Medicinal Potentialities of Plant Defensins: A Review with Applied Perspectives



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
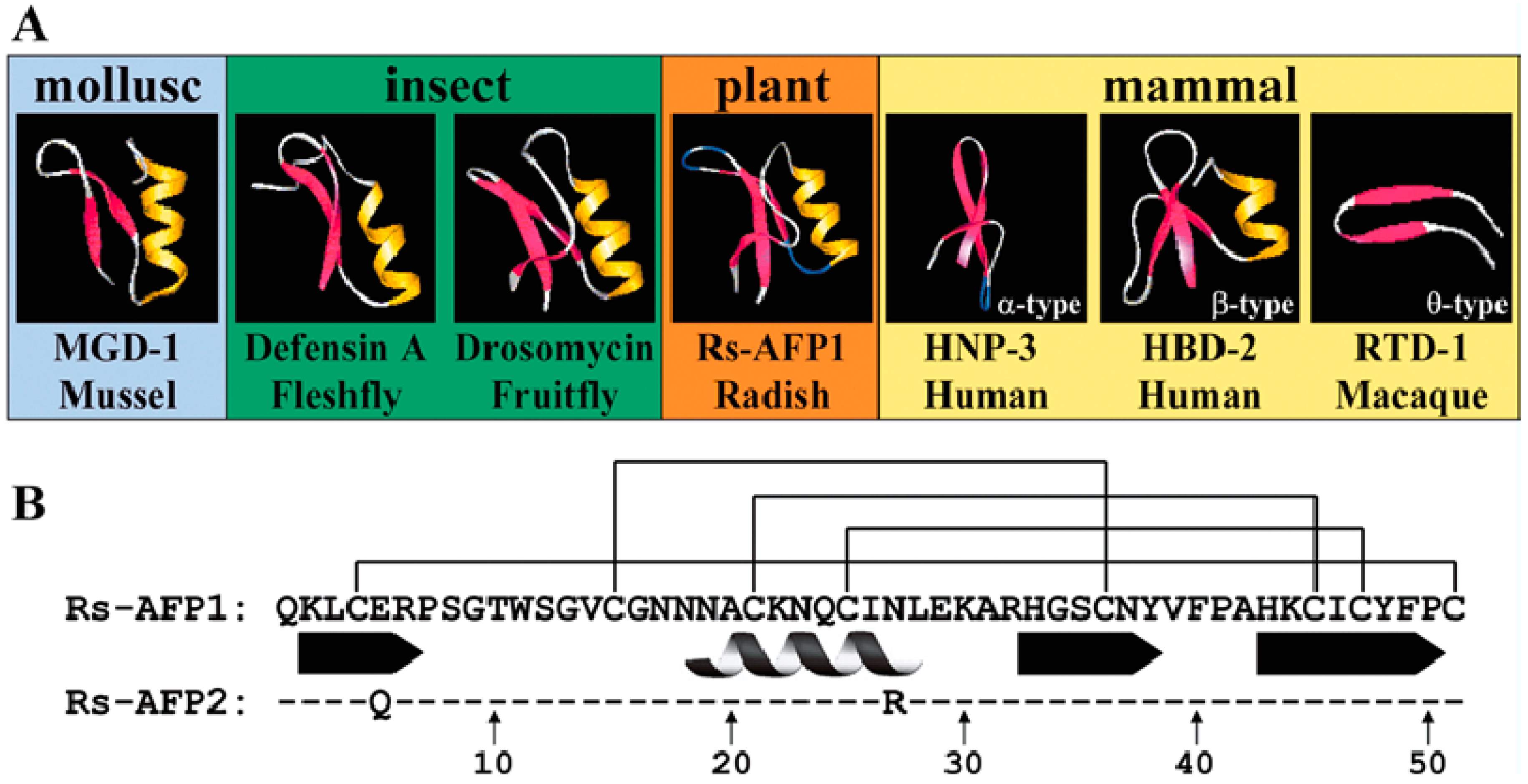
2. Origin of Defensins
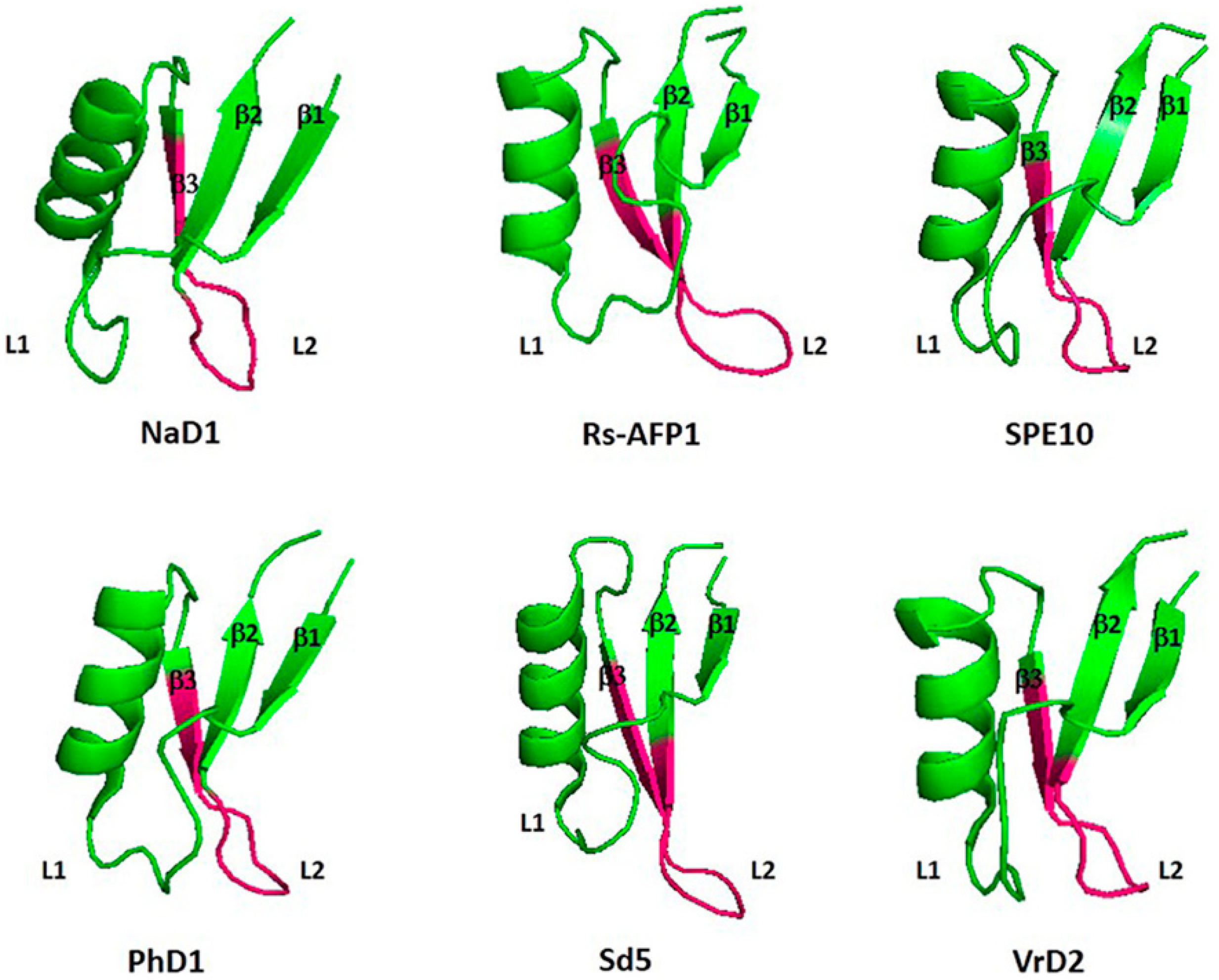
3. Structure of Defensins
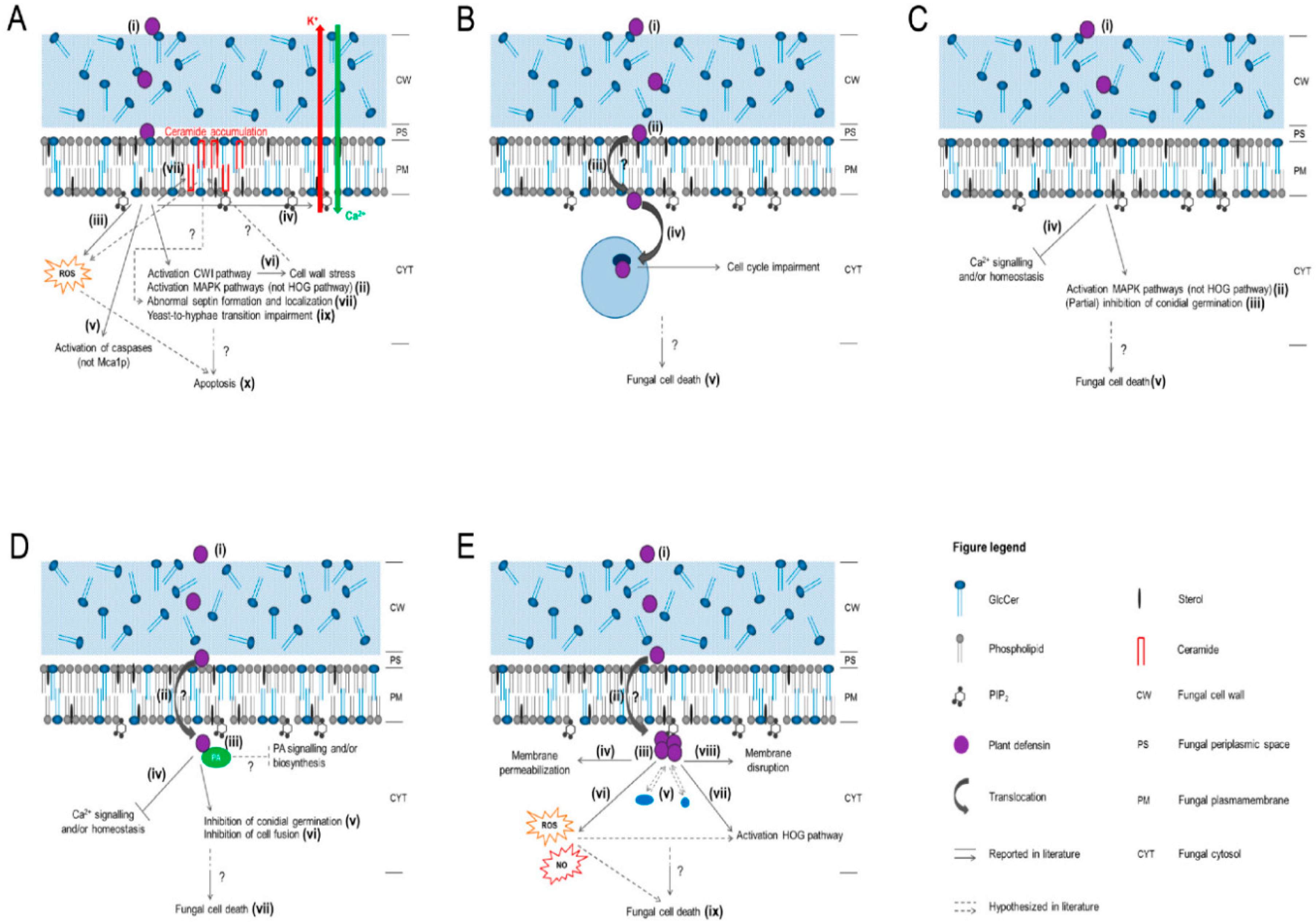
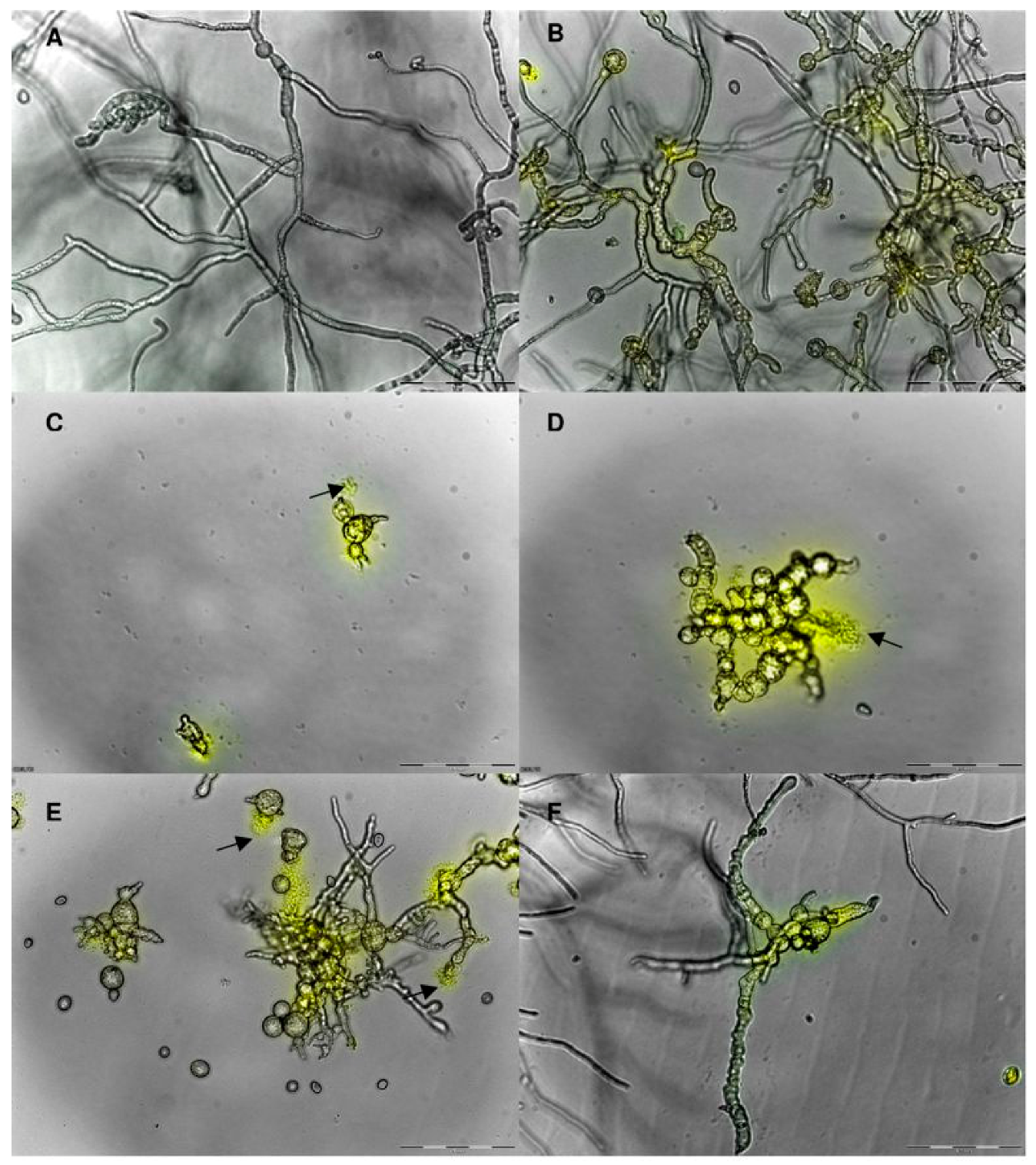
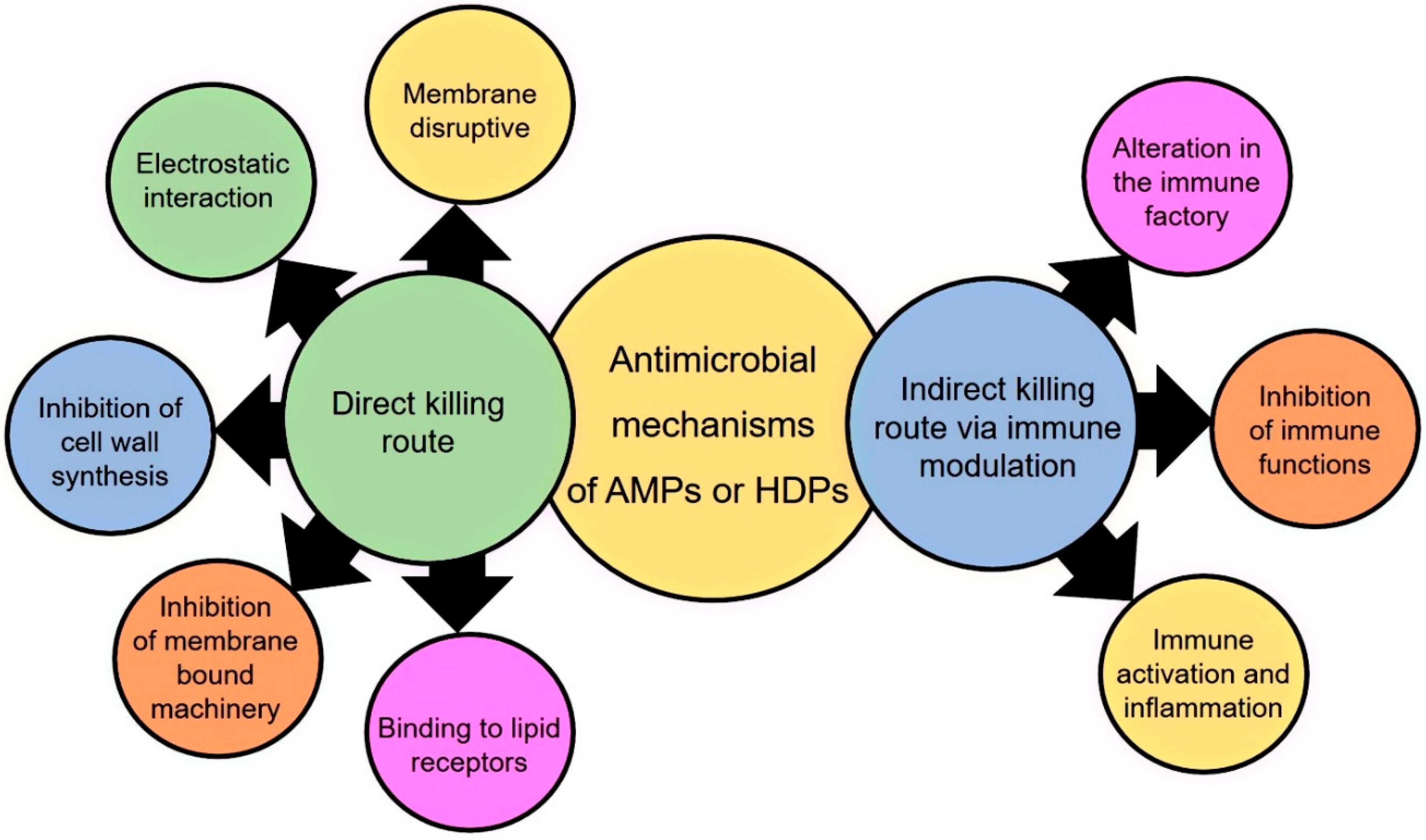
4. Mode of Action
5. Role of Defensins in Plant Defense
6. Peptides Involved in the Stress Response
7. Involvement of Peptides in Reproduction
8. Biological Functionalities of Plant Defensins
8.1. Plant Defensins—Antimicrobial Activity
8.2. Plant Defensins—Anticancer Activity
9. Concluding Remarks and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gachomo, E.W.; Seufferheld, M.J.; Kotchoni, S.O. Melanization of appressoria is critical for the pathogenicity of Diplocarpon rosae. Mol. Biol. Rep. 2010, 37, 3583–3591. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Ann. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Lacerda, A.; Vasconcelos, É.A.R.; Pelegrini, P.B.; Grossi-de-Sa, M.F. Antifungal defensins and their role in plant defense. Front. Microbial. 2014, 5, 116. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, B.; Tytgat, J. Phylogenetic distribution, functional epitopes and evolution of the CSab superfamily. Cell Mol. Life Sci. 2005, 62, 2257–2269. [Google Scholar] [CrossRef]
- Thomma, B.P.; Cammue, B.P.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, A.O.; Gomes, V.M. Plant defensins—Prospects for the biological functions and biotechnological properties. Peptides 2009, 30, 1007–1020. [Google Scholar] [CrossRef]
- Colilla, F.J.; Rocher, A.; Mendez, E. gamma-purothionins: Amino acid sequence of two polypeptides of a new family of thionins from wheat endosperm. FEBS Lett. 1990, 270, 191–194. [Google Scholar] [CrossRef]
- Mendez, E.; Moreno, A.; Colilla, F.; Pelaez, F.; Limas, G.G.; Mendez, R.; Soriano, F.; Salinas, M.; de Haro, C. Primary structure and inhibition of protein synthesis in eukaryotic cell-free system of a novel thionin, g-hordothionin, from barley endosperm. Eur. J. Biochem. 1990, 194, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Pelegrini, P.B.; Franco, O.L. Plant g-thionins: Novel sinsites on the mechanisms of actions of a multi-functional class of defense proteins. Int. J. Biochem. Cell Biol. 2005, 37, 2239–2253. [Google Scholar] [CrossRef]
