
Unilateral Mitochondrial–Hemodynamic Coupling and Bilateral Connectivity in the Prefrontal Cortices of Young and Older Healthy Adults

 ,
,  , and
, and
Abstract
:1. Introduction
1.1. Unilateral Metabolic–Hemodynamic Coupling and Bilateral Connectivity
1.2. Three Infraslow Oscillation Bands in Cerebral CCO and HbO Signals
1.3. Aim of This Study
2. Materials and Methods
2.1. Participants
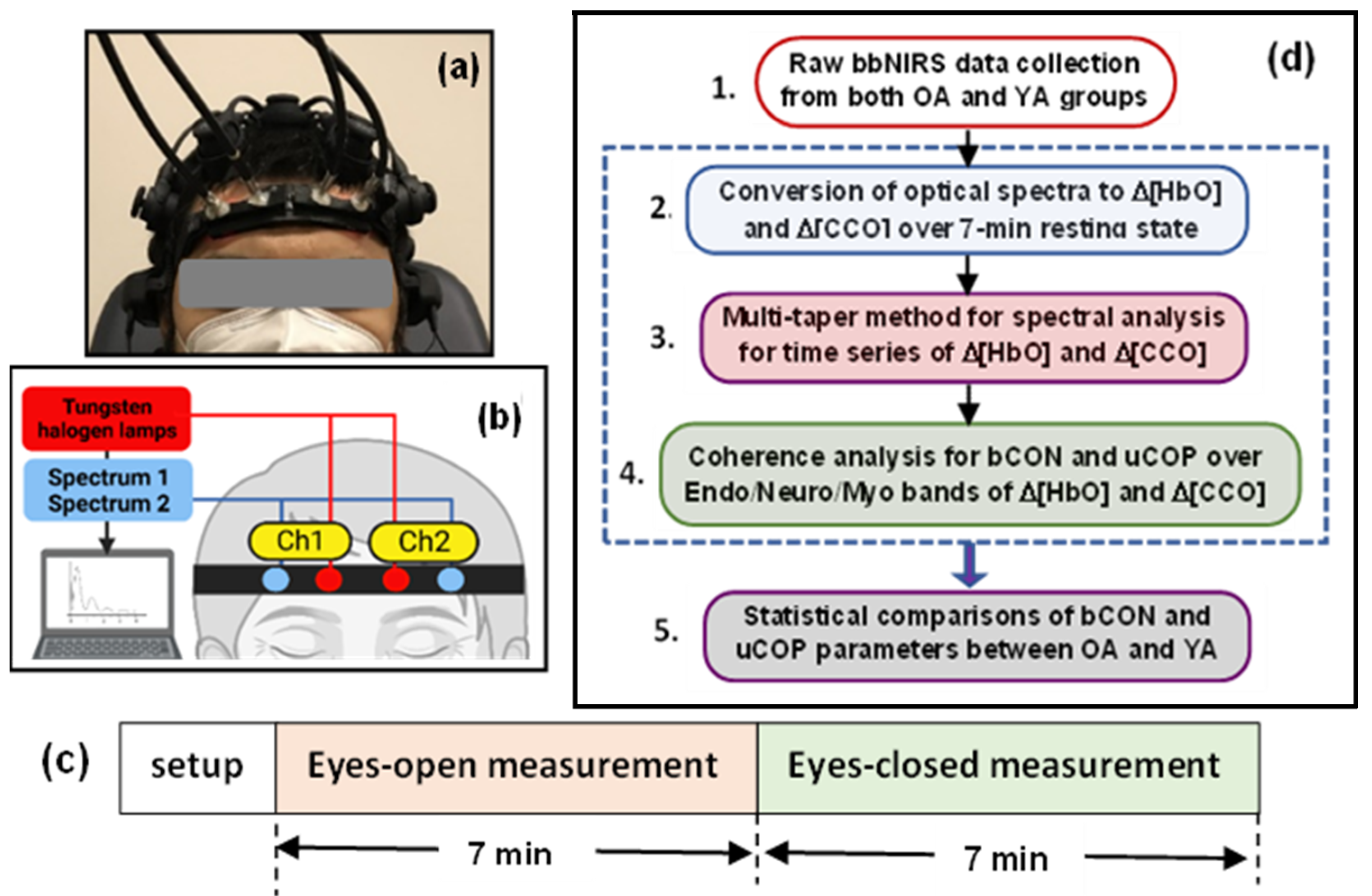
2.2. Experiment Protocol and Setup
2.3. Broadband Near-Infrared Spectroscopy and Its Measurements
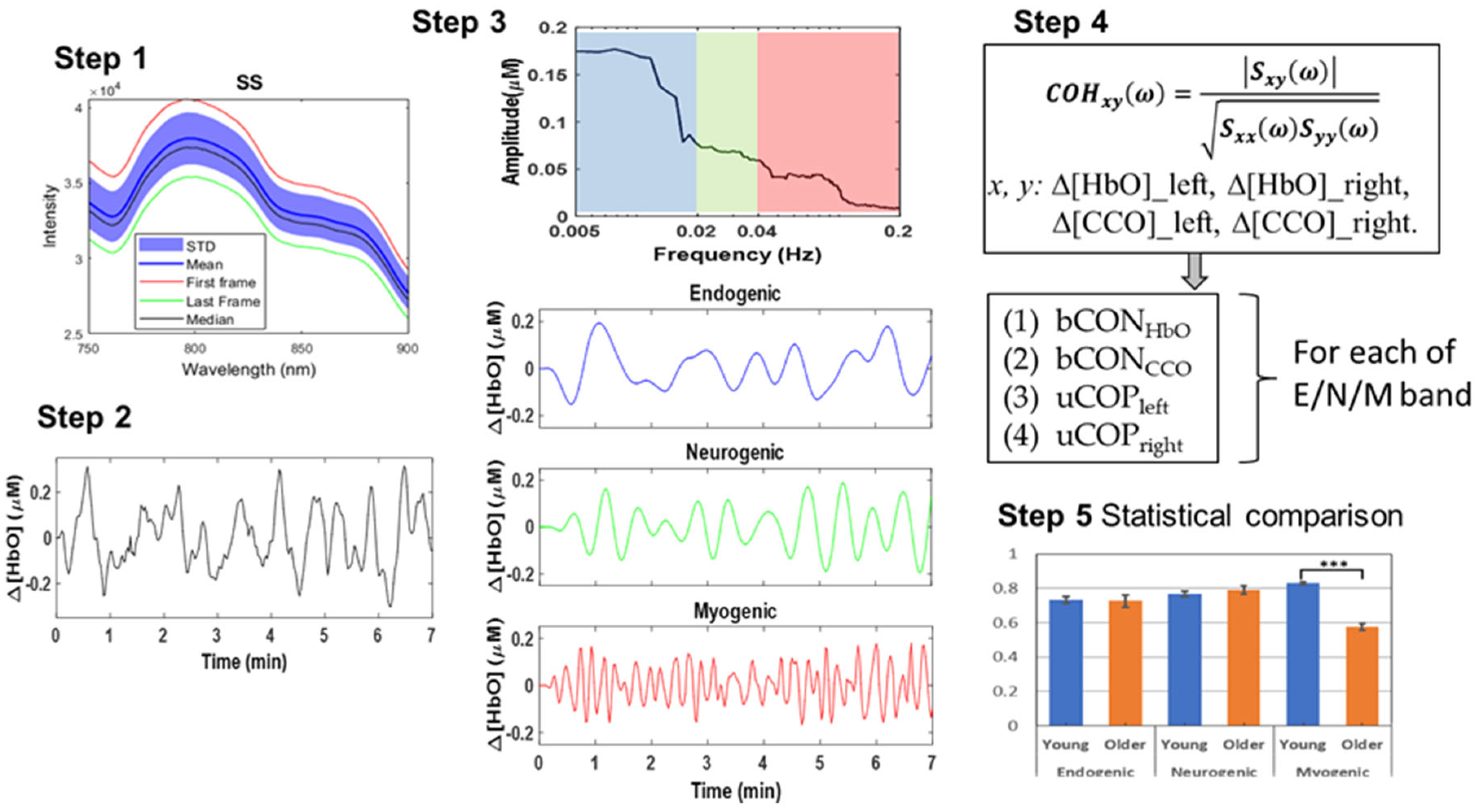
2.4. Data Analysis
- Step 1: Raw bbNIRS data collection from both older and young adult groups
- Step 2: Conversion of ΔOD(t, λ) to Δ[HbO](t, λ) and Δ[CCO](t, λ) over the 7-min resting state
- Step 3: Spectral analysis of Δ[HbO](t, λ) and Δ[CCO](t, λ)
- Step 5: Statistical Analysis
3. Results
3.1. Between-Group Comparisons of Bilateral Prefrontal Connectivity (bCON)
3.2. Between-Group Comparisons of Unilateral Prefrontal Coupling (uCOP)
3.3. Comparisons of bCON and uCOP Metrics under Eyes-Open and Eyes-Closed Conditions in Older Adults
3.4. Gender Comparisons of bCON and uCOP Metrics in Young Adults
4. Discussion
4.1. Age Effect on Bilateral Hemodynamic Connectivity of the Resting Prefrontal Cortex
4.2. Age Effect on Bilateral Metabolic Connectivity of the Resting Prefrontal Cortex
4.3. Age Effect on Unilateral Metabolic–Hemodynamic Coupling of the Resting Prefrontal Cortex
4.4. Signals Measured under Eyes-Open and Eyes-Closed Conditions
4.5. Gender Difference in Resting Bilateral Connectivity and Unilateral Coupling
4.6. Discussion on Using t-Tests to Compare Young and Older Adults
4.7. Limitations of the Study and Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| bbNIRS | broadband near-infrared spectroscopy |
| CCO | redox state of cytochrome c oxidase |
| Δ[CCO] | changes of redox state CCO concentration |
| HbO | oxygenated hemoglobin |
| Δ[HbO] | changes of HbO concentration |
| bCON | bilateral connectivity |
| bCONHbO | bilateral connectivity of HbO |
| bCONCCO | bilateral connectivity of redox state of CCO |
| uCOP | unilateral coupling |
| uCOPleft | unilateral coupling on the left prefrontal cortex |
| uCOPright | unilateral coupling on the right prefrontal cortex |
| E/M/N | endogenic/myogenic/neurogenic |
References
