stigmasterol isolated from the root extract of Pluchea indica Less (Asteraceae). Phytomedicine 2007, 14, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ye, W.; Yin, Z.; Zhao, S. Chemical constituents from flos Sesamum indicum L. Yao Xue Xue Bao 2007, 42, 286–291. [Google Scholar] [PubMed]
- Upadhyay, K.; Gupta, N.K.; Dixit, V.K. Development and characterization of phyto-vesicles of β-sitosterol for the treatment of androgenetic alopecia. Arch. Dermatol. Res. 2012, 304, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Prager, N.; Bickett, K.; French, N.; Marcovici, G. A randomized, double-blind, placebo-controlled trial to determine the effectiveness of botanically derived inhibitors of 5-α-reductase in the treatment of androgenetic alopecia. J. Altern. Complement. Med. 2002, 8, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Klippel, K.; Hiltl, D.; Schipp, B. A multicentric, placebo-controlled, double-blind clinical trial of β-sitosterol (phytosterol) for the treatment of benign prostatic hyperplasia. Br. J. Urol. 1997, 80, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Breza, J.; Dzurny, O.; Borowka, A.; Hanus, T.; Petrik, R.; Blane, G.; Chadha-Boreham, H. Efficacy and acceptability of tadenan®(Pygeum africanum extract) in the treatment of benign prostatic hyperplasia (BPH): A multicentre trial in central Europe. Curr. Med. Res. Opin. 1998, 14, 127–139. [Google Scholar] [PubMed]
- Berges, R.; Windeler, J.; Trampisch, H.; Senge, T.; β-sitosterol study group. Randomised, placebo-controlled, double-blind clinical trial of β-sitosterol in patients with benign prostatic hyperplasia. Lancet 1995, 345, 1529–1532. [Google Scholar] [CrossRef]
- Barry, M.J.; Roehrborn, C.G. Extracts from “clinical evidence”: Benign prostatic hyperplasia. BMJ 2001, 323, 1042. [Google Scholar] [CrossRef] [PubMed]
- Lomenick, B.; Shi, H.; Huang, J.; Chen, C. Identification and characterization of β-sitosterol target proteins. Bioorg. Med. Chem. Lett. 2015, 25, 4976–4979. [Google Scholar] [CrossRef] [PubMed]
- Assmann, G.; Cullen, P.; Erbey, J.; Ramey, D.R.; Kannenberg, F.; Schulte, H. Plasma sitosterol elevations are associated with an increased incidence of coronary events in men: Results of a nested case-control analysis of the prospective cardiovascular münster (PROCAM) study. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Hedman, M.; Miettinen, T.A.; Gylling, H.; Ketomäki, A.; Antikainen, M. Serum noncholesterol sterols in children with heterozygous familial hypercholesterolemia undergoing pravastatin therapy. J. Pediatr. 