
Biomolecular Pathways of Cryoinjuries in Low-Temperature Storage for Mammalian Specimens


Abstract
:1. Introduction
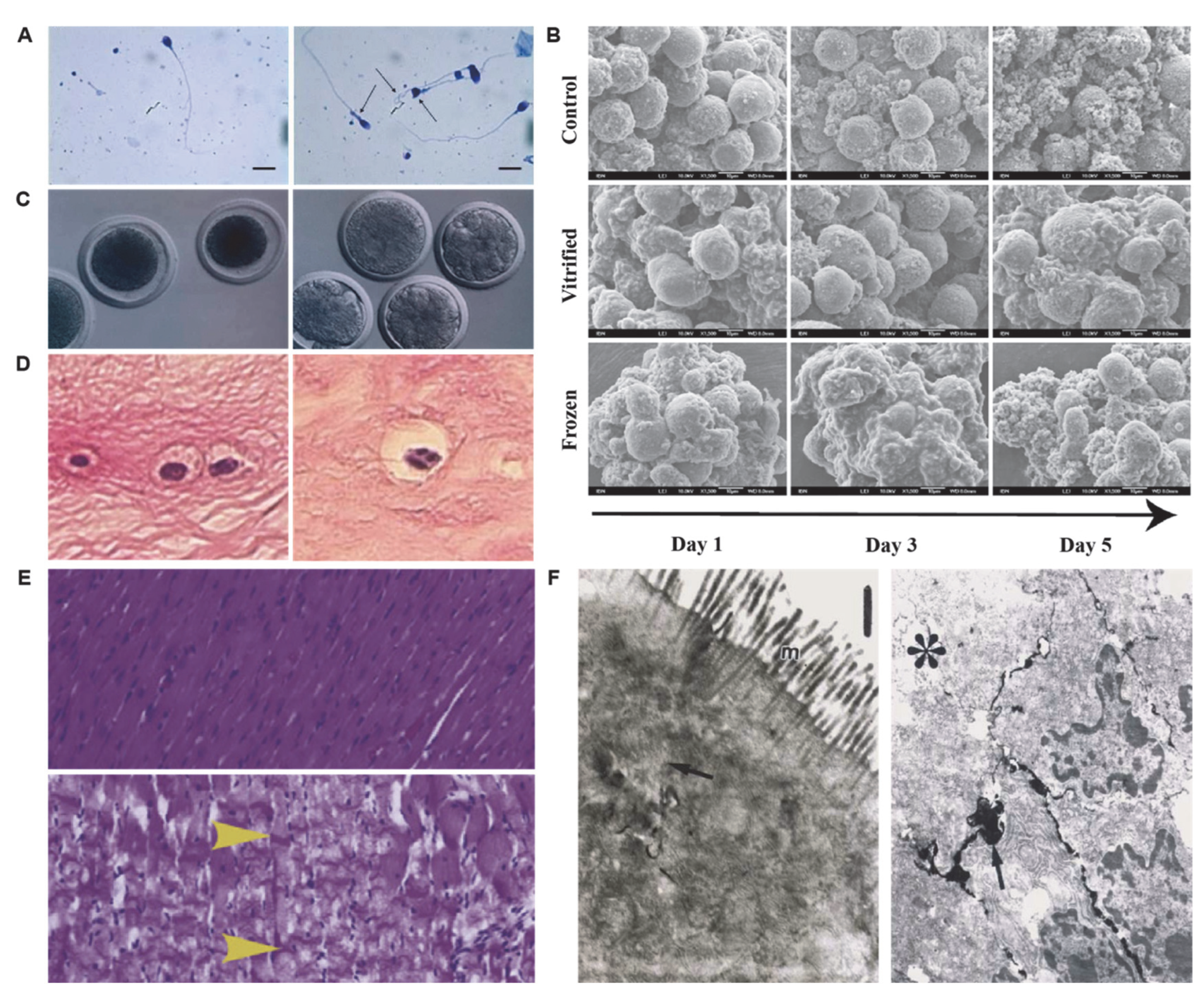
2. Morphological Changes in Preservation
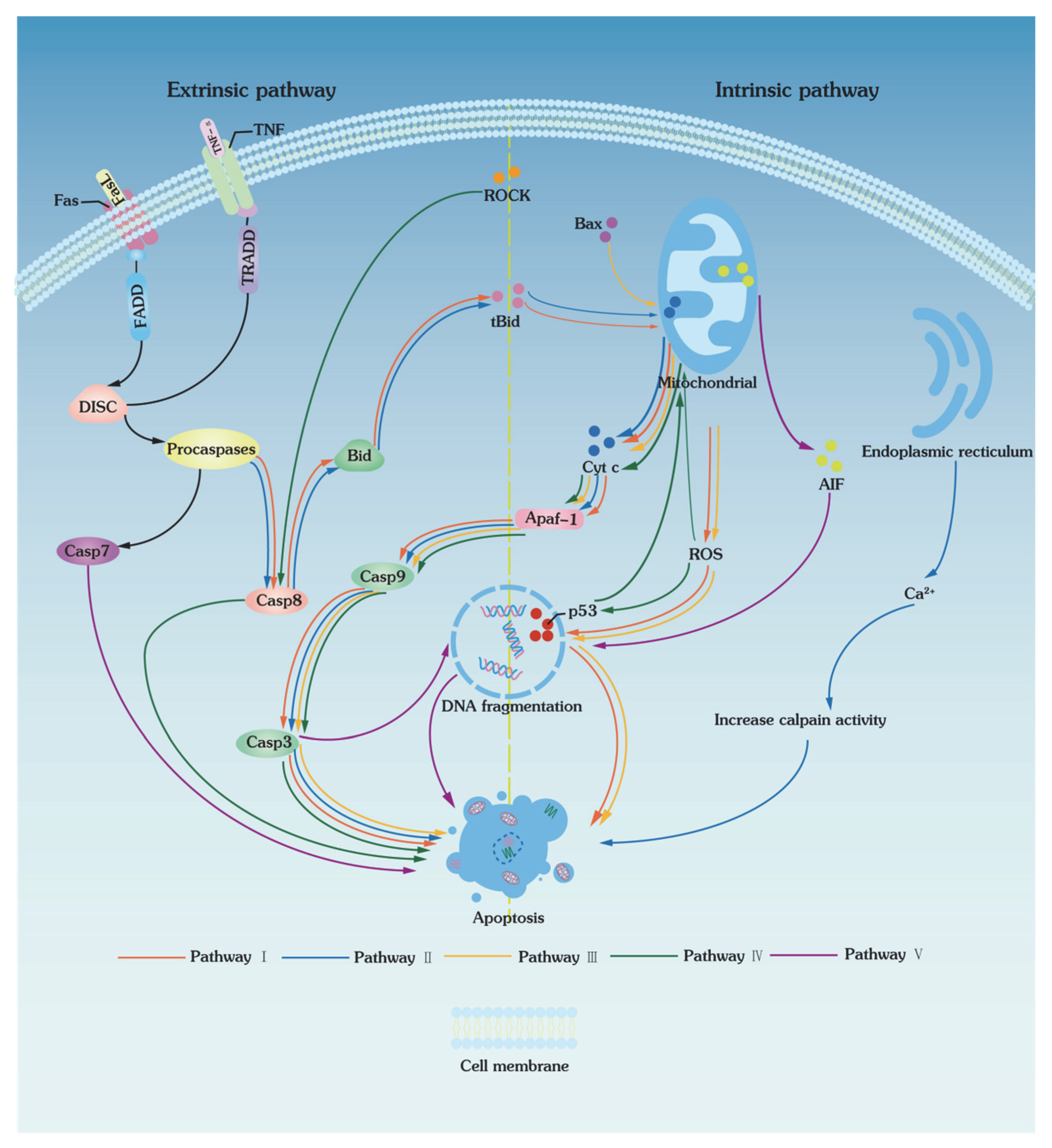
3. Apoptotic Pathways of Low-Temperature Biopreservation
3.1. Extrinsic Pathways
3.2. Intrinsic Pathways
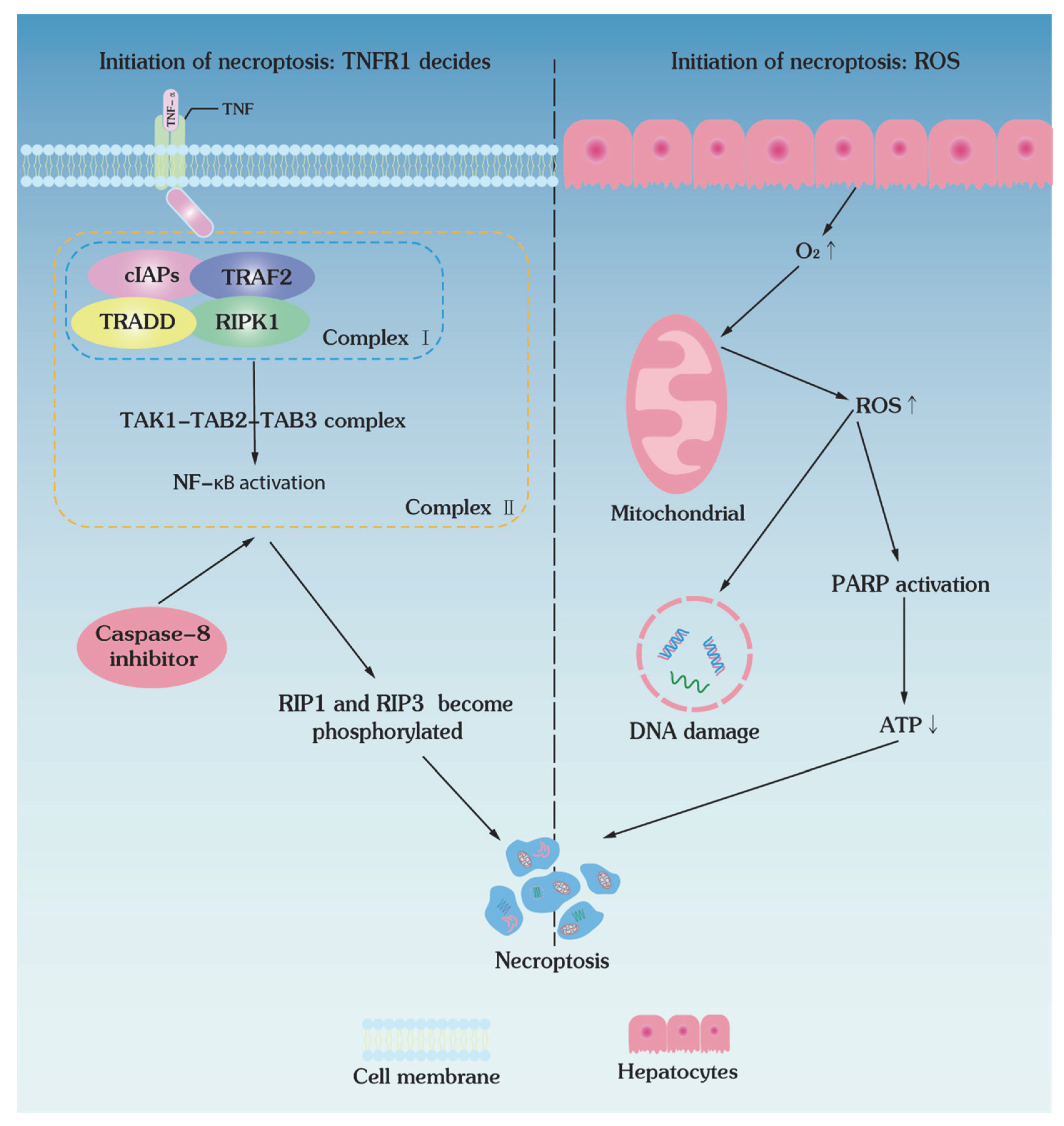
4. Necroptotic Pathways of Low-Temperature Biopreservation
4.1. Initiation of Necroptosis: TNFR1 Decides
4.2. Initiation of Necroptosis: ROS Decides
5. Molecular Pathways of Ischemia–Reperfusion Injuries
5.1. Ischemia Injury
5.2. Reperfusion Injury
6. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Vries, R.J.; Tessier, S.N.; Banik, P.D.; Nagpal, S.; Cronin, S.E.J.; Ozer, S.; Hafiz, E.O.A.; van Gulik, T.M.; Yarmush, M.L.; Markmann, J.F.; et al. Supercooling extends preservation time of human livers. Nat. Biotechnol. 2019, 37, 1131. [Google Scholar] [CrossRef]
- Zhan, L.; Rao, J.S.; Sethia, N.; Slama, M.Q.; Han, Z.; Tobolt, D.; Etheridge, M.; Peterson, Q.P.; Dutcher, C.S.; Bischof, J.C.; et al. Pancreatic islet cryopreservation by vitrification achieves high viability, function, recovery and clinical scalability for transplantation. Nat. Med. 2022, 28, 798. [Google Scholar] [CrossRef] [PubMed]
- Grau, S.; Martin-Garcia, E.; Ferrandez, O.; Martin, R.; Tejedor-Vaquero, S.; Gimeno, R.; Magri, G.; Maldonado, R. COVID-19 mRNA Vaccines Preserve Immunogenicity after Re-Freezing. Vaccines 2022, 10, 594. [Google Scholar] [CrossRef]
- Zhu, C.; Niu, Q.; Yuan, X.; Chong, J.; Ren, L. NonFreezable Preservation of Human Red Blood Cells at −8 °C. ACS Biomater. Sci. Eng. 2022, 8, 2644–2653. [Google Scholar] [CrossRef]
- Dou, M.; Lu, C.; Rao, W. Bioinspired materials and technology for advanced cryopreservation. Trends Biotechnol. 2022, 40, 93–106. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, L.; Liu, W.; Shen, J.; Sun, H.; Liang, J.; Lv, G.; Chen, G.; Yang, Y.; Ou, J. 5-Aminolevulinate improves metabolic recovery and cell survival of the liver following cold preservation. Theranostics 2022, 12, 2908–2927. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Ekpo, M.D.; Xiao, J.; Zhao, H.; Bai, X.; Liang, Y.; Zhao, G.; Liu, D.; Tan, S. Principles and Protocols for Post-Cryopreservation Quality Evaluation of Stem Cells in Novel Biomedicine. Front. Pharmacol. 2022, 13, 907943. [Google Scholar] [CrossRef]
- Li, M.; Li, Q.; Wang, Y.; Huang, J.; Liu, P. Blastocyst cryopreservation and cryopreservation-warming transfer is an effective embryo transfer strategy for day 1 rescue intracytoplasmic sperm injection cycles. Sci. Rep. 2021, 11, 8260. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.J.; Zhang, X.Y.; Ding, G.L.; Li, R.; Wang, L.; Jin, L.; Lin, X.H.; Gao, L.; Sheng, J.Z.; Huang, H.F. Aquaporin7 plays a crucial role in tolerance to hyperosmotic stress and in the survival of oocytes during cryopreservation. Sci. Rep. 2015, 5, 17741. [Google Scholar] [CrossRef] [Green Version]
- Yokonishi, T.; Sato, T.; Komeya, M.; Katagiri, K.; Kubota, Y.; Nakabayashi, K.; Hata, K.; Inoue, K.; Ogonuki, N.; Ogura, A.; et al. Offspring production with sperm grown in vitro from cryopreserved testis tissues. Nat. Commun. 2014, 5, 4320. [Google Scholar] [CrossRef] [Green Version]
- Nevi, L.; Cardinale, V.; Carpino, G.; Costantini, D.; Di Matteo, S.; Cantafora, A.; Melandro, F.; Brunelli, R.; Bastianelli, C.; Aliberti, C.; et al. Cryopreservation protocol for human biliary tree stem/progenitors, hepatic and pancreatic precursors. Sci. Rep. 2017, 7, 6080. [Google Scholar] [CrossRef] [Green Version]
- Yong, K.W.; Pingguan-Murphy, B.; Xu, F.; Abas, W.A.W.; Choi, J.R.; Omar, S.Z.; Azmi, M.A.N.; Chua, K.H.; Safwani, W. Phenotypic and Functional Characterization of Long-Term Cryopreserved Human Adipose-derived Stem Cells. Sci. Rep. 2015, 5, 9596. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, P.M.; Banales, J.M. Applications of organoids in regenerative medicine: A proof-of-concept for biliary injury. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 371–372. [Google Scholar] [CrossRef]
- Wu, K.Z.; Shardt, N.; Laouar, L.; Elliott, J.A.W.; Jomha, N.M. Vitrification of particulated articular cartilage via calculated protocols. npj Regen. Med. 2021, 6, 15. [Google Scholar] [CrossRef]