- Motzkin, J.C.; Newman, J.P.; Kiehl, K.A.; Koenigs, M. Reduced prefrontal connectivity in psychopathy. J. Neurosci. 2011, 31, 17348–17357. [Google Scholar] [CrossRef]
- Racz, F.S.; Mukli, P.; Nagy, Z.; Eke, A. Increased prefrontal cortex connectivity during cognitive challenge assessed by fNIRS imaging. Biomed. Opt. Express 2017, 8, 3842–3855. [Google Scholar] [CrossRef]
- Yu, J.W.; Lim, S.H.; Kim, B.; Kim, E.; Kim, K.; Kyu Park, S.; Seok Byun, Y.; Sakong, J.; Choi, J.W. Prefrontal functional connectivity analysis of cognitive decline for early diagnosis of mild cognitive impairment: A functional near-infrared spectroscopy study. Biomed. Opt. Express 2020, 11, 1725–1741. [Google Scholar] [CrossRef]
- Jobson, D.D.; Hase, Y.; Clarkson, A.N.; Kalaria, R.N. The role of the medial prefrontal cortex in cognition, ageing and dementia. Brain Commun. 2021, 3, fcab125. [Google Scholar] [CrossRef] [PubMed]
- Sampath, D.; Sathyanesan, M.; Newton, S.S. Cognitive dysfunction in major depression and Alzheimer’s disease is associated with hippocampal–prefrontal cortex dysconnectivity. Neuropsychiatr. Dis. Treat. 2017, 13, 1509. [Google Scholar] [CrossRef] [PubMed]
- Grady, C. The cognitive neuroscience of ageing. Nat. Rev. Neurosci. 2012, 13, 491–505. [Google Scholar] [CrossRef]
- Jones, L.K. Neurophysiological Development across the Life Span; American Counseling Association: Alexandria, VA, USA, 2017; pp. 27–44. [Google Scholar]
- Shahdadian, S.; Wang, X.; Kang, S.; Carter, C.; Chaudhari, A.; Liu, H. Prefrontal cortical connectivity and coupling of infraslow oscillation in the resting human brain: A 2-channel broadband NIRS study. Cereb. Cortex Commun. 2022, 3, tgac033. [Google Scholar] [CrossRef]
- Swami, S. Executive functions and decision making: A managerial review. IIMB Manag. Rev. 2013, 25, 203–212. [Google Scholar] [CrossRef]
- Wang, X.; Tian, F.; Reddy, D.D.; Nalawade, S.S.; Barrett, D.W.; Gonzalez-Lima, F.; Liu, H. Up-regulation of cerebral cytochrome-c-oxidase and hemodynamics by transcranial infrared laser stimulation: A broadband near-infrared spectroscopy study. J. Cereb. Blood Flow Metab. 2017, 37, 3789–3802. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.C.; Gonzalez-Lima, F. Low-level light therapy of the eye and brain. Eye Brain 2011, 3, 49–67. [Google Scholar] [CrossRef]
