
Using Micro-Electrode-Array Recordings and Retinal Disease Models to Elucidate Visual Functions: Simultaneous Recording of Local Electroretinograms and Ganglion Cell Action Potentials Reveals the Origin of Retinal Oscillatory Potentials

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation
2.3. Retinal Recordings
2.4. Light Stimulation
2.5. Pharmacology
2.6. Data Analysis and Statistics
3. Results
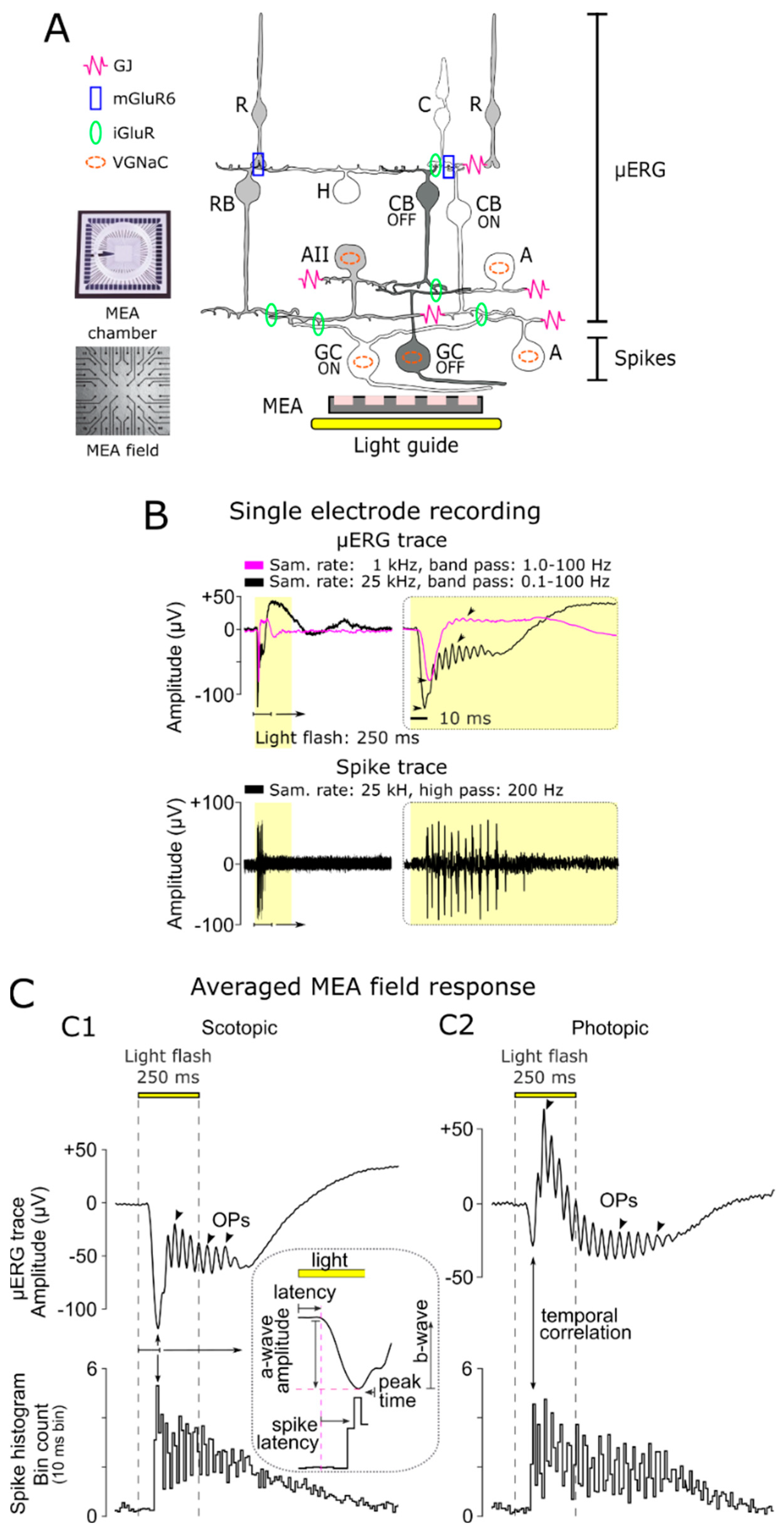
3.1. Characteristics of Light-Evoked Responses in Retinal Explants
3.2. Differentiation of Cellular Origin of Neuroretinal Signals
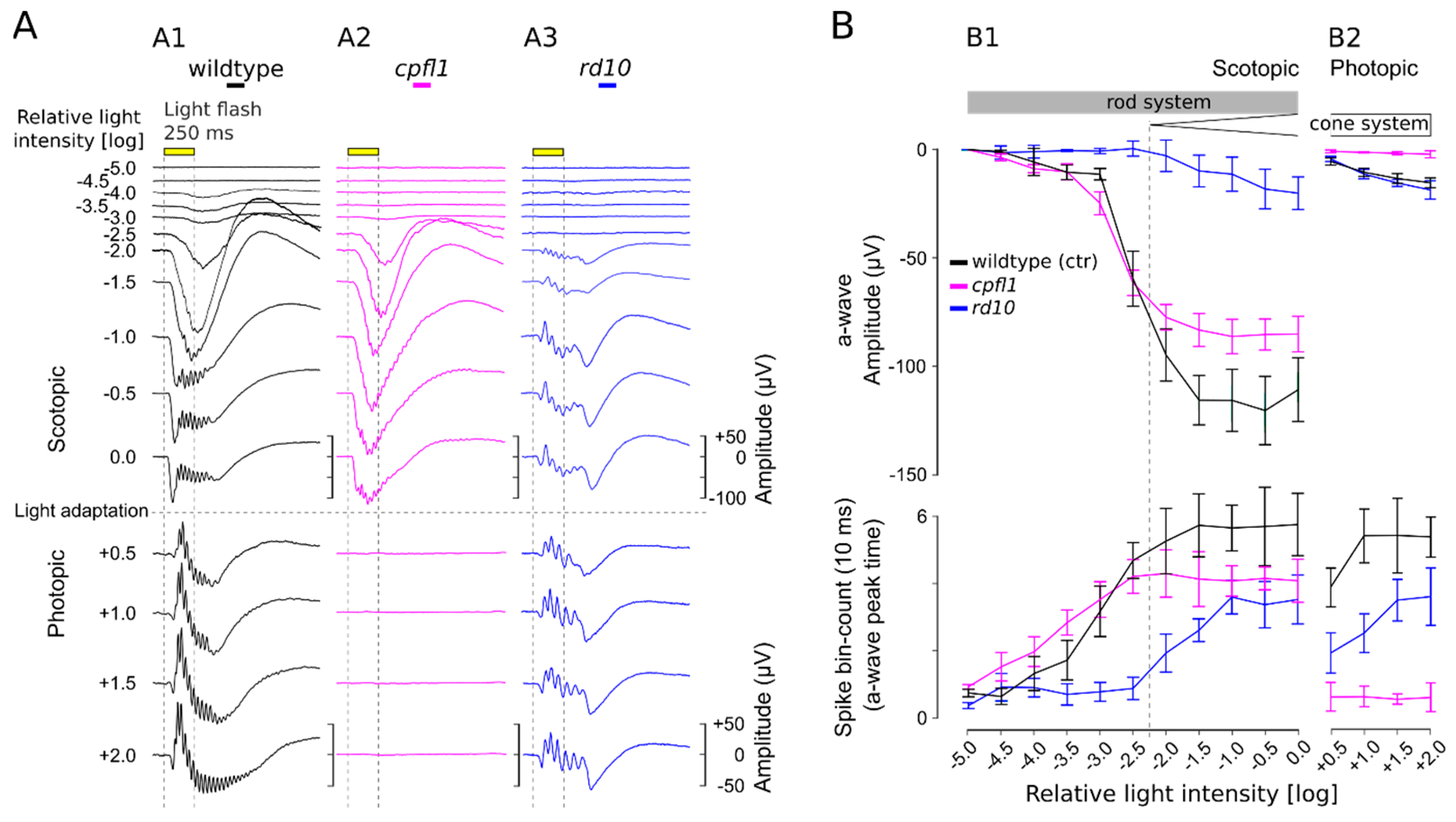
3.3. Scotopic and Photopic µERG Protocol for Retinal Explants
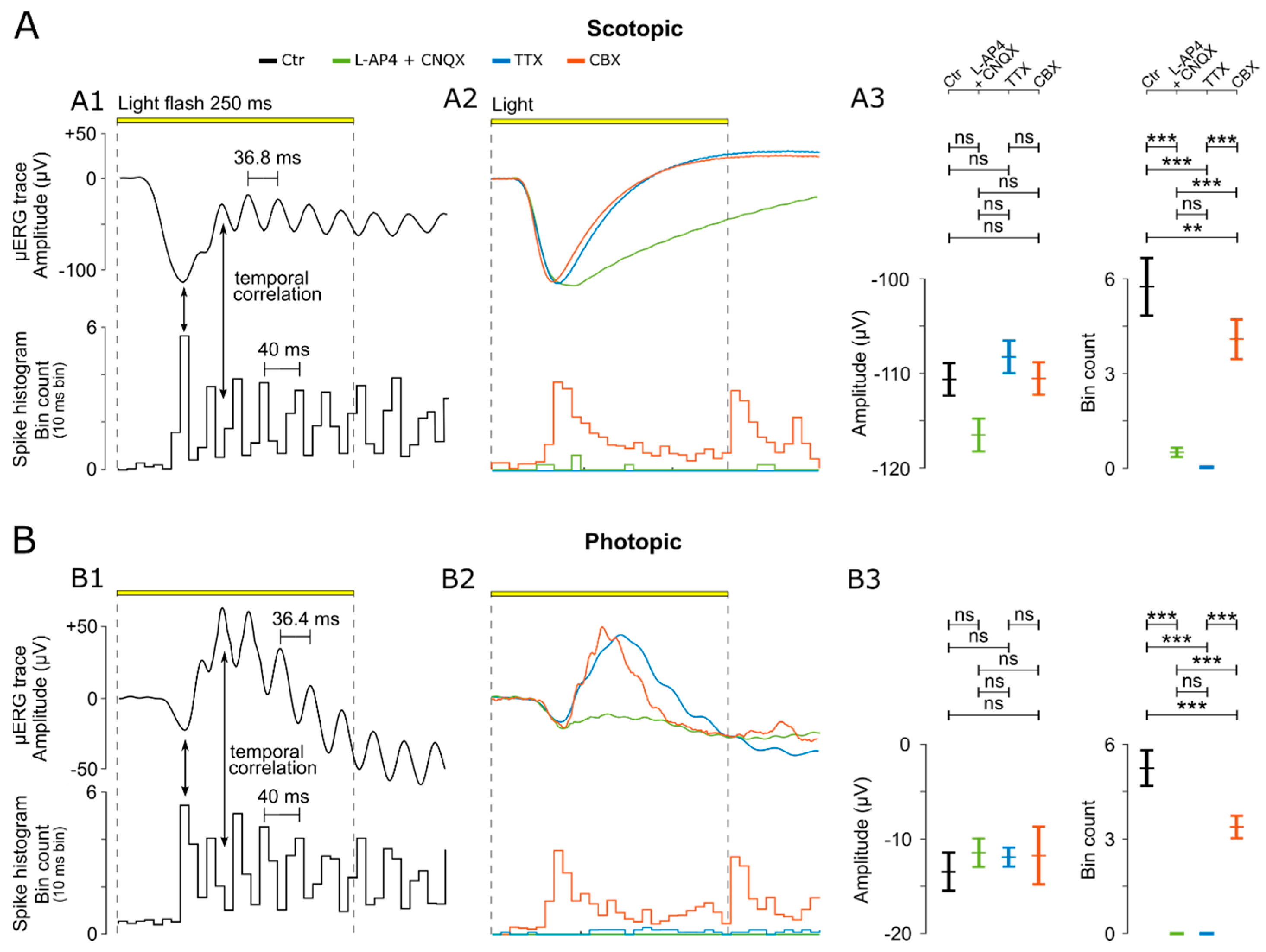
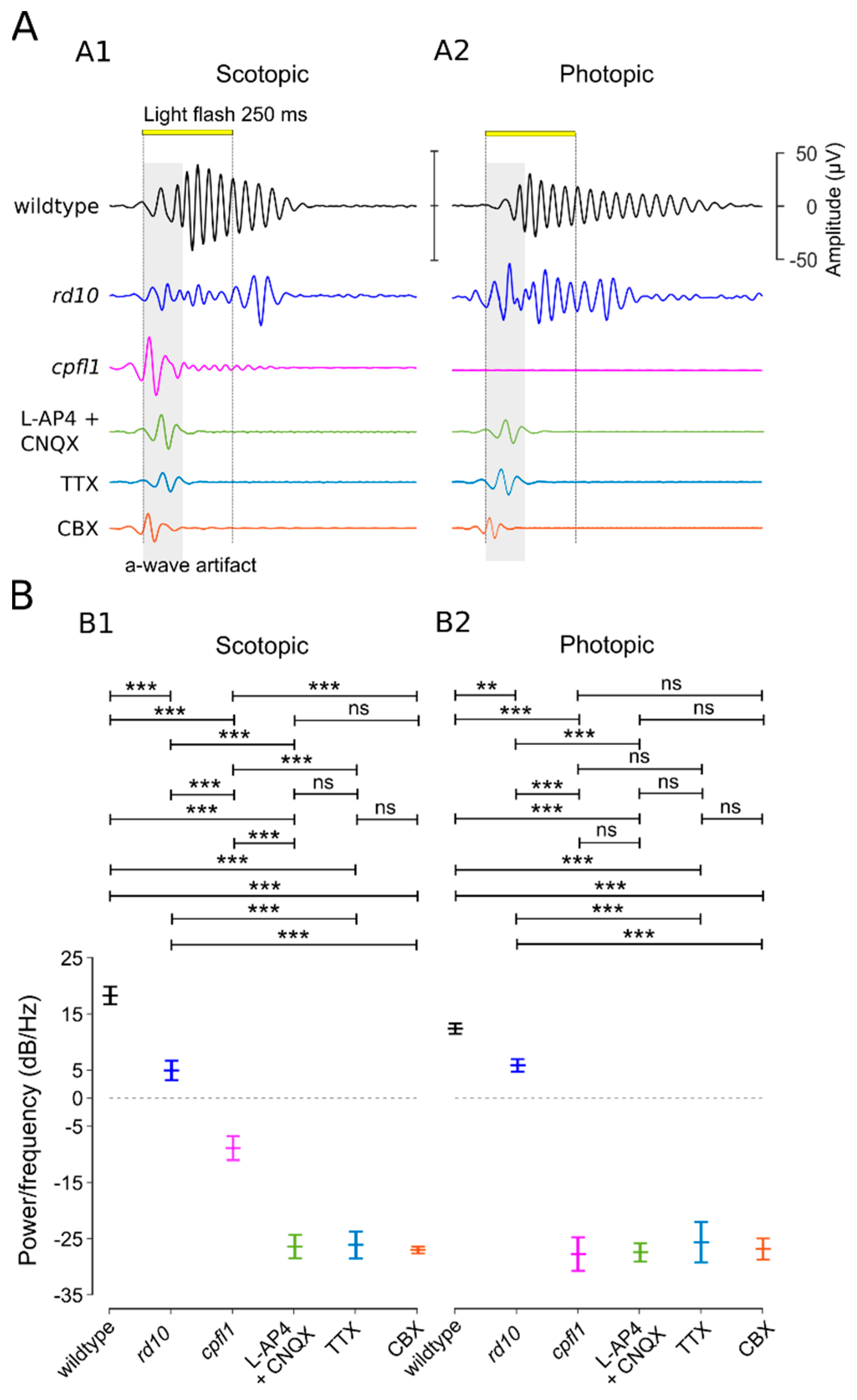
3.4. OPs Originate in the ACs and Synchronize GC Activity
4. Discussion
4.1. Ex Vivo µERG Recordings of Retinal Explants
4.2. Modulation of µERG Responses by Amacrine Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ERG | electroretinogram |
| µERG | micro-ERG |
| GC | ganglion cell |
| AC | amacrine cell |
| OP | oscillatory potentials |
| BC | bipolar cells |
| MEA | multi-electrode array |
| cpfl1 | B6.CXB1-Pde6ccpfl1/J |
| rd10 | C57BL/6J-Pde6brd10 |
| ACSF | artificial cerebrospinal fluid |
| mM | milli molar |
| L-AP4 | L-2-amino-4-phosphonobutyric acid |
| NBQX | 2,3-Dioxo-6-nitro-1,2,3,4-tetrahydro-benzo[f] quinoxaline-7-sulfonamide |
| TTX | tetrodotoxin |
| CBX | carbenoxolone |
| kHz | kilo hertz |
| GJ | gap junction |
| mGluR6 | metabotropic glutamate receptors |
| iGluR | inotropic glutamate receptors |
| VGNaC | voltage-gated sodium channels |
| R | rod |
| C | cone |
| RB | rod bipolar cell |
| H | horizontal cell |
| CB OFF | OFF-type cone bipolar cell |
| CB ON | ON-type cone bipolar cell |
| AII | amacrine cell-type AII |
| µV | microvolt |
| Hz | hertz |
| kHz | kilo hertz |
| Log | logarithm of base 10 |
| SEM | standard error mean |
| ns | not significant |
| ANOVA | analysis of variance |
| P | statistical significance value |
| ctr | control |
| dB | decibel |
| GABA | gamma-aminobutyric acid |
References
- Bearse, M.A., Jr. Principles and Practice of Clinical Electrophysiology of Vision, 2nd ed.; Optometry and Vision Science; MIT Press: Cambridge, MA, USA, 2007; Volume 84. [Google Scholar]