- Hentze, J.L.; Kringelbach, T.M.; Novotny, G.W.; Hamid, B.H.; Ravn, V.; Christensen, I.J.; Hogdall, C.; Hogdall, E. Optimized Biobanking Procedures for Preservation of RNA in Tissue: Comparison of Snap-Freezing and RNAlater-Fixation Methods. Biopreserv. Biobank. 2019, 17, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Hansell, C. In the deep freeze. Nat. Chem. 2014, 6, 270. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.W.; Lee, G.W.; An, S.; Seong, K.Y.; Lee, J.S.; Yang, S.Y. Enhanced Cellular Cryopreservation by Biopolymer-Associated Suppression of RhoA/ROCK Signaling Pathway. Materials 2021, 14, 6056. [Google Scholar] [CrossRef]
- de Rougemont, O.; Breitenstein, S.; Leskosek, B.; Weber, A.; Graf, R.; Clavien, P.A.; Dutkowski, P. One hour hypothermic oxygenated perfusion (HOPE) protects nonviable liver allografts donated after cardiac death. Ann. Surg. 2009, 250, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P. Kinetics of Water Loss from Cells at Subzero Temperatures and Likelihood of Intracellular Freezing. J. Gen. Physiol. 1963, 47, 347. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Zhao, G. Ice Inhibition for Cryopreservation: Materials, Strategies, and Challenges. Adv. Sci. 2021, 8, 2002425. [Google Scholar] [CrossRef]
- Hirano, T.; Kawai, T.; Tsunoda, N.; Yamaya, G.; Obara, M.; Miyamoto, I.; Yamada, H. Clinical evaluation of cryosurgery for vascular lesions in the oral cavity. J. Oral Maxillofac. Surg. Med. Pathol. 2021, 33, 587–591. [Google Scholar] [CrossRef]
- Gignoux, B.M.; Ducerf, C.; Mabrut, J.Y.; Rivoire, M.; Rode, A.; Baulieux, J. Cryosurgery for primary and metastatic liver cancers. Ann. Chir. 2001, 126, 950–959. [Google Scholar] [CrossRef]
- Afsar, F.S.; Erkan, C.D.; Karaca, S. Clinical practice trends in cryosurgery: A retrospective study of cutaneous lesions. Postep. Dermatol. I Alergol. 2015, 32, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Bojic, S.; Murray, A.; Bentley, B.L.; Spindler, R.; Pawlik, P.; Cordeiro, J.L.; Bauer, R.; de Magalhaes, J.P. Winter is coming: The future of cryopreservation. BMC Biol. 2021, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P.; Rall, W.F.; Leibo, S.P. Kinetics of Water-Loss and the Likelihood of Intracellular Freezing in Mouse Ova–Influence of The Method of Calculating the Temperature-Dependence of Water Permeability. Cell Biophys. 1984, 6, 197–213. [Google Scholar] [CrossRef]
- Lee, Y.A.; Kim, Y.H.; Ha, S.J.; Kim, K.J.; Kim, B.J.; Kim, B.G.; Choi, S.H.; Kim, I.C.; Schmidt, J.A.; Ryu, B.Y. Cryopreservation of porcine spermatogonial stem cells by slow-freezing testis tissue in trehalose. J. Anim. Sci. 2014, 92, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; He, X.; Yarmush, M.L. Advanced technologies for the preservation of mammalian biospecimens. Nat. Biomed. Eng. 2021, 5, 793–804. [Google Scholar] [CrossRef]
- Gurruchaga, H.; del Burgo, L.S.; Hernandez, R.M.; Orive, G.; Selden, C.; Fuller, B.; Ciriza, J.; Pedraz, J.L. Advances in the slow freezing cryopreservation of microencapsulated cells. J. Control. Release 2018, 281, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ryu, K.J.; Kim, B.; Kang, D.; Kim, Y.Y.; Kim, T. Comparison between Slow Freezing and Vitrification for Human Ovarian Tissue Cryopreservation and Xenotransplantation. Int. J. Mol. Sci. 2019, 20, 3346. [Google Scholar] [CrossRef] [Green Version]
- Mazur, P.; Leibo, S.P.; Chu, E.H.Y. 2-Factor Hypothesis of Freezing Injury—Evidence from Chinese-Hamster Tissue-Culture Cells. Exp. Cell Res. 1972, 71, 345. [Google Scholar] [CrossRef]
- Liu, J.; Phy, J.; Yeomans, E. Theoretic considerations regarding slow cooling and vitrification during cryopreservation. Theriogenology 2012, 78, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Fahy, G.M.; Macfarlane, D.R.; Angell, C.A.; Meryman, H.T. Vitrification as an Approach to Cryopreservation. Cryobiology 1984, 21, 407–426. [Google Scholar] [CrossRef]