- Obrig, H.; Wenzel, R.; Kohl, M.; Horst, S.; Wobst, P.; Steinbrink, J.; Thomas, F.; Villringer, A. Near-infrared spectroscopy: Does it function in functional activation studies of the adult brain? Int. J. Psychophysiol. 2000, 35, 125–142. [Google Scholar] [CrossRef]
- Stefanovska, A.; Bracic, M.; Kvernmo, H.D. Wavelet analysis of oscillations in the peripheral blood circulation measured by laser Doppler technique. IEEE Trans. Biomed. Eng. 1999, 46, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Bracic, M.; Stefanovska, A. Wavelet-based analysis of human blood-flow dynamics. Bull. Math. Biol. 1998, 60, 919–935. [Google Scholar] [CrossRef] [PubMed]
- Vermeij, A.; Meel-van den Abeelen, A.S.; Kessels, R.P.; van Beek, A.H.; Claassen, J.A. Very-low-frequency oscillations of cerebral hemodynamics and blood pressure are affected by aging and cognitive load. Neuroimage 2014, 85 Pt 1, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Bernjak, A.; Clarkson, P.B.; McClintock, P.V.; Stefanovska, A. Low-frequency blood flow oscillations in congestive heart failure and after beta1-blockade treatment. Microvasc. Res. 2008, 76, 224–232. [Google Scholar] [CrossRef]
- Hillman, E.M.C. Coupling Mechanism and Significance of the BOLD Signal: A Status Report. Annu. Rev. Neurosci. 2014, 37, 161–181. [Google Scholar] [CrossRef]
- Kvernmo, H.D.; Stefanovska, A.; Kirkeboen, K.A.; Kvernebo, K. Oscillations in the human cutaneous blood perfusion signal modified by endothelium-dependent and endothelium-independent vasodilators. Microvasc. Res. 1999, 57, 298–309. [Google Scholar] [CrossRef]
- Wang, X.; Ma, L.C.; Shahdadian, S.; Wu, A.; Truong, N.C.D.; Liu, H. Metabolic Connectivity and Hemodynamic-Metabolic Coherence of Human Prefrontal Cortex at Rest and Post Photobiomodulation Assessed by Dual-Channel Broadband NIRS. Metabolites 2022, 12, 42. [Google Scholar] [CrossRef]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef]
- Shahdadian, S.; Wang, X.; Kang, S.; Carter, C.; Liu, H. Site-specific effects of 800- and 850-nm forehead transcranial photobiomodulation on prefrontal bilateral connectivity and unilateral coupling in young adults. Neurophotonics 2023, 10, 025012. [Google Scholar] [CrossRef]
- Vita, J.A.; Keaney, J.F., Jr. Endothelial function: A barometer for cardiovascular risk? Circulation 2002, 106, 640–642. [Google Scholar] [CrossRef]
