Camptothecin Induces PD-L1 and Immunomodulatory Cytokines in Colon Cancer Cells




Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Cell Culture, and Treatments
2.2. Cytokine Gene Expression and Analysis
2.3. Gene Functional Annotation
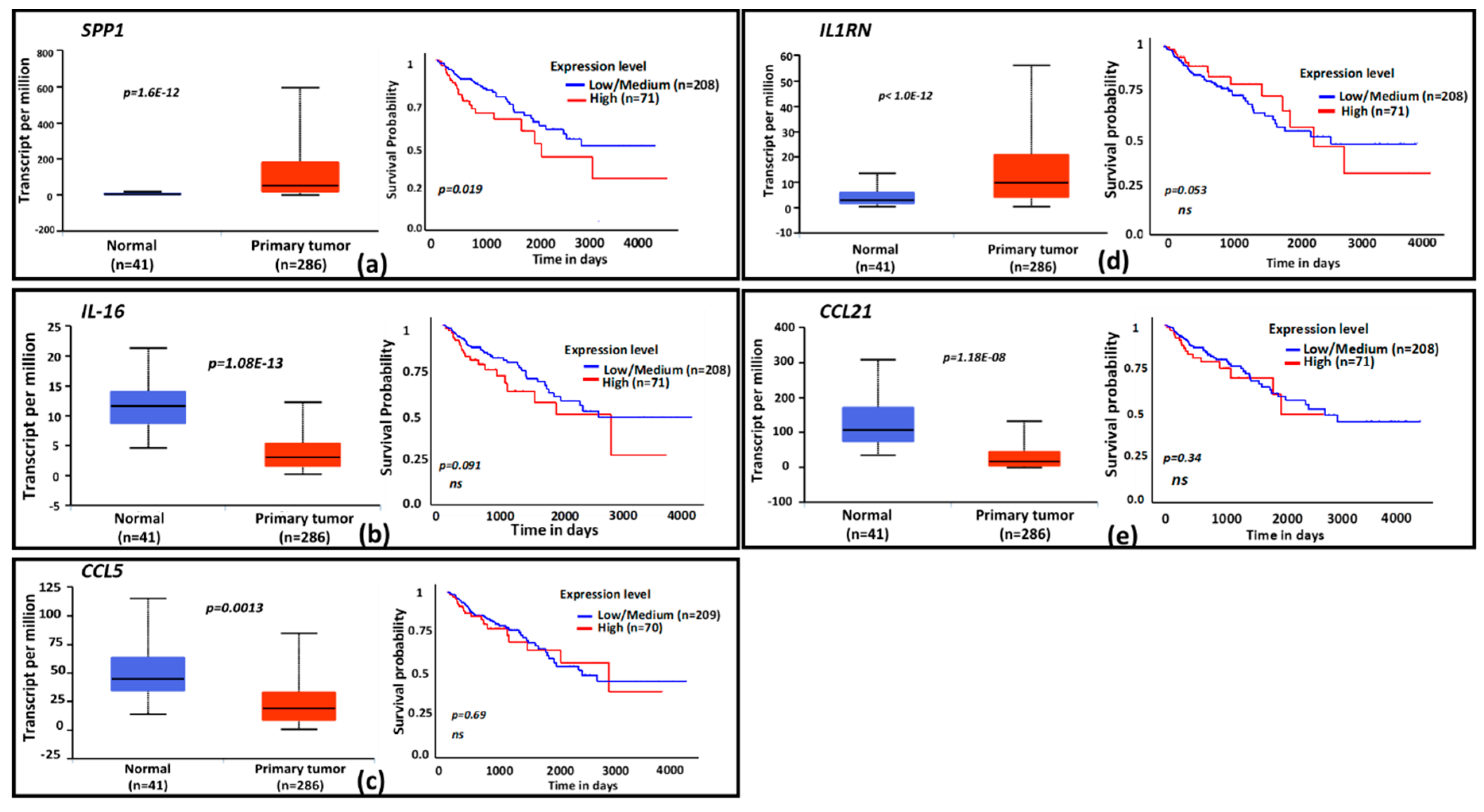
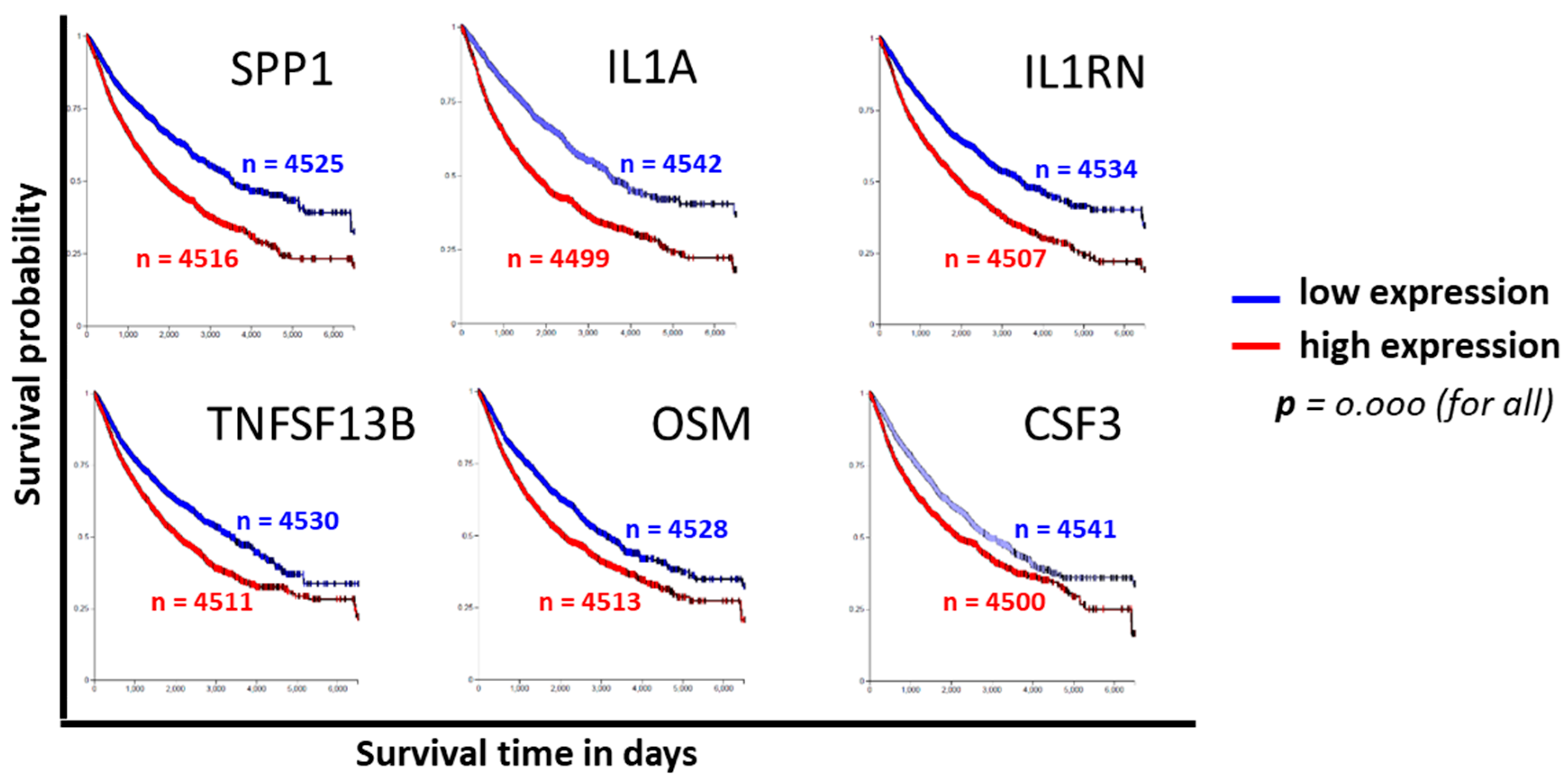
2.4. Bioinformatics Database Search and Analysis
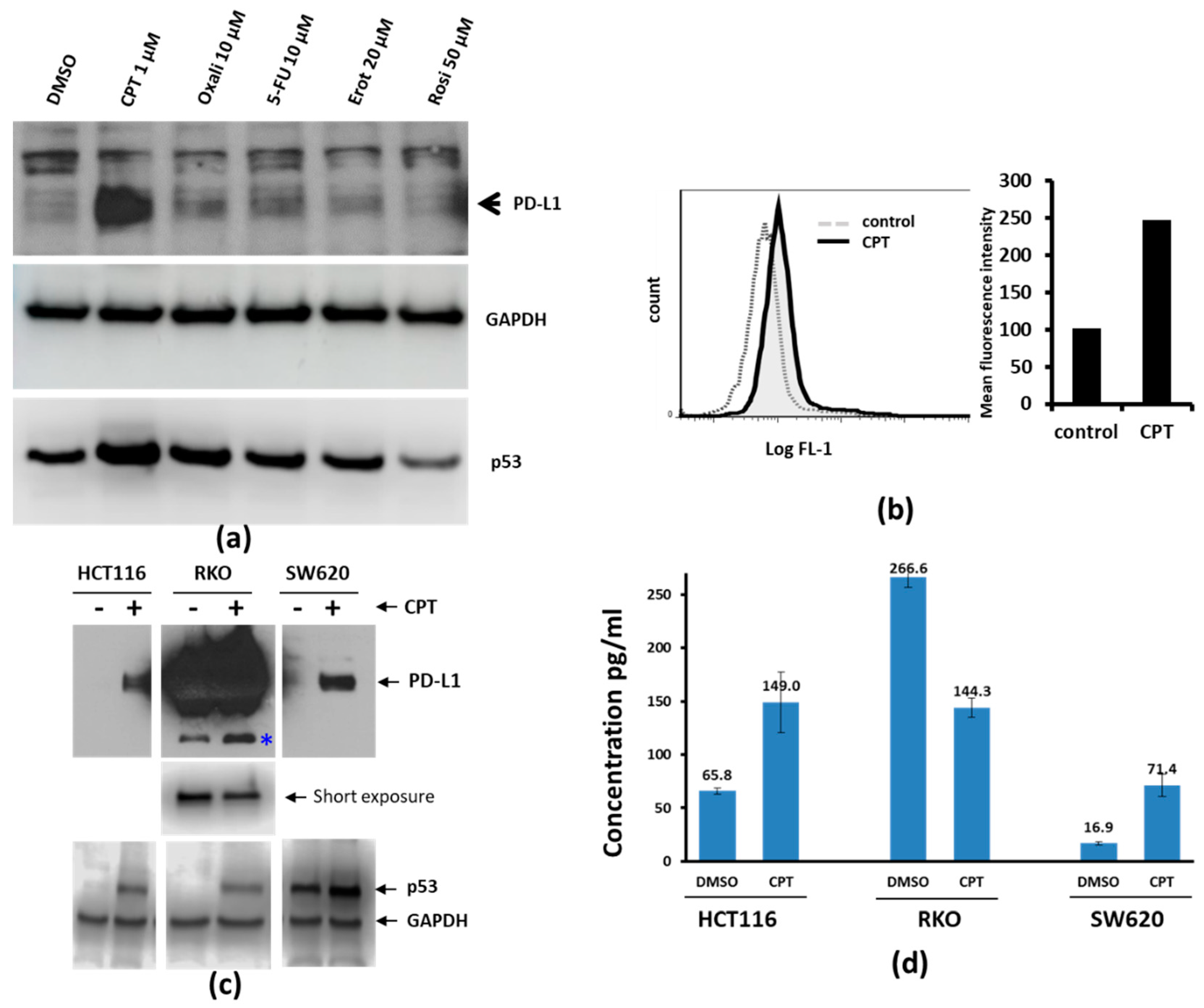
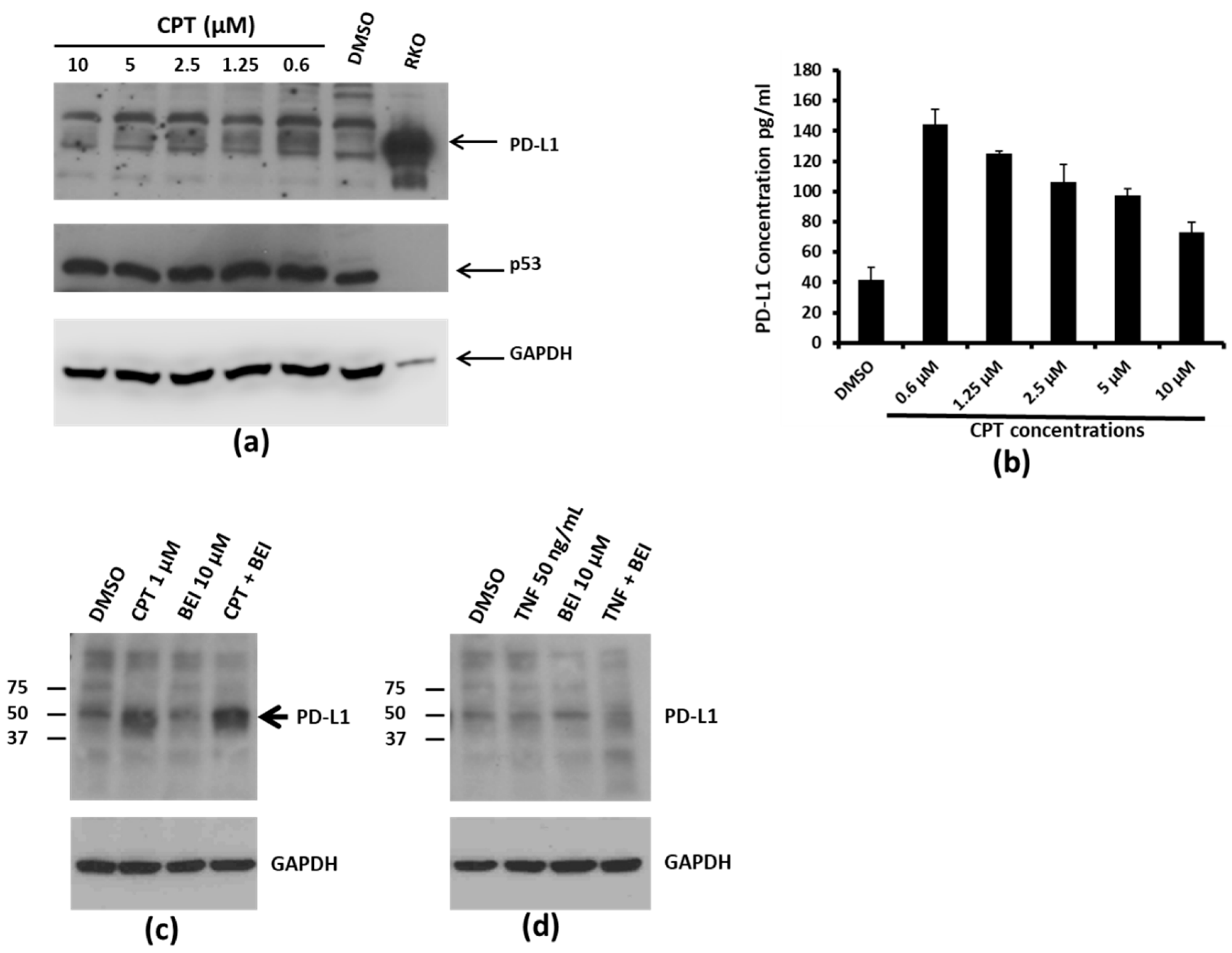
2.5. Immunoblotting
2.6. PD-L1 Enzyme Linked Immunosorbent Assay (ELISA)
2.7. Flow Cytometry for Surface Expression
3. Results
3.1. CPT Potently Upregulates PD-L1 Compared to Other Drugs
3.2. Concentration-Dependent, NF-kB Activation-Independent Upregulation of PD-L1
3.3. Cytokine Genes Induced by CPT Treatment
3.4. Clinical Relevance of CPT-Induced Cytokines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Le, D.T.; Hubbard-Lucey, V.M.; Morse, M.A.; Heery, C.R.; Dwyer, A.; Marsilje, T.H.; Brodsky, A.N.; Chan, E.; Deming, D.A.; Diaz, L.A., Jr.; et al. A Blueprint to Advance Colorectal Cancer Immunotherapies. Cancer Immunol. Res. 2017, 5, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Su, D.M.; Liang, M.; Fu, J. Chemopreventive agents induce programmed death-1-ligand 1 (PD-L1) surface expression in breast cancer cells and promote PD-L1-mediated T cell apoptosis. Mol. Immunol. 2008, 45, 1470–1476. [Google Scholar] [CrossRef]
- Van Der Kraak, L.; Goel, G.; Ramanan, K.; Kaltenmeier, C.; Zhang, L.; Normolle, D.P.; Freeman, G.J.; Tang, D.; Nason, K.S.; Davison, J.M.; et al. 5-Fluorouracil upregulates cell surface B7-H1 (PD-L1) expression in gastrointestinal cancers. J. Immunother. Cancer 2016, 4, 65. [Google Scholar] [CrossRef]