- Rienzi, L.; Gracia, C.; Maggiulli, R.; LaBarbera, A.R.; Kaser, D.J.; Ubaldi, F.M.; Vanderpoel, S.; Racowsky, C. Oocyte, embryo and blastocyst cryopreservation in ART: Systematic review and meta-analysis comparing slow-freezing versus vitrification to produce evidence for the development of global guidance. Hum. Reprod. Update 2017, 23, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Characteristics of human embryonic stem cells cryopreserved by conventional slow freezing and vitrification. Clin. Biochem. 2005, 38, 861.
- Sanfilippo, S.; Canis, M.; Smitz, J.; Sion, B.; Darcha, C.; Janny, L.; Brugnon, F. Vitrification of human ovarian tissue: A practical and relevant alternative to slow freezing. Reprod. Biol. Endocrinol. 2015, 13, 67. [Google Scholar] [CrossRef] [Green Version]
- Tam, E.; McGrath, M.; Sladkova, M.; AlManaie, A.; Alostaad, A.; de Peppo, G.M. Hypothermic and cryogenic preservation of tissue-engineered human bone. Ann. N. Y. Acad. Sci. 2020, 1460, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Cheng, Y.; Pan, Q.; Zhang, Y.J.; Jia, D.G.; Liu, Y.F. Effect of the Selective NLRP3 Inflammasome Inhibitor mcc950 on Transplantation Outcome in a Pig Liver Transplantation Model with Organs from Donors after Circulatory Death Preserved by Hypothermic Machine Perfusion. Transplantation 2019, 103, 353–362. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, J.F. Hypothermic organ preservation by static storage methods: Current status and a view to the future. Cryobiology 2010, 60, S13–S19. [Google Scholar] [CrossRef] [PubMed]
- Tchilikidi, K.Y. Liver graft preservation methods during cold ischemia phase and normothermic machine perfusion. World J. Gastrointest. Surg. 2019, 11, 126–142. [Google Scholar] [CrossRef]
- Que, W.; Hu, X.; Fujino, M.; Terayama, H.; Sakabe, K.; Fukunishi, N.; Zhu, P.; Yi, S.-Q.; Yamada, Y.; Zhong, L.; et al. Prolonged Cold Ischemia Time in Mouse Heart Transplantation Using Supercooling Preservation. Transplantation 2020, 104, 1879–1889. [Google Scholar] [CrossRef]
- Shamseddine, I.; Pennec, F.; Biwole, P.; Fardoun, F. Supercooling of phase change materials: A review. Renew. Sustain. Energy Rev. 2022, 158, 112172. [Google Scholar] [CrossRef]
- Rodriguez, J.V.; Almada, L.L.; Mamprin, M.E.; Guibert, E.E.; Furno, G. Subzero nonfreezing storage of rat hepatocytes using modified University of Wisconsin solution (mUW) and 1,4-butanediol. I- effects on cellular metabolites during cold storage. Ann. Hepatol. 2009, 8, 57–62. [Google Scholar] [CrossRef]
- Huang, H.; Yarmush, M.L.; Usta, O.B. Long-term deep-supercooling of large-volume water and red cell suspensions via surface sealing with immiscible liquids. Nat. Commun. 2018, 9, 3201. [Google Scholar] [CrossRef] [Green Version]
- Wakayama, S.; Ito, D.; Hayashi, E.; Ishiuchi, T.; Wakayama, T. Healthy cloned offspring derived from freeze-dried somatic cells. Nat. Commun. 2022, 13, 3666. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, G.R.; Torok, Z.; Bali, R.; Dwyre, D.M.; Little, E.; Walker, N.J.; Tablin, F.; Crowe, J.H.; Tsvetkova, N.M. Loading red blood cells with trehalose: A step towards biostabilization. Cryobiology 2004, 49, 123–136. [Google Scholar] [CrossRef]
- Keskintepe, L.; Eroglu, A. Cryopreservation and Freeze-Drying Protocols. In Methods in Molecular Biology, 3rd ed.; Wolkers, W.F., Oldenhof, H., Eds.; Springer: New York, NY, USA, 2015; Volume 1257, pp. 489–497. [Google Scholar]
- Ammar, M.M.; Waly, G.H.; Saniour, S.H.; Moussa, T.A. Growth factor release and enhanced encapsulated periodontal stem cells viability by freeze-dried platelet concentrate loaded thermo-sensitive hydrogel for periodontal regeneration. Saudi Dent. J. 2018, 30, 355–364. [Google Scholar] [CrossRef]
- Reardon, A.J.F.; Elliott, J.A.W.; McGann, L.E. Investigating membrane and mitochondrial cryobiological responses of HUVEC using interrupted cooling protocols. Cryobiology 2015, 71, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Huang, Z.; Liu, B.; He, X.; Xue, S.; Yan, X.; Jaganathan, G.K. Investigating solution effects injury of human T lymphocytes and its prevention during interrupted slow cooling. Cryobiology 2021, 99, 20–27. [Google Scholar] [CrossRef]
- Zhou, X.L.; Al Naib, A.; Sun, D.W.; Lonergan, P. Bovine oocyte vitrification using the Cryotop method: Effect of cumulus cells and vitrification protocol on survival and subsequent development. Cryobiology 2010, 61, 66–72. [Google Scholar] [CrossRef]