- Bruix, M.; Gonzalez, C.; Santoro, J.; Soriano, F.; Rocher, A.; Mendez, E.; Rico, M. 1H-NMR studies on the structure of a new thionin from barley endosperm. Biopolymers 1995, 36, 751–763. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Terras, F.R.G.; Cammue, B.P.A.; Osborn, R.W. Plant Defensins—Novel antimicrobial peptides as components of the host-defense system. Plant Physiol. 1995, 108, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Cociancich, S.; Goyffon, M.; Bontems, F.; Bulet, P.; Bouet, F.; Menez, A.; Hoffmann, J. Purification and characterization of a scorpion defensin, a 4 kDa antibacterial peptide presenting structural similarities with insect defensis and scorpion toxins. Biochem. Biophys. Res. Commun. 1993, 194, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Kawabata, S.; Shigenaga, T.; Takayenoki, Y.; Cho, J.; Nakajima, H.; Hirat, M.; Iwanaga, S. A novel big defensin identified in horseshoe crab hemocytes: Isolation, amino acid sequence, and antibacterial activity. J. Biochem. 1995, 117, 1131–1137. [Google Scholar] [CrossRef]
- Charlet, M.; Chernysh, S.; Philippe, H.; Hetru, C.; Hoffmann, J.A.; Bulet, P. Isolation of several cysteine-rich antimicrobial peptides from the blood of a mollusc, Mytilus edulis. J. Biol. Chem. 1996, 271, 21808–21813. [Google Scholar] [CrossRef] [PubMed]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; So¨nksen, C.P.; Ludvigsen, S.; Raventós, D.; Buskov, S.; Christensen, B.; De Maria, L.; et al. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef]
- Zou, J.; Mercier, C.; Koussounadis, A.; Secombes, C. Discovery of multiple beta defensing like homologues in teleost fish. Mol. Immunol. 2007, 44, 638–647. [Google Scholar] [CrossRef]
- Sugiarto, H.; Yu, P.-L. Avian antimicrobial peptides: The defense role of bdefensins. Biochem. Biophys. Res. Commun. 2004, 323, 721–727. [Google Scholar] [CrossRef]