- Deanfield, J.E.; Halcox, J.P.; Rabelink, T.J. Endothelial function and dysfunction: Testing and clinical relevance. Circulation 2007, 115, 1285–1295. [Google Scholar] [CrossRef]
- Di Marco, L.Y.; Farkas, E.; Martin, C.; Venneri, A.; Frangi, A.F. Is vasomotion in cerebral arteries impaired in Alzheimer’s disease? J. Alzheimer’s Dis. 2015, 46, 35–53. [Google Scholar] [CrossRef] [PubMed]
- West, R.L. An application of prefrontal cortex function theory to cognitive aging. Psychol. Bull. 1996, 120, 272–292. [Google Scholar] [CrossRef] [PubMed]
- Cazzell, M.; Li, L.; Lin, Z.J.; Patel, S.J.; Liu, H. Comparison of neural correlates of risk decision making between genders: An exploratory fNIRS study of the Balloon Analogue Risk Task (BART). NeuroImage 2012, 62, 1896–1911. [Google Scholar] [CrossRef]
- Li, L.; Cazzell, M.; Zeng, L.; Liu, H. Are there gender differences in young vs. aging brains under risk decision-making? An optical brain imaging study. Brain Imaging Behav. 2017, 11, 1085–1098. [Google Scholar] [CrossRef]
- de Roever, I.; Bale, G.; Cooper, R.J.; Tachtsidis, I. Functional NIRS Measurement of Cytochrome-C-Oxidase Demonstrates a More Brain-Specific Marker of Frontal Lobe Activation Compared to the Haemoglobins. Adv. Exp. Med. Biol. 2017, 977, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, T.; Carter, C.; Wang, X.; Wu, A.; Liu, H. Photobiomodulation at Different Wavelengths Boosts Mitochondrial Redox Metabolism and Hemoglobin Oxygenation: Lasers vs. Light-Emitting Diodes In Vivo. Metabolites 2022, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, T.; Wang, X.; Wu, A.; Kallioniemi, E.; Husain, M.M.; Liu, H. Transcranial Photobiomodulation (tPBM) With 1064-nm Laser to Improve Cerebral Metabolism of the Human Brain In Vivo. Lasers Surg. Med. 2020, 52, 807–813. [Google Scholar] [CrossRef]
- Truong, N.C.D.; Shahdadian, S.; Kang, S.; Wang, X.; Liu, H. Influence of the Signal-To-Noise Ratio on Variance of Chromophore Concentration Quantification in Broadband Near-Infrared Spectroscopy. Front. Photonics 2022, 3, 908931. [Google Scholar] [CrossRef]
- Jacques, S.L. Optical properties of biological tissues: A review. Phys. Med. Biol. 2013, 58, R37–R61. [Google Scholar] [CrossRef]
- Kolyva, C.; Tachtsidis, I.; Ghosh, A.; Moroz, T.; Cooper, C.E.; Smith, M.; Elwell, C.E. Systematic investigation of changes in oxidized cerebral cytochrome c oxidase concentration during frontal lobe activation in healthy adults. Biomed. Opt. Express 2012, 3, 2550–2566. [Google Scholar] [CrossRef]