- Abazari, A.; Jomha, N.M.; Elliott, J.A.W.; McGann, L.E. Cryopreservation of articular cartilage. Cryobiology 2013, 66, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Chow-shi-yee, M.; Grondin, M.; Ouellet, F.; Averill-Bates, D.A. Control of stress-induced apoptosis by freezing tolerance-associated wheat proteins during cryopreservation of rat hepatocytes. Cell Stress Chaperones 2020, 25, 869–886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Q.; Ding, N.; Zeng, Y.-F.; Xiang, Y.-Y.; Yang, M.-W.; Hong, F.-F.; Yang, S.-L. New progress in roles of nitric oxide during hepatic ischemia reperfusion injury. World J. Gastroenterol. 2017, 23, 2505–2510. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.-E.; Ahn, J.S.; Kim, Y.-H.; Oh, H.-J.; Kim, B.-J.; Ryu, B.-Y. Necrostatin-1 improves the cryopreservation efficiency of murine spermatogonial stem cells via suppression of necroptosis and apoptosis. Theriogenology 2020, 158, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Karlsson, J.O.M.; Aksan, A. FTIR Analysis of Molecular Changes Associated with Warming Injury in Cryopreserved Leukocytes. Langmuir 2019, 35, 7552–7559. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.U.; Lien, Y.R.; Chen, H.F.; Chang, L.J.; Tsai, Y.Y.; Yang, Y.S. Observational clinical follow-up of oocyte cryopreservation using a slow-freezing method with 1,2-propanediol plus sucrose followed by ICSI. Hum. Reprod. 2005, 20, 1975–1980. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zhang, Q.; Zhao, Y.; Ren, L.; Yuan, X. Trehalose-functional glycopeptide enhances glycerol-free cryopreservation of red blood cells. J. Mater. Chem. B 2019, 7, 5695–5703. [Google Scholar] [CrossRef]
- Marques, L.S.; Fossati, A.A.N.; Rodrigues, R.B.; Da Rosa, H.T.; Izaguirry, A.P.; Ramalho, J.B.; Moreira, J.C.F.; Santos, F.W.; Zhang, T.; Streit, D.P. Slow freezing versus vitrification for the cryopreservation of zebrafish (Danio rerio) ovarian tissue. Sci. Rep. 2019, 9, 15353. [Google Scholar] [CrossRef] [Green Version]
- Perumal, P. Reduced glutathione and cysteine hydrochloride on crossbred bull semen. Nat. Preced. 2012, 46, 636–641. [Google Scholar] [CrossRef]
- Mitrus, I.; Smagur, A.; Fidyk, W.; Czech, M.; Prokop, M.; Chwieduk, A.; Glowala-Kosinska, M.; Czerw, T.; Sobczyk-Kruszelnicka, M.; Mendrek, W.; et al. Reduction of DMSO concentration in cryopreservation mixture from 10% to 7.5% and 5% has no impact on engraftment after autologous peripheral blood stem cell transplantation: Results of a prospective, randomized study. Bone Marrow Transplant. 2018, 53, 274–280. [Google Scholar] [CrossRef] [Green Version]
- Tasoulis, M.K.; Douzinas, E.E. Hypoxemic reperfusion of ischemic states: An alternative approach for the attenuation of oxidative stress mediated reperfusion injury. J. Biomed. Sci. 2016, 23, 7. [Google Scholar] [CrossRef] [Green Version]
- Karangwa, S.; Panayotova, G.; Dutkowski, P.; Porte, R.J.; Guarrera, J.V.; Schlegel, A. Hypothermic machine perfusion in liver transplantation. Int. J. Surg. 2020, 82, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and its clinical applications. Integr. Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef]
- Chen, Z.; Kebschull, L.; Foell, D.A.; Rauen, U.; Hansen, U.; Heitplatz, B.; Hessler, M.; Senninger, N.; Vogel, T.; Mohr, A.; et al. A novel histidine-tryptophan-ketoglutarate formulation ameliorates intestinal injury in a cold storage and ex vivo warm oxygenated reperfusion model in rats. Biosci. Rep. 2020, 40, BSR20191989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valipour, J.; Nashtaei, M.S.; Khosravizadeh, Z.; Mahdavinezhad, F.; Nekoonam, S.; Esfandyari, S.; Amidi, F. Effect of sulforaphane on apoptosis, reactive oxygen species and lipids peroxidation of human sperm during cryopreservation. Cryobiology 2021, 99, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Cherkashina, D.V.; Sosimchyk, I.A.; Semenchenko, O.A.; Semenchenko, A.Y.; Volina, V.V.; Petrenko, A.Y. Bioregulators of stem and progenitor cells in preservation solution reduce cold ischemia-reperfusion injury of isolated rat livers. Biofactors 2016, 42, 287–295. [Google Scholar] [CrossRef]