- Yan, F.; Pang, J.; Peng, Y.; Molina, J.R.; Yang, P.; Liu, S. Elevated Cellular PD1/PD-L1 Expression Confers Acquired Resistance to Cisplatin in Small Cell Lung Cancer Cells. PLoS ONE 2016, 11, e0162925. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Demaria, S.; Formenti, S. Current clinical trials testing the combination of immunotherapy with radiotherapy. J. Immunother. Cancer 2016, 4, 51. [Google Scholar] [CrossRef]
- Blank, C.; Mackensen, A. Contribution of the PD-L1/PD-1 pathway to T-cell exhaustion: An update on implications for chronic infections and tumor evasion. Cancer Immunol. Immunother. CII 2007, 56, 739–745. [Google Scholar] [CrossRef]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K.; et al. Tumor-associated B7-H1 promotes T-cell apoptosis: A potential mechanism of immune evasion. Nat. Med. 2002, 8, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Chacon, J.A.; Schutsky, K.; Powell, D.J. The Impact of Chemotherapy, Radiation and Epigenetic Modifiers in Cancer Cell Expression of Immune Inhibitory and Stimulatory Molecules and Anti-Tumor Efficacy. Vaccines 2016, 4, 43. [Google Scholar] [CrossRef]
- Qin, X.; Liu, C.; Zhou, Y.; Wang, G. Cisplatin induces programmed death-1-ligand 1(PD-L1) over-expression in hepatoma H22 cells via Erk /MAPK signaling pathway. Cell. Mol. Biol. 2010, 56, OL1366-72. [Google Scholar] [PubMed]