- Bainbridge, A.; Tachtsidis, I.; Faulkner, S.D.; Price, D.; Zhu, T.; Baer, E.; Broad, K.D.; Thomas, D.L.; Cady, E.B.; Robertson, N.J.; et al. Brain mitochondrial oxidative metabolism during and after cerebral hypoxia-ischemia studied by simultaneous phosphorus magnetic-resonance and broadband near-infrared spectroscopy. NeuroImage 2014, 102 Pt 1, 173–183. [Google Scholar] [CrossRef]
- Bale, G.; Mitra, S.; Meek, J.; Robertson, N.; Tachtsidis, I. A new broadband near-infrared spectroscopy system for in-vivo measurements of cerebral cytochrome-c-oxidase changes in neonatal brain injury. Biomed. Opt. Express 2014, 5, 3450–3466. [Google Scholar] [CrossRef]
- Park, J.; Lindberg, C.R.; Vernon III, F.L. Multitaper spectral analysis of high-frequency seismograms. J. Geophys. Res. Solid. Earth 1987, 92, 12675–12684. [Google Scholar] [CrossRef]
- Babadi, B.; Brown, E.N. A review of multitaper spectral analysis. IEEE Trans. Biomed. Eng. 2014, 61, 1555–1564. [Google Scholar] [CrossRef]
- Popov, T.; Oostenveld, R.; Schoffelen, J.M. FieldTrip Made Easy: An Analysis Protocol for Group Analysis of the Auditory Steady State Brain Response in Time, Frequency, and Space. Front. Neurosci. 2018, 12, 711. [Google Scholar] [CrossRef] [PubMed]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011, 2011, 156869. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.M.; Schoffelen, J.M. A Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational Pitfalls. Front. Syst. Neurosci. 2015, 9, 175. [Google Scholar] [CrossRef]
- Percival, D.B.; Walden, A.T. Spectral Analysis for Physical Applications; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Li, L.; Babawale, O.; Yennu, A.; Trowbridge, C.; Hulla, R.; Gatchel, R.J.; Liu, H. Whole-cortical graphical networks at wakeful rest in young and older adults revealed by functional near-infrared spectroscopy. Neurophotonics 2018, 5, 035004. [Google Scholar] [CrossRef]
- Devore, J.; Peck, R. Statistics; Duxbury Press: New York, NY, USA, 1997. [Google Scholar]
- Welch, P. The use of fast Fourier transform for the estimation of power spectra: A method based on time averaging over short, modified periodograms. IEEE Trans. Audio Electroacoust. 1967, 15, 70–73. [Google Scholar] [CrossRef]