- Robson, A.G.; Nilsson, J.; Li, S.; Jalali, S.; Fulton, A.B.; Tormene, A.P.; Holder, G.E.; Brodie, S.E. ISCEV guide to visual electrodiagnostic procedures. Doc. Ophthalmol. 2018, 136, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, K.T. The electroretinogram: Its components and their origins. UCLA Forum Med. Sci. 1969, 8, 319–378. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.H.; Invergo, B.; Shimomura, K.; Takahashi, J.S.; Troy, J.B. Interpretation of the mouse electroretinogram. Doc. Ophthalmol. 2007, 115, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Frishman, L.J.; Wang, M.H. Electroretinogram of Human, Monkey and Mouse. In Adler’s Physiology of the Eye; Saunders: Philadelphia, PA, USA, 2011. [Google Scholar]
- Duncan, J.L.; LaVail, M.M.; Yasumura, D.; Matthes, M.T.; Yang, H.; Trautmann, N.; Chappelow, A.V.; Feng, W.; Earp, H.S.; Matsushima, G.K.; et al. An RCS-like retinal dystrophy phenotype in mer knockout mice. Investig. Ophthalmol. Vis. Sci. 2003, 44, 826–838. [Google Scholar] [CrossRef] [Green Version]
- Peachey, N.S.; Ball, S.L. Electrophysiological analysis of visual function in mutant mice. Doc. Ophthalmol. 2003, 107, 13–36. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, N.; Muehlfriedel, R.L.; Fischer, M.D.; Fahl, E.; Humphries, P.; Biel, M.; Seeliger, M.W. Vision tests in the mouse: Functional phenotyping with electroretinography. Front. Biosci. (Landmark Ed.) 2009, 14, 2730–2737. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.J.; Webb-Wood, S.; Faulkner, A.E.; Jabbar, S.B.; Biousse, V.; Newman, N.J.; Do, V.T.; Boatright, J.H.; Wallace, D.C.; Pardue, M.T. Retinal function and structure in Ant1-deficient mice. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6744–6752. [Google Scholar] [CrossRef]
- Schaeferhoff, K.; Michalakis, S.; Tanimoto, N.; Fischer, M.D.; Becirovic, E.; Beck, S.C.; Huber, G.; Rieger, N.; Riess, O.; Wissinger, B.; et al. Induction of STAT3-related genes in fast degenerating cone photoreceptors of cpfl1 mice. Cell. Mol. Life Sci. 2010, 67, 3173–3186. [Google Scholar] [CrossRef]
- Hare, W.A.; Ton, H. Effects of APB, PDA, and TTX on ERG responses recorded using both multifocal and conventional methods in monkey. Doc. Ophthalmol. 2002, 105, 189–222. [Google Scholar] [CrossRef]