- Zhu, S. Evidence for myxobacterial origin of eukaryotic defensins. Immunogenetics 2007, 59, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S. Discovery of six families of fungal defensin-like peptides provides insights into origin and evolution of the CSab defensins. Mol. Immunol. 2008, 45, 828–838. [Google Scholar] [CrossRef]
- Lay, F.T.; Anderson, M.A. Defensins–components of the innate immune system in plants. Curr. Protein Pept. Sci. 2005, 6, 85–101. [Google Scholar] [CrossRef]
- Janssen, B.J.C.; Schirra, H.J.; Lay, F.T.; Anderson, M.A.; Craik, D.J. Structure of Petunia hybrid defensing 1, a novel plant defensen with five disulfide bonds. Biochemistry 2003, 42, 8214–8222. [Google Scholar] [CrossRef] [PubMed]
- De Coninck, B.; Cammue, B.P.A.; Thevissen, K. Modes of antifungal action and in planta functions of plant defensins and defensin-like peptides. Fungal Biol. Rev. 2013, 26, 109–120. [Google Scholar] [CrossRef]
- Vander Weerden, N.; Anderson, M.A. Plant defensins: Commons fold, multiple functions. Fungal Biol. Rev. 2013, 26, 121–131. [Google Scholar] [CrossRef]
- Vriens, K.; Cammue, B.; Thevissen, K. Antifungal plant defensins: Mechanisms of action and production. Molecules 2014, 19, 12280–12303. [Google Scholar] [CrossRef]
- Hegedus, N.; Marx, F. Antifungal proteins: More than antimicrobials? Fungal Biol. Rev. 2013, 26, 132–145. [Google Scholar] [CrossRef]
- Wilmes, M.; Cammuer, B.P.A.; Sahl, H.-G.; Thevisse, K. Antibiotic activities of host defense peptides: More to it than lipid bilayer perturbation. Nat. Prod. Rep. 2011, 28, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Fant, F.; Vranken, W.; Broekaert, W.; Borremans, F. Determination of the three-dimensional solution structure of Raphanus sativus antifungal protein 1 by 1HNMR. J. Mol. Biol. 1998, 279, 257–270. [Google Scholar] [CrossRef]
- Lay, F.T.; Schirra, H.J.; Scalon, M.J.; Anderson, M.A.; Craik, D.J. The three-dimensional solution structure of NaD1, a new floral defensin from Nicotiana alata and its application to a homology model of the crop defense protein alfAFP. J. Mol. Biol. 2003, 325, 175–188. [Google Scholar] [CrossRef]
- Sagaram, U.S.; Pandurangi, R.; Karu, J.; Smith, T.J.; Shah, D.M. Structure-activity determinants in antifungal plant defensins MsDef1 and MtDef4 with different modes of action against Fusarium graminearum. PLoS ONE 2011, 6, e18550. [Google Scholar] [CrossRef] [PubMed]