2006, 148, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Thongtang, N.; Lin, J.; Schaefer, E.J.; Lowe, R.S.; Tomassini, J.E.; Shah, A.K.; Tershakovec, A.M. Effects of ezetimibe added to statin therapy on markers of cholesterol absorption and synthesis and LDL-C lowering in hyperlipidemic patients. Atherosclerosis 2012, 225, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Yamanashi, Y.; Takada, T.; Suzuki, H. Niemann-Pick C1-like 1 overexpression facilitates ezetimibe-sensitive cholesterol and β-sitosterol uptake in Caco-2 cells. J. Pharmacol. Exp. Ther. 2007, 320, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Katz, M. Corn oil and its minor constituents as inhibitors of DMBA-induced chromosomal breaks in vivo. Mutation Res. 1984, 136, 247–253. [Google Scholar] [CrossRef]
- Maguire, L.; Konoplyannikov, M.; Ford, A.; Maguire, A.R.; O’Brien, N.M. Comparison of the cytotoxic effects of β-sitosterol oxides and a cholesterol oxide, 7β-hydroxycholesterol, in cultured mammalian cells. Br. J. Nutr. 2003, 90, 767–775. [Google Scholar] [CrossRef] [PubMed]
- MacLatchy, D.; Peters, L.; Nickle, J.; van Der Kraak, G. Exposure to β-sitosterol alters the endocrine status of goldfish differently than 17β-estradiol. Environ. Toxicol. Chem. 1997, 16, 1895–1904. [Google Scholar] [CrossRef]
- Nieminen, P.; Pölönen, I.; Ikonen, K.; Määttänen, M.; Mustonen, A.-M. Evaluation of reproductive safety of β-sitosterol on the American mink (neovison vison). Chemosphere 2008, 71, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X.; Liu, H.; Guo, H.; Zhang, M.; Mei, D.; Liu, C.; He, L.; Liu, L.; Liu, X. Impaired hepatic and intestinal ATP-binding cassette transporter G5/8 was associated with high exposure of β-sitosterol and the potential risks to blood–brain barrier integrity in diabetic rats. J. Pharm. Pharmacol. 2014, 66, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Carey, M.C. Thermodynamic and molecular determinants of sterol solubilities in bile salt micelles. J. Lipid Res. 1987, 28, 1144–1155. [Google Scholar] [PubMed]
- Imanaka, H.; Koide, H.; Shimizu, K.; Asai, T.; Kinouchi Shimizu, N.; Ishikado, A.; Makino, T.; Oku, N. Chemoprevention of tumor metastasis by liposomal. Beta-sitosterol intake. Biol. Pharm. Bull. 2008, 31, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Borowy-Borowski, H.; Sodja, C.; Docherty, J.; Roy Walker, P.; Sikorska, M. Unique technology for solubilization and delivery of highly lipophilic bioactive molecules. J. Drug Target. 2004, 12, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, B.; Bergenståhl, B.; Kronberg, B. A method for the preparation of submicron particles of sparingly water-soluble drugs by precipitation in oil-in-water emulsions. II: Influence of the emulsifier, the solvent, and the drug substance. J. Pharm. Sci. 1993, 82, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, L.; Yang, Y.; Xu, X.; Huang, Y. Tumor targeting by pH-sensitive, biodegradable, cross-linked N-(2-hydroxypropyl) methacrylamide copolymer micelles. Biomaterials 2014, 35, 6622–6635. [Google Scholar] [CrossRef] [PubMed]