- Zhao, C.; Yasumura, D.; Li, X.; Matthes, M.; Lloyd, M.; Nielsen, G.; Ahern, K.; Snyder, M.; Bok, D.; Dunaief, J.L.; et al. mTOR-mediated dedifferentiation of the retinal pigment epithelium initiates photoreceptor degeneration in mice. J. Clin. Investig. 2011, 121, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Robson, J.G.; Frishman, L.J. Photoreceptor and bipolar cell contributions to the cat electroretinogram: A kinetic model for the early part of the flash response. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 1996, 13, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Penn, R.D.; Hagins, W.A. Signal transmission along retinal rods and the origin of the electroretinographic a-wave. Nature 1969, 223, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Hagins, W.A.; Penn, R.D.; Yoshikami, S. Dark current and photocurrent in retinal rods. Biophys. J. 1970, 10, 380–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, D.C.; Birch, D.G. A quantitative measure of the electrical activity of human rod photoreceptors using electroretinography. Vis. Neurosci. 1990, 5, 379–387. [Google Scholar] [CrossRef]
- Goto, Y.; Peachey, N.S.; Ziroli, N.E.; Seiple, W.H.; Gryczan, C.; Pepperberg, D.R.; Naash, M.I. Rod phototransduction in transgenic mice expressing a mutant opsin gene. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 1996, 13, 577–585. [Google Scholar] [CrossRef]
- Lyubarsky, A.L.; Falsini, B.; Pennesi, M.E.; Valentini, P.; Pugh, E.N. UV- and Midwave-Sensitive Cone-Driven Retinal Responses of the Mouse: A Possible Phenotype for Coexpression of Cone Photopigments. J. Neurosci. 1999, 19, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, T.D.; Pugh, E.N., Jr. A quantitative account of the activation steps involved in phototransduction in amphibian photoreceptors. J. Physiol. 1992, 449, 719–758. [Google Scholar] [CrossRef]
- Pugh, E.N.; Falsini, B.; Lyubarsky, A.L. The Origin of the Major Rod- and Cone-Driven Components of the Rodent Electroretinogram and the Effect of Age and Light-Rearing History on the Magnitude of These Components. In Photostasis and Related Phenomena; Williams, T.P., Thistle, A.B., Eds.; Springer: Boston, MA, USA, 1998; pp. 93–128. [Google Scholar]
- Tanimoto, N.; Sothilingam, V.; Seeliger, M.W. Functional phenotyping of mouse models with ERG. Methods Mol. Biol. 2013, 935, 69–78. [Google Scholar] [CrossRef]
- Benchorin, G.; Calton, M.A.; Beaulieu, M.O.; Vollrath, D. Assessment of Murine Retinal Function by Electroretinography. Bio-Protocol 2017, 7, e2218. [Google Scholar] [CrossRef] [Green Version]
- Robson, J.G.; Frishman, L.J. Response linearity and kinetics of the cat retina: The bipolar cell component of the dark-adapted electroretinogram. Vis. Neurosci. 1995, 12, 837–850. [Google Scholar] [CrossRef]