- Selitrennikoff, C.P. Antifungal proteins. App. Environ. Microbiol. 2001, 67, 2883–2894. [Google Scholar] [CrossRef]
- de Beer, A.; Vivier, M.A. Four plant defensins from an indigenous South African Brassicaceae species display divergent activities against two test pathogens despite high sequence similarity in the encoding genes. BMC Res. Notes 2011, 4, 459. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Olmedo, F.; Molina, A.; Alamillo, J.M.; Rodriguez-Palenzuela, P. Plant defense peptides. Biopolymers 1998, 47, 479–491. [Google Scholar] [CrossRef]
- Chen, K.-C.; Lin, C.-Y.; Chung, M.-C.; Kuan, C.C.; Sung, H.Y.; Tsou, S.C.S.; Kuo, G.; Chen, C.-S. Cloning and characterization of a cDNA encoding an antimicrobial protein from mung bean seeds. Bot. Bull. Acad. Sin. 2002, 43, 251–259. [Google Scholar]
- de Oliveira Carvalho, A.; Moreira Gomes, V. Plant defensins and defensin-like peptides-biological activities and biotechnological applications. Curr. Pharm. Des. 2011, 7, 4270–4293. [Google Scholar] [CrossRef]
- Franco, O.L. Peptide promiscuity: An evolutionary concept for plant defense. FEBS Lett. 2011, 585, 995–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione is a key player in metal-induced oxidative stress defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Yeaman, M.R. Multidimensional signatures in antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2004, 101, 7363–7368. [Google Scholar] [CrossRef]
- Méndez, E.; Rocher, A.; Calero, M.; Girbés, T.; Citores, L.; Soriano, F. Primary structure of omega-hordothionin, a member of a novel family of thionins from barley endosperm, and its inhibition of protein synthesis in eukaryotic and prokaryotic cell-free systems. Eur. J. Biochem. 1996, 239, 67–73. [Google Scholar] [CrossRef]
- Mirouze, M.; Sels, J.; Richard, O.; Czernic, P.; Loubet, S.; Jacquier, A.; François, I.E.; Cammue, B.; Lebrun, M.; Berthomieu, P. A putative novel role for plantd efensins: A defensin from the zinc hyper-accumulating plant, Arabidopsis halleri, confers zinc tolerance. Plant J. 2006, 47, 329–342. [Google Scholar] [CrossRef]