- Lacatusu, I.; Badea, N.; Stan, R.; Meghea, A. Novel bio-active lipid nanocarriers for the stabilization and sustained release of sitosterol. Nanotechnology 2012, 23, 455702. [Google Scholar] [CrossRef] [PubMed]
- Farkas, E.; Schubert, R.; Zelkó, R. Effect of β-sitosterol concentration and high pressure homogenization on the chlorhexidine release from vesicular gels. Int. J. Pharm. 2006, 307, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.P.; Jarowski, C.I. Oral absorption efficiency of acid-labile antibiotics from lipid-drug delivery systems. J. Pharm. Sci. 1975, 64, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Joseph, A.; Hill, J.; Patel, J.; Patel, S.; Kincl, F.A. Sustained-release hormonal preparations XV: Release of progesterone from cholesterol pellets in vivo. J. Pharm. Sci. 1977, 66, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Maitani, Y.; Yano, S.; Hattori, Y.; Furuhata, M.; Hayashi, K. Liposome vector containing biosurfactant-complexed DNA as herpes simplex virus thymidine kinase gene delivery system. J. Liposome Res. 2006, 16, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qi, X.R.; Gao, Y.; Wei, L.; Maitani, Y.; Nagai, T. Mechanisms of co-modified liver-targeting liposomes as gene delivery carriers based on cellular uptake and antigens inhibition effect. J. Control. Release 2007, 117, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Hiruta, Y.; Hattori, Y.; Kawano, K.; Obata, Y.; Maitani, Y. Novel ultra-deformable vesicles entrapped with bleomycin and enhanced to penetrate rat skin. J. Control. Release 2006, 113, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Maitani, Y.; Nakamura, K.; Suenaga, H.; Kamata, K.; Takayama, K.; Nagai, T. The enhancing effect of soybean-derived sterylglucoside and β-sitosterol β-d-glucoside on nasal absorption in rabbits. Int. J. Pharm. 2000, 200, 17–26. [Google Scholar] [CrossRef]
- Nakamura, K.; Takayama, K.; Nagai, T.; Maitani, Y. Regional intestinal absorption of FITC–dextran 4,400 with nanoparticles based on β-sitosterol β-d-glucoside in rats. J. Pharm. Sci. 2003, 92, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Elmowafy, M.; Viitala, T.; Ibrahim, H.M.; Abu-Elyazid, S.K.; Samy, A.; Kassem, A.; Yliperttula, M. Silymarin loaded liposomes for hepatic targeting: In vitro evaluation and HepG2 drug uptake. Eur. J. Pharm. Sci. 2013, 50, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Hench, P.S.; Kendall, E.C.; Slocumb, C.H.; Polley, H.F. Effects of cortisone acetate and pituitary ACTH on rheumatoid arthritis, rheumatic fever and certain other conditions: A study in clinical physiology. Arch. Intern. Med. 1950, 85, 545–666. [Google Scholar] [CrossRef]