- Tanaka, K.; Miyata, H.; Sugimura, K.; Kanemura, T.; Hamada-Uematsu, M.; Mizote, Y.; Yamasaki, M.; Wada, H.; Nakajima, K.; Takiguchi, S.; et al. Negative influence of programmed death-1-ligands on the survival of esophageal cancer patients treated with chemotherapy. Cancer Sci. 2016, 107, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Dell’Agnola, C.; Biragyn, A. Clinical utilization of chemokines to combat cancer: The double-edged sword. Expert Rev. Vaccines 2007, 6, 267–283. [Google Scholar] [CrossRef]
- Fridman, W.H.; Pages, F.; Sautes-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef]
- Schiavoni, G.; Gabriele, L.; Mattei, F. The tumor microenvironment: A pitch for multiple players. Front. Oncol. 2013, 3, 90. [Google Scholar] [CrossRef]
- Shurin, M.R.; Shurin, G.V.; Lokshin, A.; Yurkovetsky, Z.R.; Gutkin, D.W.; Chatta, G.; Zhong, H.; Han, B.; Ferris, R.L. Intratumoral cytokines/chemokines/growth factors and tumor infiltrating dendritic cells: Friends or enemies? Cancer Metastasis Rev. 2006, 25, 333–356. [Google Scholar] [CrossRef]
- Whiteside, T.L. The role of immune cells in the tumor microenvironment. Cancer Treat. Res. 2006, 130, 103–124. [Google Scholar]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Chowdhury, R.; Gales, D.; Valenzuela, P.; Miller, S.; Yehualaeshet, T.; Manne, U.; Francia, G.; Samuel, T. Bromoethylindole (BEI-9) redirects NF-kappaB signaling induced by camptothecin and TNFalpha to promote cell death in colon cancer cells. Apoptosis Int. J. Program. Cell Death 2017, 22, 1553–1563. [Google Scholar] [CrossRef]