- Ferreira, R.B.; Monteiro, S.A.R.A.; Freitas, R.; Santos, C.N.; Chen, Z.; Batista, L.M.; Teixeira, A.R. The role of plant defence proteins in fungal pathogenesis. Mol. Plant Pathol. 2007, 8, 677–700. [Google Scholar] [CrossRef] [PubMed]
- Parisi, K.; Shafee, T.M.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell. Dev. Biol. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Terras, F.R.G.; Schoofs, H.M.E.; De Bolle, M.F.C.; Van Leuven, F.; Rees, S.B.; Vanderleyden, J.; Cammue, B.P.; Broekaert, W.F. Analysis of two novel classes of plant antifungal proteins from radish (Raphanus sativus L.) seeds. J. Biol. Chem. 1992, 267, 15301–15309. [Google Scholar] [PubMed]
- Fujimuram, M.; Ideguchi, M.; Minami, Y.; Watanabe, K.; Tadera, K. Purification, characterization and sequencing of novel antimicrobial peptides Tu-AMP1 and Tu-AMP2 from bulbs of tulip (Tulipa gesneriana L.). Biosci. Biotechnol. Biochem. 2004, 68, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-H.; Hsu, M.-P.; Tan, C.-H.; Sung, H.-Y.; Kuo, C.G.; Fan, M.-J.; Chen, H.-M.; Chen, S.; Chen, C.-S. Cloning and characterization of a plant defensin VaD1 from azuki bean. J. Agric. Food Chem. 2005, 53, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. Vulgarinin, a broad-spectrum antifungal peptide from haricot beans (Phaseolus vulgaris). IJBCB 2005, 37, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.L.; Murad, A.M.; Leite, J.R.; Mendes, P.A.M.; Prates, M.V.; Bloch, C., Jr. Identification of a cowpea g-thionin with bactericidal activity. FEBS J. 2006, 273, 3489–3497. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Zhang, X.Q.; Wang, H.X.; Ng, T.B. Amitogenic defensin from white cloud beans (Phaseolus vulgaris). Peptides 2006, 27, 2075–2081. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.-J.; Lai, H.-C.; Chang, Y.-S.; Sheu, M.-J.; Lu, T.-L.; Huang, S.-S.; Lin, Y.H. Antimicrobial, dehydroascorbate reductase, and monodehydroascorbate reductase activities of defensin from sweet potato [Ipomoea batatas (L.) Lam. ‘tainong 57′] storage roots. J. Agric. Food Chem. 2008, 56, 2989–2995. [Google Scholar] [CrossRef]