- Tian, N.; Slaughter, M.M. Correlation of dynamic responses in the ON bipolar neuron and the b-wave of the electroretinogram. Vis. Res. 1995, 35, 1359–1364. [Google Scholar] [CrossRef] [Green Version]
- Robson, J.G.; Maeda, H.; Saszik, S.M.; Frishman, L.J. In vivo studies of signaling in rod pathways of the mouse using the electroretinogram. Vis. Res. 2004, 44, 3253–3268. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, R.H.; Linsenmeier, R.A.; Griff, E.R. Chapter 2 Retinal pigment epithelial cell contributions to the electroretinogram and electrooculogram. Prog. Retin. Res. 1985, 4, 33–66. [Google Scholar] [CrossRef]
- Hanitzsch, R.; Lichtenberger, T. Two neuronal retinal components of the electroretinogram c-wave. Doc. Ophthalmol. 1997, 94, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Samuels, I.S.; Sturgill, G.M.; Grossman, G.H.; Rayborn, M.E.; Hollyfield, J.G.; Peachey, N.S. Light-evoked responses of the retinal pigment epithelium: Changes accompanying photoreceptor loss in the mouse. J. Neurophysiol. 2010, 104, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachtmeister, L.; Dowling, J.E. The oscillatory potentials of the mudpuppy retina. Investig. Ophthalmol. Vis. Sci. 1978, 17, 1176–1188. [Google Scholar]
- Dong, C.J.; Agey, P.; Hare, W.A. Origins of the electroretinogram oscillatory potentials in the rabbit retina. Vis. Neurosci. 2004, 21, 533–543. [Google Scholar] [CrossRef]
- Mojumder, D.K.; Sherry, D.M.; Frishman, L.J. Contribution of voltage-gated sodium channels to the b-wave of the mammalian flash electroretinogram. J. Physiol. 2008, 586, 2551–2580. [Google Scholar] [CrossRef]
- Shirao, Y.; Kawasaki, K. Electrical responses from diabetic retina. Prog. Retin. Eye Res. 1998, 17, 59–76. [Google Scholar] [CrossRef]
- Dimopoulos, I.S.; Freund, P.R.; Redel, T.; Dornstauder, B.; Gilmour, G.; Sauve, Y. Changes in rod and cone-driven oscillatory potentials in the aging human retina. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5058–5073. [Google Scholar] [CrossRef] [Green Version]