- Magalhaes, R.; Nugraha, B.; Pervaiz, S.; Yu, H.; Kuleshova, L.L. Influence of cell culture configuration on the post-cryopreservation viability of primary rat hepatocytes. Biomaterials 2012, 33, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Men, H.S.; Monson, R.L.; Parrish, J.J.; Rutledge, J.J. Degeneration of cryopreserved bovine oocytes via apoptosis during subsequent culture. Cryobiology 2003, 47, 73–81. [Google Scholar] [CrossRef]
- Villalba, R.; Pena, J.; Navarro, P.; Luque, E.; Jimena, I.; Romero, A.; Gomez Villagran, J.L. Cryopreservation increases apoptosis in human menisci. Knee Surg. Sport. Traumatol. Arthrosc. 2012, 20, 298–303. [Google Scholar] [CrossRef]
- Wan, L.; Powell-Palm, M.J.; Lee, C.; Gupta, A.; Weegman, B.P.; Clemens, M.G.; Rubinsky, B. Preservation of rat hearts in subfreezing temperature isochoric conditions to-8 degrees C and 78 MPa. Biochem. Biophys. Res. Commun. 2018, 496, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Oner, S.; Ercan, F.; Arbak, S. Time-dependent morphological alterations of cold-stored small bowel in Euro-Collins and Ringer’s lactate solutions. Acta Histochem. 2004, 106, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y. Apoptosis and necrosis: Intracellular ATP level as a determinant for cell death modes. Cell Death Differ. 1997, 4, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Dallaporta, B.; Resche-Rigon, M. The mitochondrial death/life regulator in apoptosis and necrosis. Annu. Rev. Physiol. 1998, 60, 619–642. [Google Scholar] [CrossRef]
- Linkermann, A.; Green, D.R. Necroptosis. N. Engl. J. Med. 2014, 370, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Bissoyi, A.; Pramanik, K. Role of the Apoptosis Pathway in Cryopreservation-Induced Cell Death in Mesenchymal Stem Cells Derived from Umbilical Cord Blood. Biopreserv. Biobank. 2014, 12, 246–254. [Google Scholar] [CrossRef]
- Desoutter, J.; Ossart, C.; Lacassagne, M.N.; Regnier, A.; Marolleau, J.P.; Harrivel, V. Cryopreservation and thawing of hematopoietic stem cell CD34-induced apoptosis through caspase pathway activation: Key role of granulocytes. Cytotherapy 2019, 21, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; Wang, H.C.; Wang, H.X.; Ruan, L.H.; Zhang, Y.M.; Li, J.T.; Tian, S.; Zhang, Y.C. Oxidative stress and activities of caspase-8,-9, and-3 are involved in cryopreservation-induced apoptosis in granulosa cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 166, 52–55. [Google Scholar] [CrossRef]
- Martin, G.; Cagnon, N.; Sabido, O.; Sion, B.; Grizard, G.; Durand, P.; Levy, R. Kinetics of occurrence of some features of apoptosis during the cryopreservation process of bovine spermatozoa. Hum. Reprod. 2007, 22, 380–388. [Google Scholar] [CrossRef]
- Feng, T.Y.; Li, Q.; Ren, F.; Xi, H.M.; Lv, D.L.; Li, Y.; Hu, J.H. Melatonin Protects Goat Spermatogonial Stem Cells against Oxidative Damage during Cryopreservation by Improving Antioxidant Capacity and Inhibiting Mitochondrial Apoptosis Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 5954635. [Google Scholar] [CrossRef]
- Ball, B.A. Oxidative stress, osmotic stress and apoptosis: Impacts on sperm function and preservation in the horse. Anim. Reprod. Sci. 2008, 107, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.E.; Oh, H.J.; Ahn, J.S.; Kim, Y.H.; Kim, B.J.; Ryu, B.Y. Antioxidant or Apoptosis Inhibitor Supplementation in Culture Media Improves Post-Thaw Recovery of Murine Spermatogonial Stem Cells. Antioxidants 2021, 10, 754. [Google Scholar] [CrossRef] [PubMed]
- Bissoyi, A.; Nayak, B.; Pramanik, K.; Sarangi, S.K. Targeting Cryopreservation-Induced Cell Death: A Review. Biopreserv. Biobank. 2014, 12, 23–34. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Pero, M.E.; Zullo, G.; Esposito, L.; Iannuzzi, A.; Lombardi, P.; De Canditiis, C.; Neglia, G.; Gasparrini, B. Inhibition of apoptosis by caspase inhibitor Z-VAD-FMK improves cryotolerance of in vitro derived bovine embryos. Theriogenology 2018, 108, 127–135. [Google Scholar] [CrossRef]
- Heng, B.C.; Clement, M.V.; Cao, T. Caspase inhibitor Z-VAD-FMK enhances the freeze-thaw survival rate of human embryonic stem cells. Biosci. Rep. 2007, 27, 257–264. [Google Scholar] [CrossRef]