- van der Weerden, N.L.; Lay, F.T.; Anderson, M.A. The plant defensin, NaD1, enters the cytoplasm of Fusarium oxysporum hyphae. J. Biol. Chem. 2008, 283, 14445–14452. [Google Scholar] [CrossRef]
- Zhang, Y.; Lewis, K. Fabatins: New antimicrobial plant peptides. FEMS Microbiol. Lett. 1997, 149, 59–64. [Google Scholar]
- Thevissen, K.; Cammue, B.P.A.; Lemaire, K.; Winderickx, J.; Dickson, R.C.; Lester, R.L.; Ferket, K.K.A.; Van Even, F.; Parret, A.H.A.; Broekaert, W.F. A gene-encoding sphingolipid biosynthesis enzyme determines the sensitivity of Saccharomyces cerevisiae to an antifungal plant defensin from dahlia (Dahlia merckii). Proc. Natl. Acad. Sci. USA 2000, 97, 9531–9536. [Google Scholar] [CrossRef] [PubMed]
- Poon, I.K.; Baxter, A.A.; Lay, F.T.; Mills, G.D.; Adda, C.G.; Payne, J.A.; Phan, T.K.; Ryan, G.F.; White, J.A.; Veneer, P.K. Phosphoinositide-mediated oligomerization of a defensin induces cell lysis. Elife 2014, 3, e01808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.C.; Kang, Y.H.; Chun, H.J.; Koo, J.C.; Cheong, Y.H.; Kim, C.Y.; Kim, M.C.; Chuang, W.S.; Kim, J.C.; Yoo, J.H.; et al. Characterization of a stamen-specific cDNA encoding a novel plant defensin in Chinese cabbage. Plant Mol. Biol. 2002, 50, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Y.; Ng, T.B. A new antifungal peptide from rice beans. J. Peptide Res. 2002, 60, 81–87. [Google Scholar] [CrossRef]
- Wisniewski, M.E.; Bassett, C.L.; Artlip, T.S.; Webb, R.P.; Janisiewicz, W.J.; Norelli, J.L.; Goldway, M.; Droby, S. Characterization of a defensin in bark and fruit tissues of peach and antimicrobial activity of a recombinant defensin in the yeast, Pichia pastoris. Physiol. Plant 2003, 119, 563–572. [Google Scholar] [CrossRef]
- Vriens, K.; Peigneur, S.; De Coninck, B.; Tytgat, J.; Cammue, B.P.; Thevissen, K. The antifungal plant defensin AtPDF2. 3 from Arabidopsis thaliana blocks potassium channels. Sci. Rep. 2016, 6, 32121. [Google Scholar] [CrossRef]
- Olli, S.; Kirti, P.B. Cloning, characterization and antifungal activity of defensing Tfgd1 from Trigonella foenum-graecum L. J. Biochem. Mol. Biol. 2006, 39, 278–283. [Google Scholar]