- Midena, E.; Torresin, T.; Longhin, E.; Midena, G.; Pilotto, E.; Frizziero, L. Early Microvascular and Oscillatory Potentials Changes in Human Diabetic Retina: Amacrine Cells and the Intraretinal Neurovascular Crosstalk. J. Clin. Med. 2021, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Lachapelle, P.; Benoit, J.; Little, J.M.; Faubert, J. The diagnostic use of the second oscillatory potential in clinical electroretinography. Doc. Ophthalmol. 1989, 73, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Bui, B.V.; Edmunds, B.; Cioffi, G.A.; Fortune, B. The gradient of retinal functional changes during acute intraocular pressure elevation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 202–213. [Google Scholar] [CrossRef]
- Lachapelle, P. The human suprathreshold photopic oscillatory potentials: Method of analysis and clinical application. Doc. Ophthalmol. 1994, 88, 1–25. [Google Scholar] [CrossRef]
- Li, Q.; Zemel, E.; Miller, B.; Perlman, I. Early retinal damage in experimental diabetes: Electroretinographical and morphological observations. Exp. Eye Res. 2002, 74, 615–625. [Google Scholar] [CrossRef]
- Hancock, H.A.; Kraft, T.W. Oscillatory potential analysis and ERGs of normal and diabetic rats. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1002–1008. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cohen, E.D.; Qian, H. Rod and Cone Coupling Modulates Photopic ERG Responses in the Mouse Retina. Front. Cell Neurosci. 2020, 14, 566712. [Google Scholar] [CrossRef] [PubMed]
- Wachtmeister, L. Oscillatory potentials in the retina: What do they reveal. Prog. Retin. Eye Res. 1998, 17, 485–521. [Google Scholar] [CrossRef]
- Stett, A.; Egert, U.; Guenther, E.; Hofmann, F.; Meyer, T.; Nisch, W.; Haemmerle, H. Biological application of microelectrode arrays in drug discovery and basic research. Anal. Bioanal. Chem. 2003, 377, 486–495. [Google Scholar] [CrossRef]
- Green, D.G.; Kapousta-Bruneau, N.V. A dissection of the electroretinogram from the isolated rat retina with microelectrodes and drugs. Vis. Neurosci. 1999, 16, 727–741. [Google Scholar] [CrossRef]
- Fujii, M.; Sunagawa, G.A.; Kondo, M.; Takahashi, M.; Mandai, M. Evaluation of micro Electroretinograms Recorded with Multiple Electrode Array to Assess Focal Retinal Function. Sci. Rep. 2016, 6, 30719. [Google Scholar] [CrossRef]
- Jae, S.A.; Ahn, K.N.; Kim, J.Y.; Seo, J.H.; Kim, H.K.; Goo, Y.S. Electrophysiological and Histologic Evaluation of the Time Course of Retinal Degeneration in the rd10 Mouse Model of Retinitis Pigmentosa. Korean J. Physiol. Pharmacol. 2013, 17, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Dinculescu, A.; Dai, X.; Du, W.; Smith, W.C.; Pang, J. Review: The history and role of naturally occurring mouse models with Pde6b mutations. Mol. Vis. 2013, 19, 2579–2589. [Google Scholar]