- Fadlalla, K.; Elgendy, R.; Gilbreath, E.; Pondugula, S.R.; Yehualaeshet, T.; Mansour, M.; Serbessa, T.; Manne, U.; Samuel, T. 3-(2-Bromoethyl)-indole inhibits the growth of cancer cells and NF-kappaB activation. Oncol. Rep. 2015, 34, 495–503. [Google Scholar] [CrossRef]
- Zhao, M.; Liang, F.; Zhang, B.; Yan, W.; Zhang, J. The impact of osteopontin on prognosis and clinicopathology of colorectal cancer patients: A systematic meta-analysis. Sci. Rep. 2015, 5, 12713. [Google Scholar] [CrossRef]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef]
- Panneerselvam, J.; Srivastava, A.; Muralidharan, R.; Wang, Q.; Zheng, W.; Zhao, L.; Chen, A.; Zhao, Y.D.; Munshi, A.; Ramesh, R. IL-24 modulates the high mobility group (HMG) A1/miR222 /AKT signaling in lung cancer cells. Oncotarget 2016, 7, 70247–70263. [Google Scholar] [CrossRef] [PubMed]
- Angiolillo, A.L.; Sgadari, C.; Taub, D.D.; Liao, F.; Farber, J.M.; Maheshwari, S.; Kleinman, H.K.; Reaman, G.H.; Tosato, G. Human interferon-inducible protein 10 is a potent inhibitor of angiogenesis in vivo. J. Exp. Med. 1995, 182, 155–162. [Google Scholar] [CrossRef]
- Nardinocchi, L.; Sonego, G.; Passarelli, F.; Avitabile, S.; Scarponi, C.; Failla, C.M.; Simoni, S.; Albanesi, C.; Cavani, A. Interleukin-17 and interleukin-22 promote tumor progression in human nonmelanoma skin cancer. Eur. J. Immunol. 2015, 45, 922–931. [Google Scholar] [CrossRef]
- Huber, S.; Gagliani, N.; Zenewicz, L.A.; Huber, F.J.; Bosurgi, L.; Hu, B.; Hedl, M.; Zhang, W.; O’Connor, W., Jr.; Murphy, A.J.; et al. IL-22BP is regulated by the inflammasome and modulates tumorigenesis in the intestine. Nature 2012, 491, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Simson, L.; Ellyard, J.I.; Dent, L.A.; Matthaei, K.I.; Rothenberg, M.E.; Foster, P.S.; Smyth, M.J.; Parish, C.R. Regulation of carcinogenesis by IL-5 and CCL11: A potential role for eosinophils in tumor immune surveillance. J. Immunol. 2007, 178, 4222–4229. [Google Scholar] [CrossRef]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Du, W.; Chen, Z.; Xiang, C. Upregulation of PD-L1 by SPP1 mediates macrophage polarization and facilitates immune escape in lung adenocarcinoma. Exp. Cell Res. 2017, 359, 449–457. [Google Scholar] [CrossRef]
- Bocci, G.; Falcone, A.; Fioravanti, A.; Orlandi, P.; Di Paolo, A.; Fanelli, G.; Viacava, P.; Naccarato, A.G.; Kerbel, R.S.; Danesi, R.; et al. Antiangiogenic and anticolorectal cancer effects of metronomic irinotecan chemotherapy alone and in combination with semaxinib. Br. J. Cancer 2008, 98, 1619–1629. [Google Scholar] [CrossRef]
- Allegrini, G.; Falcone, A.; Fioravanti, A.; Barletta, M.T.; Orlandi, P.; Loupakis, F.; Cerri, E.; Masi, G.; Di Paolo, A.; Kerbel, R.S.; et al. A pharmacokinetic and pharmacodynamic study on metronomic irinotecan in metastatic colorectal cancer patients. Br. J. Cancer 2008, 98, 1312–1319. [Google Scholar] [CrossRef]
- Kerbel, R.S.; Shaked, Y. The potential clinical promise of ‘multimodality’ metronomic chemotherapy revealed by preclinical studies of metastatic disease. Cancer Lett. 2017, 400, 293–304. [Google Scholar] [CrossRef]