- Dixon, W.G.; Bansback, N. Understanding the side effects of glucocorticoid therapy: Shining a light on a drug everyone thinks they know. Ann. Rheum. Dis. 2012, 71, 1761–1764. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Fiorito, A.; Panourgia, M.P.; Sangiorgi, Z.; Gaddi, A. Effects of a new soy/β-sitosterol supplement on plasma lipids in moderately hypercholesterolemic subjects. J. Am. Diet. Assoc. 2002, 102, 1807–1811. [Google Scholar] [CrossRef]
- Louw, I.; Halland, A.; Bouic, P.; Freestone, M.; Lamprecht, J. A pilot study of the clinical effects of a mixture of beta-sitosterol and beta-sitosterol glucoside in active rheumatoid arthritis. Am. J. Clin. Nutr. 2002, 75, 351S. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
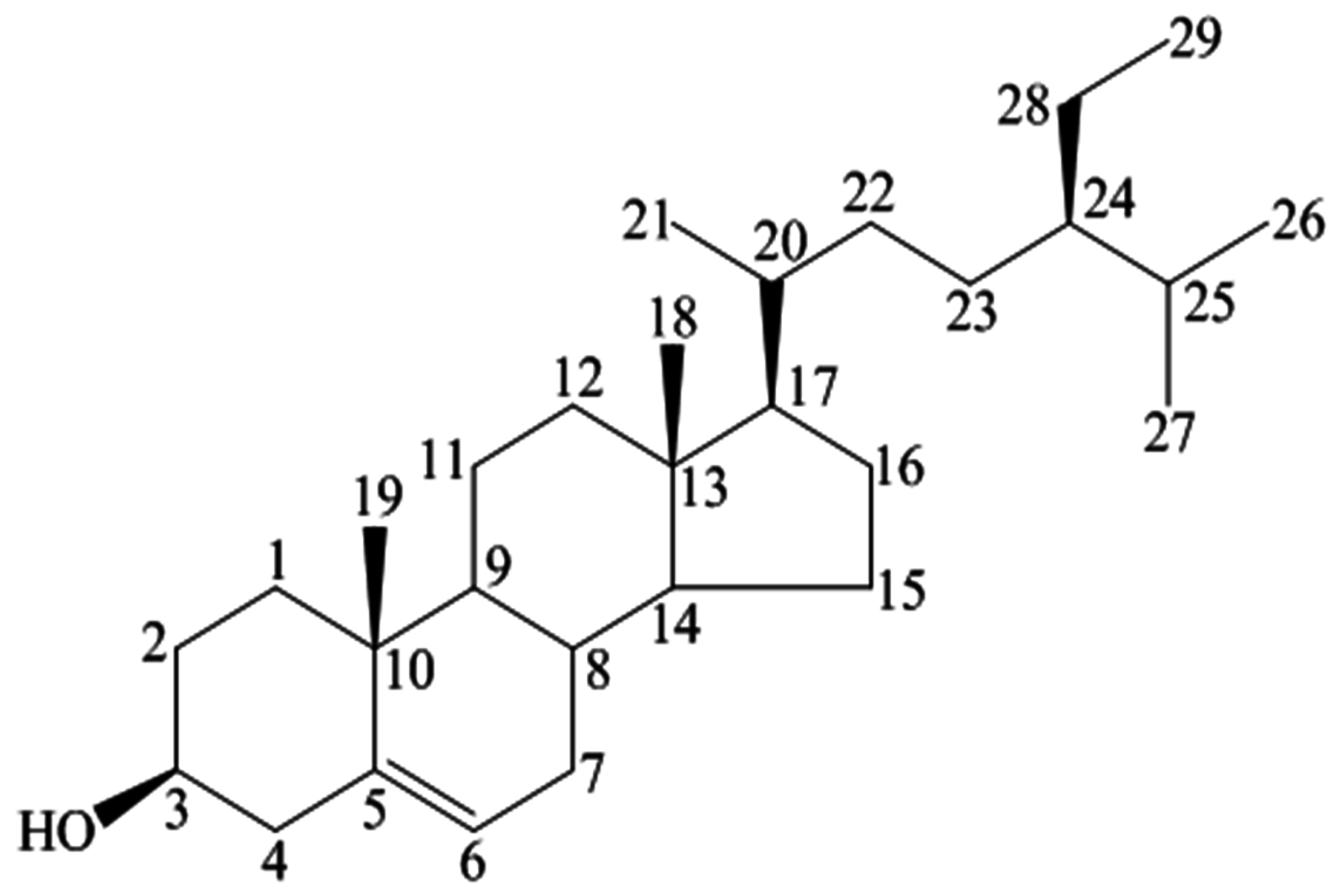
| Position | 1H | 13C |
|---|---|---|
| 3 | 3.53 (tdd, 1H, J = 4.5, 4.2, 3.8 Hz) | 72.0 |
| 5 | 5.36 (t, 1H, J = 6.4 Hz) | 140.9 |
| 18 | 1.01 (s, 3H) | 12.0 |
| 19 | 0.68 (s, 3H) | 19.0 |
| 21 | 0.93 (d,3H, J = 6.5 Hz) | 19.2 |
| 26 | 0.83 (d, 3H, J = 6.4 Hz) | 20.1 |
| 27 | 0.81 (d, 3H, J = 6.4 Hz) | 19.6 |
| 29 | 0.84 (t, 3H, J = 7.2 Hz) | 12.2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bin Sayeed, M.S.; Karim, S.M.R.; Sharmin, T.; Morshed, M.M. Critical Analysis on Characterization, Systemic Effect, and Therapeutic Potential of Beta-Sitosterol: A Plant-Derived Orphan Phytosterol. Medicines 2016, 3, 29. https://doi.org/10.3390/medicines3040029
Bin Sayeed MS, Karim SMR, Sharmin T, Morshed MM. Critical Analysis on Characterization, Systemic Effect, and Therapeutic Potential of Beta-Sitosterol: A Plant-Derived Orphan Phytosterol. Medicines. 2016; 3(4):29. https://doi.org/10.3390/medicines3040029
Chicago/Turabian StyleBin Sayeed, Muhammad Shahdaat, Selim Muhammad Rezaul Karim, Tasnuva Sharmin, and Mohammed Monzur Morshed. 2016. "Critical Analysis on Characterization, Systemic Effect, and Therapeutic Potential of Beta-Sitosterol: A Plant-Derived Orphan Phytosterol" Medicines 3, no. 4: 29. https://doi.org/10.3390/medicines3040029
APA StyleBin Sayeed, M. S., Karim, S. M. R., Sharmin, T., & Morshed, M. M. (2016). Critical Analysis on Characterization, Systemic Effect, and Therapeutic Potential of Beta-Sitosterol: A Plant-Derived Orphan Phytosterol. Medicines, 3(4), 29. https://doi.org/10.3390/medicines3040029