- Chang, B.; Grau, T.; Dangel, S.; Hurd, R.; Jurklies, B.; Sener, E.C.; Andreasson, S.; Dollfus, H.; Baumann, B.; Bolz, S.; et al. A homologous genetic basis of the murine cpfl1 mutant and human achromatopsia linked to mutations in the PDE6C gene. Proc. Natl. Acad. Sci. USA 2009, 106, 19581–19586. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.; Hawes, N.L.; Hurd, R.E.; Davisson, M.T.; Nusinowitz, S.; Heckenlively, J.R. Retinal degeneration mutants in the mouse. Vis. Res. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, W.; Dietter, J.; Zrenner, E. Electrical activation of degenerated photoreceptors in blind mouse retina elicited network-mediated responses in different types of ganglion cells. Sci. Rep. 2018, 8, 16998. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Yao, G.; Zhang, K.; Hofeldt, K.J.; Chang, B. Study of rod- and cone-driven oscillatory potentials in mice. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2732–2738. [Google Scholar] [CrossRef] [Green Version]
- Carter-Dawson, L.D.; LaVail, M.M. Rods and cones in the mouse retina. I. Structural analysis using light and electron microscopy. J. Comp. Neurol. 1979, 188, 245–262. [Google Scholar] [CrossRef]
- Gargini, C.; Terzibasi, E.; Mazzoni, F.; Strettoi, E. Retinal organization in the retinal degeneration 10 (rd10) mutant mouse: A morphological and ERG study. J. Comp. Neurol. 2007, 500, 222–238. [Google Scholar] [CrossRef] [Green Version]
- Sieving, P.A.; Murayama, K.; Naarendorp, F. Push-pull model of the primate photopic electroretinogram: A role for hyperpolarizing neurons in shaping the b-wave. Vis. Neurosci. 1994, 11, 519–532. [Google Scholar] [CrossRef]
- Masland, R.H. The fundamental plan of the retina. Nat. Neurosci. 2001, 4, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Jarsky, T.; Murphy, G.J.; Rieke, F.; Singer, J.H. Voltage-gated Na channels in AII amacrine cells accelerate scotopic light responses mediated by the rod bipolar cell pathway. J. Neurosci. 2010, 30, 4650–4659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awatramani, G.; Wang, J.; Slaughter, M.M. Amacrine and ganglion cell contributions to the electroretinogram in amphibian retina. Vis. Neurosci. 