- Fioravanti, A.; Canu, B.; Ali, G.; Orlandi, P.; Allegrini, G.; Di Desidero, T.; Emmenegger, U.; Fontanini, G.; Danesi, R.; Del Tacca, M.; et al. Metronomic 5-fluorouracil, oxaliplatin and irinotecan in colorectal cancer. Eur. J. Pharmacol. 2009, 619, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.A.; Minn, A.J. Combination Cancer Therapy with Immune Checkpoint Blockade: Mechanisms and Strategies. Immunity 2018, 48, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Niimi, A.; Yasuhara, T.; Permata, T.B.M.; Hagiwara, Y.; Isono, M.; Nuryadi, E.; Sekine, R.; Oike, T.; Kakoti, S.; et al. DNA double-strand break repair pathway regulates PD-L1 expression in cancer cells. Nat. Commun. 2017, 8, 1751. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Overall Upregulated by CPT Treatment (Fold Change vs. Control) | ||
|---|---|---|---|
| 1 µM | 5 µM | Major Functions Summary | |
| SPP1 | 47.2 | 32.6 | Secretes Phospho-Protein; osteoclast attachment; enhances IL-12 and reduces IL-10 production; induces PD-L1, promotes tumor [19] |
| IL-12B | 45.1 | 104.3 | Interleukin 12B; growth factor for activated T and NK-cells |
| CCL22 | 36.9 | 20 | C-C Motif Chemokine Ligand 22; chemotactic for activated T cells, monocytes, DC, and NK cells; attracts T-regs [20] |
| IL-24 | 34.4 | 42.4 | Interleukin 24; terminal differentiation of melanoma cell, may be pro-apoptotic; reduces tumor cell migration [21] |
| CXCL10 | 29.1 | 15 | C-X-C Motif Chemokine Ligand 10; attracts and stimulates T cells, monocytes and NK cells; antimicrobial; anti-angiogenic [22] |
| IL1RN | 27.9 | 27.4 | Interleukin 1 Receptor Antagonist; inhibits IL-1a and b binding to IL1R, antagonist for IL-1; anti-inflammatory |