- Solis, J.; Medrano, G.; Ghislain, M. Inhibitory effect of a defensin gene from the Andean crop maca (Lepidium meyenii) against Phytophthora infestans. J. Plant Physiol. 2007, 164, 1071–1082. [Google Scholar] [CrossRef]
- Finkina, E.I.; Shramova, E.I.; Tagaev, A.A.; Ovchinnikova, T.V. A novel defensin from the lentil Lens culinaris seeds. Biochem. Biophys. Res. Commun. 2008, 371, 860–865. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Al-Benna, S.; Shai, Y.; Jacobsen, F.; Steinstraesser, L. Oncolytic activities of host defense peptides. Int. J. Mol. Sci. 2011, 12, 8027–8051. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Rodríguez, J.J.; Ochoa-Zarzosa, A.; López-Gómez, R.; López-Meza, J.E. Plant antimicrobial peptides as potential anticancer agents. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. Sesquin, a potent defensin-like antimicrobial peptide from ground beans with inhibitory activities toward tumor cells and HIV-1 reverse transcriptase. Peptide 2005, 26, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. Limenin, a defensin-like peptide with multiple exploitable activities from shelf beans. J. Peptide Sci. 2006, 12, 341–346. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B. Lunatusin, a trypsin-stable antimicrobial peptide from Lima beans (Phaseolus lunatus L.). Peptides 2005, 26, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Wong, J.H.; Ng, T.B. A defensin with highly potent antipathogenic activities from the seeds of purple pole bean. Biosci. Rep. 2010, 30, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.H.K.; Ng, T.B. Coccinin, an antifungal peptide with antiproliferative and HIV-1 reverse transcriptase inhibitory activities from large scarlet runner beans. Peptides 2004, 25, 2063–2068. [Google Scholar] [CrossRef]
- Ngai, P.H.K.; Ng, T.B. Phaseococcin, an antifungal protein with antiproliferative and anti-HIV-1 reverse transcriptase activities from small scarlet runner beans. Biochem. Cell Biol. 2005, 83, 212–220. [Google Scholar] [CrossRef]
- Anaya-Lopez, J.L.; Lopez-Meza, J.E.; Baizabal-Aguirre, V.M.; Cano-Camacho, H.; Ochoa-Zarzosa, A. Fungicidal and cytotoxic activity of a Capsicum chinense defensin expressed by endothelial cells. Biotechnol. Lett. 2006, 28, 1101–1108. [Google Scholar] [CrossRef]