2001, 18, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Vighi, E.; Trifunovic, D.; Veiga-Crespo, P.; Rentsch, A.; Hoffmann, D.; Sahaboglu, A.; Strasser, T.; Kulkarni, M.; Bertolotti, E.; van den Heuvel, A.; et al. Combination of cGMP analogue and drug delivery system provides functional protection in hereditary retinal degeneration. Proc. Natl. Acad. Sci. USA 2018, 115, E2997–E3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.; Rodgers, J.; Hickey, D.; Foster, R.G.; Peirson, S.N.; Hankins, M.W. Characterisation of light responses in the retina of mice lacking principle components of rod, cone and melanopsin phototransduction signalling pathways. Sci. Rep. 2016, 6, 28086. [Google Scholar] [CrossRef]
- Deans, M.R.; Volgyi, B.; Goodenough, D.A.; Bloomfield, S.A.; Paul, D.L. Connexin36 is essential for transmission of rod-mediated visual signals in the mammalian retina. Neuron 2002, 36, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Volgyi, B.; Deans, M.R.; Paul, D.L.; Bloomfield, S.A. Convergence and segregation of the multiple rod pathways in mammalian retina. J. Neurosci. 2004, 24, 11182–11192. [Google Scholar] [CrossRef] [Green Version]
- Fain, G.; Sampath, A.P. Rod and cone interactions in the retina. F1000Research 2018, 7, 657. [Google Scholar] [CrossRef]
- Liao, F.; Liu, H.; Milla-Navarro, S.; Villa, P.; Germain, F. Origin of Retinal Oscillatory Potentials in the Mouse, a Tool to Specifically Locate Retinal Damage. Int. J. Mol. Sci. 2023, 24, 3126. [Google Scholar] [CrossRef]
- Hood, D.C.; Frishman, L.J.; Viswanathan, S.; Robson, J.G.; Ahmed, J. Evidence for a ganglion cell contribution to the primate electroretinogram (ERG): Effects of TTX on the multifocal ERG in macaque. Vis. Neurosci. 1999, 16, 411–416. [Google Scholar] [CrossRef]
- Kaneko, Y.; Watanabe, S. Expression of Nav1.1 in rat retinal AII amacrine cells. Neurosci. Lett. 2007, 424, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Stasheff, S.F. Emergence of sustained spontaneous hyperactivity and temporary preservation of OFF responses in ganglion cells of the retinal degeneration (rd1) mouse. J. Neurophysiol. 2008, 99, 1408–1421. [Google Scholar] [CrossRef] [PubMed]
- Trenholm, S.; Borowska, J.; Zhang, J.; Hoggarth, A.; Johnson, K.; Barnes, S.; Lewis, T.J.; Awatramani, G.B. Intrinsic oscillatory activity arising within the electrically coupled AII amacrine-ON cone bipolar cell network is driven by voltage-gated Na+ channels. J. Physiol. 2012, 590, 2501–2517. [Google Scholar] [CrossRef] [PubMed]