- Pagano, N.; Longobardi, V.; De Canditiis, C.; Zuchegna, C.; Romano, A.; Andrzej, K.M.; Pero, M.E.; Gasparrini, B. Effect of caspase inhibitor Z-VAD-FMK on bovine sperm cryotolerance. Reprod. Domest. Anim. 2020, 55, 530–536. [Google Scholar] [CrossRef]
- Li, X.Y.; Krawetz, R.; Liu, S.Y.; Meng, G.L.; Rancourt, D.E. ROCK inhibitor improves survival of cryopreserved serum/feeder-free single human embryonic stem cells. Hum. Reprod. 2009, 24, 580–589. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Kalia, V.; Montelaro, R.C. Caspase-mediated apoptosis and cell death of rhesus macaque CD4(+) T-cells due to cryopreservation of peripheral blood mononuclear cells can be rescued by cytokine treatment after thawing. Cryobiology 2003, 47, 44–58. [Google Scholar] [CrossRef]
- Ferrusola, C.O.; Fernandez, L.G.; Sandoval, C.S.; Garcia, B.M.; Martinez, H.R.; Tapia, J.A.; Pena, F.J. Inhibition of the mitochondrial permeability transition pore reduces “apoptosis like” changes during cryopreservation of stallion spermatozoa. Theriogenology 2010, 74, 458–465. [Google Scholar] [CrossRef]
- Zeng, C.J.; Tang, K.Y.; He, L.; Peng, W.P.; Ding, L.; Fang, D.H.; Zhang, Y. Effects of glycerol on apoptotic signaling pathways during boar spermatozoa cryopreservation. Cryobiology 2014, 68, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Shanehbandi, D.; Hamdi, K.; Rahbar, S.; Pashaiasl, M. Influence of cryopreservation on structure and function of mammalian spermatozoa: An overview. Cell Tissue Bank. 2020, 21, 1–15. [Google Scholar] [CrossRef]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of apoptosis in sperm cryoinjury. Reprod. Biomed. Online 2010, 21, 456–462. [Google Scholar] [CrossRef]
- Moustafa, M.H.; Sharma, R.K.; Thornton, J.; Mascha, E.; Abdel-Hafez, M.A.; Thomas, A.J.; Agarwal, A. Relationship between ROS production, apoptosis and DNA denaturation in spermatozoa from patients examined for infertility. Hum. Reprod. 2004, 19, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadam, M.; Fard, Y.A.; Saki, G.; Nikbakht, R. Effect of vitamin D on apoptotic marker, reactive oxygen species and human sperm parameters during the process of cryopreservation. Iran. J. Basic Med. Sci. 2019, 22, 1036–1043. [Google Scholar] [CrossRef]
- Xu, X.; Cowley, S.; Flaim, C.J.; James, W.; Seymour, L.; Cui, Z. The Roles of Apoptotic Pathways in the Low Recovery Rate After Cryopreservation of Dissociated Human Embryonic Stem Cells. Biotechnol. Prog. 2010, 26, 827–837. [Google Scholar] [CrossRef]
- Rezaei, N.; Mohammadi, M.; Mohammadi, H.; Khalatbari, A.; Zare, Z. Acrosome and chromatin integrity, oxidative stress, and expression of apoptosis-related genes in cryopreserved mouse epididymal spermatozoa treated with L-Carnitine. Cryobiology 2020, 95, 171–176. [Google Scholar] [CrossRef]
- Galluzzi, L.; Kepp, O.; Krautwald, S.; Kroemer, G.; Linkermann, A. Molecular mechanisms of regulated necrosis. Semin. Cell Dev. Biol. 2014, 35, 24–32. [Google Scholar] [CrossRef]
- Seo, J.; Nam, Y.W.; Kim, S.; Oh, D.-B.; Song, J. Necroptosis molecular mechanisms: Recent findings regarding novel necroptosis regulators. Exp. Mol. Med. 2021, 53, 1007–1017. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, P.; Hua, F.Z.; Hu, Y.H.; Xiao, F.; Liu, Q.; Huang, D.; Deng, F.M.; Wei, G.; Deng, W.; et al. Sevoflurane postconditioning reduces myocardial ischemia reperfusion injury-induced necroptosis by up-regulation of OGT-mediated O-GlcNAcylated RIPK3. Aging 2020, 12, 25452–25468. [Google Scholar] [CrossRef]
- Venkataraman, M. Effects of cryopreservation on immune responses.11. Heightened secretion of tumor necrosis factor-alpha by frozen human peripheral blood mononuclear cells. Cryobiology 1997, 34, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Vara, E.; AriasDiaz, J.; Garcia, C.; Balibrea, J.L. Influence of cryopreservation on the sensitivity of human islets to tumor necrosis factor. Pancreas 1996, 13, 29–37. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, H.; Luo, D.; Yang, X.; Yao, J.; Zhang, C.; Lv, L.; Guo, Z.; Deng, C.; Li, Y.; et al. Inhibiting Necroptosis of Spermatogonial Stem Cell as a Novel Strategy for Male Fertility Preservation. Stem Cells Dev. 2020, 29, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, Y.V.; Minasyan, S.M.; Vasina, L.V.; Demchenko, E.A.; Galagudza, M.M. Effects of Inhibitors of Necroptosis and Autophagy on Morphofunctional Characteristics of the Myocardium during Static Cold Storage of Donor Rat Heart. Bull. Exp. Biol. Med. 2015, 159, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Zamel, R.; Rai, X.