| TNF | 26.9 | 75.3 | Tumor Necrosis Factor; inflammatory signaling, induces IL-12, impairs T-reg function |
| CSF3 | 24.7 | 15.1 | Colony Stimulating Factor 3; production, differentiation, and function of granulocytes |
| LTA | 16.3 | 82.4 | Lymphotoxin Alpha; inflammatory, immunostimulatory, antiviral; cytotoxic to tumor cells, binds TNF receptor (TNFR) |
| IL-17F | 12.7 | 74.5 | Interleukin 17F; stimulates peripheral blood mononuclear cells (PBMC), T-cells, and the production of IL-6, IL-8, CSF2, potentially anti-angiogenic; protumorigenic [23] |
| CCL2 | 10.9 | 2.9 | C-C Motif Chemokine Ligand 2; tactic for monocytes and basophils, enhances monocyte anti-tumor activity |
| CCL17 | 10.7 | 16.3 | C-C Motif Chemokine Ligand 17; tactic for T-cells, antimicrobial |
| CCL5 | 5.3 | 13.7 | C-C Motif Chemokine Ligand 5; tactic for monocytes, Th cells and eosinophils; suppresses HIV replication; aka RANTES |
| IL-22 | 5.2 | 35.5 | Interleukin 22; in vivo inflammatory reactions; protumorigenic in established cancer [24] |
| IL-5 | 4.3 | 24.6 | Interleukin 5; growth and differentiation of B cells and eosinophils; clusters on Chromosome 5 with IL-4, IL-13 and CSF2 (all anti-inflammatory); eosinophil chemotaxis [25] |
| IL-13 | 3.6 | 10.5 | Interleukin 13; regulation of B cell differentiation, anti-inflammatory, produced by Th2 cells, together with IL-10, IL-4, and IL-5 |
| IL-16 | 2 | 4.6 | Interleukin 16; stimulate migration of CD4 T cells, monocytes and eosinophils; primes CD4 cells for IL-2 and IL-15 |