- Menzler, J.; Zeck, G. Network oscillations in rod-degenerated mouse retinas. J. Neurosci. 2011, 31, 2280–2291. [Google Scholar] [CrossRef] [Green Version]
- Haq, W.; Arango-Gonzalez, B.; Zrenner, E.; Euler, T.; Schubert, T. Synaptic remodeling generates synchronous oscillations in the degenerated outer mouse retina. Front. Neural. Circuits 2014, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Achberger, K.; Probst, C.; Haderspeck, J.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging organoid and organ-on-a-chip technology to generate complex multi-layer tissue models in a human retina-on-a-chip platform. Elife 2019, 8, e46188. [Google Scholar] [CrossRef]
- Sun, N.; Meng, X.; Liu, Y.; Song, D.; Jiang, C.; Cai, J. Applications of brain organoids in neurodevelopment and neurological diseases. J. Biomed. Sci. 2021, 28, 30. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haq, W.; Zrenner, E.; Ueffing, M.; Paquet-Durand, F. Using Micro-Electrode-Array Recordings and Retinal Disease Models to Elucidate Visual Functions: Simultaneous Recording of Local Electroretinograms and Ganglion Cell Action Potentials Reveals the Origin of Retinal Oscillatory Potentials. Bioengineering 2023, 10, 725. https://doi.org/10.3390/bioengineering10060725
Haq W, Zrenner E, Ueffing M, Paquet-Durand F. Using Micro-Electrode-Array Recordings and Retinal Disease Models to Elucidate Visual Functions: Simultaneous Recording of Local Electroretinograms and Ganglion Cell Action Potentials Reveals the Origin of Retinal Oscillatory Potentials. Bioengineering. 2023; 10(6):725. https://doi.org/10.3390/bioengineering10060725
Chicago/Turabian StyleHaq, Wadood, Eberhart Zrenner, Marius Ueffing, and François Paquet-Durand. 2023. "Using Micro-Electrode-Array Recordings and Retinal Disease Models to Elucidate Visual Functions: Simultaneous Recording of Local Electroretinograms and Ganglion Cell Action Potentials Reveals the Origin of Retinal Oscillatory Potentials" Bioengineering 10, no. 6: 725. https://doi.org/10.3390/bioengineering10060725
APA StyleHaq, W., Zrenner, E., Ueffing, M., & Paquet-Durand, F. (2023). Using Micro-Electrode-Array Recordings and Retinal Disease Models to Elucidate Visual Functions: Simultaneous Recording of Local Electroretinograms and Ganglion Cell Action Potentials Reveals the Origin of Retinal Oscillatory Potentials. Bioengineering, 10(6), 725. https://doi.org/10.3390/bioengineering10060725