-H.; Lu, C.; Keshavjee, S.; Keshavjee, S.; Liu, M. Ischemia-reperfusion induces death receptor-independent necroptosis via calpain-STAT3 activation in a lung transplant setting. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L595–L608. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Apaijai, N.; Chattipakorn, N.; Chattipakorn, S.C. The possible roles of necroptosis during cerebral ischemia and ischemia/reperfusion injury. Arch. Biochem. Biophys. 2020, 695, 108629. [Google Scholar] [CrossRef]
- Weinlich, R.; Oberst, A.; Beere, H.M.; Green, D.R. Necroptosis in development, inflammation and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-Y.; Yiang, G.-t.; Liao, W.-T.; Tsai, A.P.-Y.; Cheng, Y.-L.; Cheng, P.-W.; Li, C.-Y.; Li, C.-J. Current Mechanistic Concepts in Ischemia and Reperfusion Injury. Cell. Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Vanlangenakker, N.; Vanden Berghe, T.; Krysko, D.V.; Festjens, N.; Vandenabeele, P. Molecular mechanisms and pathophysiology of necrotic cell death. Curr. Mol. Med. 2008, 8, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Goerlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nastos, C.; Kalimeris, K.; Papoutsidakis, N.; Tasoulis, M.K.; Lykoudis, P.M.; Theodoraki, K.; Nastou, D.; Smyrniotis, V.; Arkadopoulos, N. Global Consequences of Liver Ischemia/Reperfusion Injury. Oxid. Med. Cell. Longev. 2014, 2014, 906965. [Google Scholar] [CrossRef] [Green Version]
- Panisello-Rosello, A.; Rosello-Catafau, J. Molecular Mechanisms and Pathophysiology of Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2018, 19, 4093. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Petrowsky, H.; Hong, J.C.; Busuttil, R.W.; Kupiec-Weglinski, J.W. Ischaemia-reperfusion injury, in liver transplantation-from bench to bedside. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Dar, W.A.; Sullivan, E.; Bynon, J.S.; Eltzschig, H.; Ju, C. Ischaemia reperfusion injury in liver transplantation: Cellular and molecular mechanisms. Liver Int. 2019, 39, 788–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, P.M.; Connolly, M.R.; Coe, T.M.; Calhoun, A.; Pollok, F.; Markmann, J.F.; Burdorf, L.; Azimzadeh, A.; Madsen, J.C.; Pierson, R.N. Minimizing Ischemia Reperfusion Injury in Xenotransplantation. Front. Immunol. 2021, 12, 681504. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Man, K. New Insights in Mechanisms and Therapeutics for Short- and Long-Term Impacts of Hepatic Ischemia Reperfusion Injury Post Liver Transplantation. Int. J. Mol. Sci. 2021, 22, 8210. [Google Scholar] [CrossRef] [PubMed]
- Jassem, W.; Heaton, N.D. The role of mitochondria in ischemia/reperfusion injury in organ transplantation. Kidney Int. 2004, 66, 514–517. [Google Scholar] [CrossRef] [Green Version]
- Kosieradzki, M.; Rowinski, W. Ischemia/Reperfusion Injury in Kidney Transplantation: Mechanisms and Prevention. Transplant. Proc. 2008, 40, 3279–3288. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.K.; Ding, G.; Verna, N.; Ibrahim, S.; Oakes, S.; Austen, W.G.; Hechtman, H.B.; Moore, F.D. IgM binding to injured tissue precedes complement activation during skeletal muscle ischemia-reperfusion. J. Surg. Res. 2004, 122, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Akbari, G. Role of Zinc Supplementation on Ischemia/Reperfusion Injury in Various Organs. Biol. Trace Elem. Res. 2020, 196, 1–9. [Google Scholar] [CrossRef]
- Ogawa, T.; Mimura, Y. Antioxidant effect of zinc on acute renal failure induced by ischemia-reperfusion injury in rats. Am. J. Nephrol. 1999, 19, 609–614. [Google Scholar] [CrossRef]
- Barekat, F.; Talebi, A.; Nematbakhsh, M. The Protective Roles of zinc and estradiol in renal ischemia/reperfusion injury in ovariectomized rats. J. Nephropathol. 2018, 7, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.T.; Toledo-Pereyra, L.H.; Quesnelle, K.M. Selectins in Liver Ischemia and Reperfusion Injury. J. Investig. Surg. 2015, 28, 292–300. [Google Scholar] [CrossRef]