- Lobo, D.S.; Pereira, I.B.; Fragel-Madeira, L.; Medeiros, L.N.; Cabral, L.M.; Faria, J.; Bellio, M.; Campos, R.C.; Linden, R.; Kurtenbach, E. Antifungal Pisum sativum defensin 1 interacts with Neurospora crassa cyclin F related to the cell cycle. Biochemistry 2007, 46, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.S.; Cabral, K.M.S.; Kurtenbach, E.; Almeida, F.C.L.; Valente, A.P. Solution structure of Pisum sativum defensin 1 by high resolution NMR: Plant defensins, identical backbone with different mechanisms of action. J. Mol. Biol. 2002, 315, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Monk, B.C.; Harding, D.R.K. Peptides motifs for cell-surface intervention. Biodrugs 2005, 19, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J.; Cheng, C.-S.; Lai, S.-M.; Hsu, M.-P.; Chen, C.-S.; Lyu, P.-C. Solution structure of the plant defensin VrD1 from mung bean and its possible role in insecticidal activity against bruchids. Proteins 2006, 63, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyska, I.; Szachowicz-Petelska, B.; Sulkowski, S.; Figaszewski, Z. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol. Cell Biochem. 2005, 276, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Fuster, M.M.; Esko, J.D. The sweet and sour of cancer: Glycans as novel therapeutic targets. Nat. Rev. Cancer 2005, 5, 526–542. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishaq, N.; Bilal, M.; Iqbal, H.M.N. Medicinal Potentialities of Plant Defensins: A Review with Applied Perspectives. Medicines 2019, 6, 29. https://doi.org/10.3390/medicines6010029
Ishaq N, Bilal M, Iqbal HMN. Medicinal Potentialities of Plant Defensins: A Review with Applied Perspectives. Medicines. 2019; 6(1):29. https://doi.org/10.3390/medicines6010029
Chicago/Turabian StyleIshaq, Nida, Muhammad Bilal, and Hafiz M.N. Iqbal. 2019. "Medicinal Potentialities of Plant Defensins: A Review with Applied Perspectives" Medicines 6, no. 1: 29. https://doi.org/10.3390/medicines6010029
APA StyleIshaq, N., Bilal, M., & Iqbal, H. M. N. (2019). Medicinal Potentialities of Plant Defensins: A Review with Applied Perspectives. Medicines, 6(1), 29. https://doi.org/10.3390/medicines6010029