| CCL21 | 1.8 | 8.5 | C-C Motif Chemokine Ligand 21; tactic for T-cells; homing of lymphocytes to secondary lymphoid organs |
| ADIPOQ | 1.6 | 4.8 | Adiponectin, C1Q And Collagen Domain Containing; anti-inflammatory adipokine, controls fat metabolism and insulin sensitivity, exclusive to adipose tissue |
| IL-1A | 1.6 | 3.6 | Interleukin 1A; inflammatory; thymocyte proliferation, B-cell maturation; secreted by Th1 and innate immune cells, with interferon (IFNγ), IL-6, tumor necrosis factor (TNFα) |
| OSM | 1.2 | 5.4 | Oncostatin-M; growth regulator, inhibits tumor cells; regulates IL-6, GM-CSF, and G-CSF production from endothelial cells |
| TNFSF13B | −1.4 | 3.2 | TNF superfamily 13B; Potent B-cell activator |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedi, D.; Henderson, H.J.; Manne, U.; Samuel, T. Camptothecin Induces PD-L1 and Immunomodulatory Cytokines in Colon Cancer Cells. Medicines 2019, 6, 51. https://doi.org/10.3390/medicines6020051
Bedi D, Henderson HJ, Manne U, Samuel T. Camptothecin Induces PD-L1 and Immunomodulatory Cytokines in Colon Cancer Cells. Medicines. 2019; 6(2):51. https://doi.org/10.3390/medicines6020051
Chicago/Turabian StyleBedi, Deepa, Henry J. Henderson, Upender Manne, and Temesgen Samuel. 2019. "Camptothecin Induces PD-L1 and Immunomodulatory Cytokines in Colon Cancer Cells" Medicines 6, no. 2: 51. https://doi.org/10.3390/medicines6020051
APA StyleBedi, D., Henderson, H. J., Manne, U., & Samuel, T. (2019). Camptothecin Induces PD-L1 and Immunomodulatory Cytokines in Colon Cancer Cells. Medicines, 6(2), 51. https://doi.org/10.3390/medicines6020051