- Zhai, Y.; Shen, X.D.; O’Connell, R.; Gao, T.; Lassman, C.; Busuttil, R.W.; Cheng, G.H.; Kupiec-Weglinski, J.W. Cutting edge: TLR4 activation mediates liver ischemia/reperfusion inflammatory response via IFN regulatory factor 3-dependent MyD88-independent pathway. J. Immunol. 2004, 173, 7115–7119. [Google Scholar] [CrossRef] [Green Version]
- Tsung, A.; Hoffman, R.A.; Izuishi, K.; Critchlow, N.D.; Nakao, A.; Chan, M.H.; Lotze, M.T.; Geller, D.A.; Billiar, T.R. Hepatic ischemia/reperfusion injury involves functional TLR4 signaling in nonparenchymal cells. J. Immunol. 2005, 175, 7661–7668. [Google Scholar] [CrossRef] [Green Version]
- Datta, G.; Fuller, B.J.; Davidson, B.R. Molecular mechanisms of liver ischemia reperfusion injury: Insights from transgenic knockout models. World J. Gastroenterol. 2013, 19, 1683–1698. [Google Scholar] [CrossRef] [PubMed]
- Vardanian, A.J.; Busuttil, R.W.; Kupiec-Weglinski, J.W. Molecular mediators of liver ischemia and reperfusion injury: A brief review. Mol. Med. 2008, 14, 337–345. [Google Scholar] [CrossRef]
- Abu-Elmagd, K.M.; Costa, G.; Bond, G.J.; Soltys, K.; Sindhi, R.; Wu, T.; Koritsky, D.A.; Schuster, B.; Martin, L.; Cruz, R.J.; et al. Five Hundred Intestinal and Multivisceral Transplantations at a Single Center Major Advances with New Challenges. Ann. Surg. 2009, 250, 567–581. [Google Scholar] [CrossRef]
- Anaya-Prado, R.; Toledo-Pereyra, L.H.; Lentsch, A.B.; Ward, P.A. Ischemia/reperfusion injury. J. Surg. Res. 2002, 105, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.S.; Da Silva, R.T.; Machado, I.F.; Panisello-Rosello, A.; Rosello-Catafau, J.; Rolo, A.P.; Palmeira, C.M. Shaping of Hepatic Ischemia/Reperfusion Events: The Crucial Role of Mitochondria. Cells 2022, 11, 688. [Google Scholar] [CrossRef]
- Lubawy, J.; Chowanski, S.; Adamski, Z.; Slocinska, M. Mitochondria as a target and central hub of energy division during cold stress in insects. Front. Zool. 2022, 19, 1. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Species | Cell Type | Cryopreservation Method | Apoptotic Pathway |
|---|---|---|---|---|
| Jian-Min et al. [77] | Rats | Granulosa cells | Vitrification | Pathway I |
| Xiangyun et al. [87] | Humans | Embryonic stem cells | Slow freezing | Pathway IV |
| Akalabya et al. [75] | Humans | Mesenchymal stem cells | Slow freezing | Pathway II |
| Xia et al. [95] | Humans | Embryonic stem cells | Slow freezing | Pathway IV |
| Noorollah et al. [96] | Humans | Sperm | Vitrification | Pathway III |
| Surojit et al. [88] | Rhesus macaques | Peripheral blood mononuclear cells | Slow freezing | Pathway V |
| Mélanie et al. [52] | Rats | Hepatocytes | Slow freezing | Pathway III |
| Laboratoire et al. [76] | Humans | Hematopoietic stem cells | Slow freezing | Pathway I |
| Study | Species | Cell or Organ Type | Preservation Method |
|---|---|---|---|
| Xie et al. [102] | Mice | Spermatogonial stem cells | Slow freezing |
| Venkataraman et al. [100] | Humans | Peripheral blood mononuclear cells | Slow freezing |
| Vara et al. [101] | Humans | Islets | Slow freezing |
| Dmitriev et al. [103] | Rats | Heart | Static cold storage |
| Kim et al. [104] | Humans | Bronchial epithelium | Hypothermic machine perfusion |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Dang, W.; He, X.; Xu, F.; Huang, H. Biomolecular Pathways of Cryoinjuries in Low-Temperature Storage for Mammalian Specimens. Bioengineering 2022, 9, 545. https://doi.org/10.3390/bioengineering9100545
Fu Y, Dang W, He X, Xu F, Huang H. Biomolecular Pathways of Cryoinjuries in Low-Temperature Storage for Mammalian Specimens. Bioengineering. 2022; 9(10):545. https://doi.org/10.3390/bioengineering9100545
Chicago/Turabian StyleFu, Ying, Wenjun Dang, Xiaocong He, Feng Xu, and Haishui Huang. 2022. "Biomolecular Pathways of Cryoinjuries in Low-Temperature Storage for Mammalian Specimens" Bioengineering 9, no. 10: 545. https://doi.org/10.3390/bioengineering9100545
APA StyleFu, Y., Dang, W., He, X., Xu, F., & Huang, H. (2022). Biomolecular Pathways of Cryoinjuries in Low-Temperature Storage for Mammalian Specimens. Bioengineering, 9(10), 545. https://doi.org/10.3390/bioengineering9100545