- Xu, G.; Huo, C.; Yin, J.; Zhong, Y.; Sun, G.; Fan, Y.; Wang, D.; Li, Z. Test-retest reliability of fNIRS in resting-state cortical activity and brain network assessment in stroke patients. Biomed. Opt. Express 2023, 14, 4217–4236. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Comparison of bCONHbO between the Two Age Groups | ||
| Frequency Bands | p Values (t-Test) | Cohen’s d |
| Endogenic | 0.79 | N/A |
| Neurogenic | 0.59 | N/A |
| Myogenic | 1.1 × 10−12 *** | 2.35 |
| (b) Comparison of bCONCCO between the Two Age Groups | ||
| Frequency Bands | p Values (t-Test) | Cohen’s d |
| Endogenic | 0.023 * | 0.37 |
| Neurogenic | 0.163 | N/A |
| Myogenic | 1.1 × 10−21 *** | 2.99 |
| (a) Comparison of uCOPleft between the two age groups | ||
| Frequency Bands | p-Values (t-Test) | Cohen’s d |
| Endogenic | 0.011 * | 0.536 |
| Neurogenic | 0.0098 ** | 0.508 |
| Myogenic | 1.4 × 10−18 *** | 1.96 |
| (b) Comparison of uCOPright between the two age groups | ||
| Frequency Bands | p-Values (t-Test) | Cohen’s d |
| Endogenic | 0.023 * | 0.44 |
| Neurogenic | 0.0007 *** | 0.78 |
| Myogenic | 3.9 × 10−18 *** | 2.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sissons, C.; Saeed, F.; Carter, C.; Lee, K.; Kerr, K.; Shahdadian, S.; Liu, H. Unilateral Mitochondrial–Hemodynamic Coupling and Bilateral Connectivity in the Prefrontal Cortices of Young and Older Healthy Adults. Bioengineering 2023, 10, 1336. https://doi.org/10.3390/bioengineering10111336
Sissons C, Saeed F, Carter C, Lee K, Kerr K, Shahdadian S, Liu H. Unilateral Mitochondrial–Hemodynamic Coupling and Bilateral Connectivity in the Prefrontal Cortices of Young and Older Healthy Adults. Bioengineering. 2023; 10(11):1336. https://doi.org/10.3390/bioengineering10111336
Chicago/Turabian StyleSissons, Claire, Fiza Saeed, Caroline Carter, Kathy Lee, Kristen Kerr, Sadra Shahdadian, and Hanli Liu. 2023. "Unilateral Mitochondrial–Hemodynamic Coupling and Bilateral Connectivity in the Prefrontal Cortices of Young and Older Healthy Adults" Bioengineering 10, no. 11: 1336. https://doi.org/10.3390/bioengineering10111336
APA StyleSissons, C., Saeed, F., Carter, C., Lee, K., Kerr, K., Shahdadian, S., & Liu, H. (2023). Unilateral Mitochondrial–Hemodynamic Coupling and Bilateral Connectivity in the Prefrontal Cortices of Young and Older Healthy Adults. Bioengineering, 10(11), 1336. https://doi.org/10.3390/bioengineering10111336