

Optimization of the Fermentative Production of Rhizomucor miehei Lipase in Aspergillus oryzae by Controlling Morphology

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms, Media, and Culture Conditions
2.2. Optimization of Nitrogen Source Concentration and Agitation
2.3. Detection of RML Activity, Protein, Sugar, and DCW
2.4. Determination of Cell Morphology
2.5. Data Analysis
3. Results
3.1. Effects of Inoculated Spore Concentration on Cell Morphology and RML Production in Shake Flasks
3.2. Optimization of the RML Production Based on Morphology Control in 5 L Bioreactor
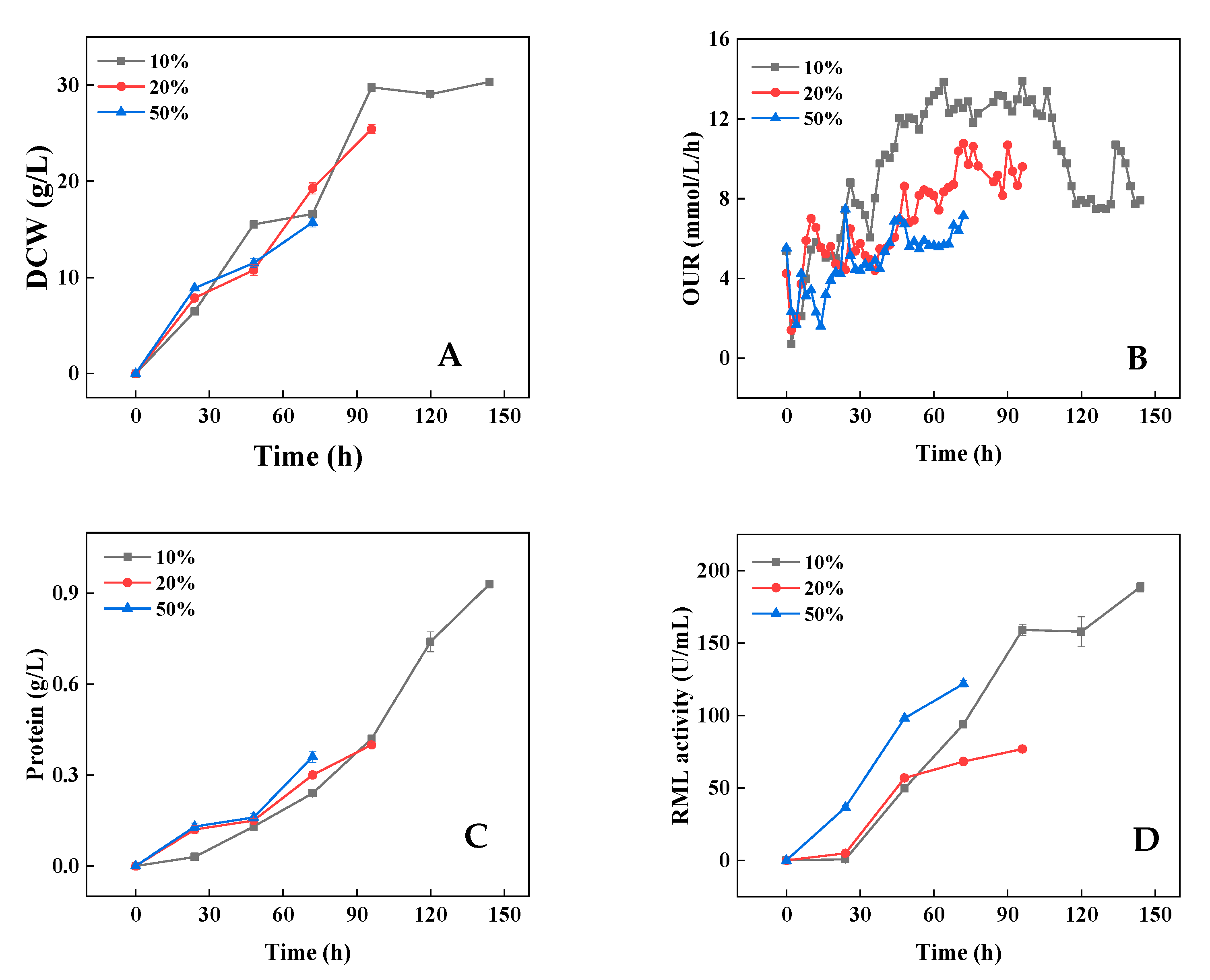
3.2.1. Morphology Control by Optimization of Nitrogen Source Supply Strategy
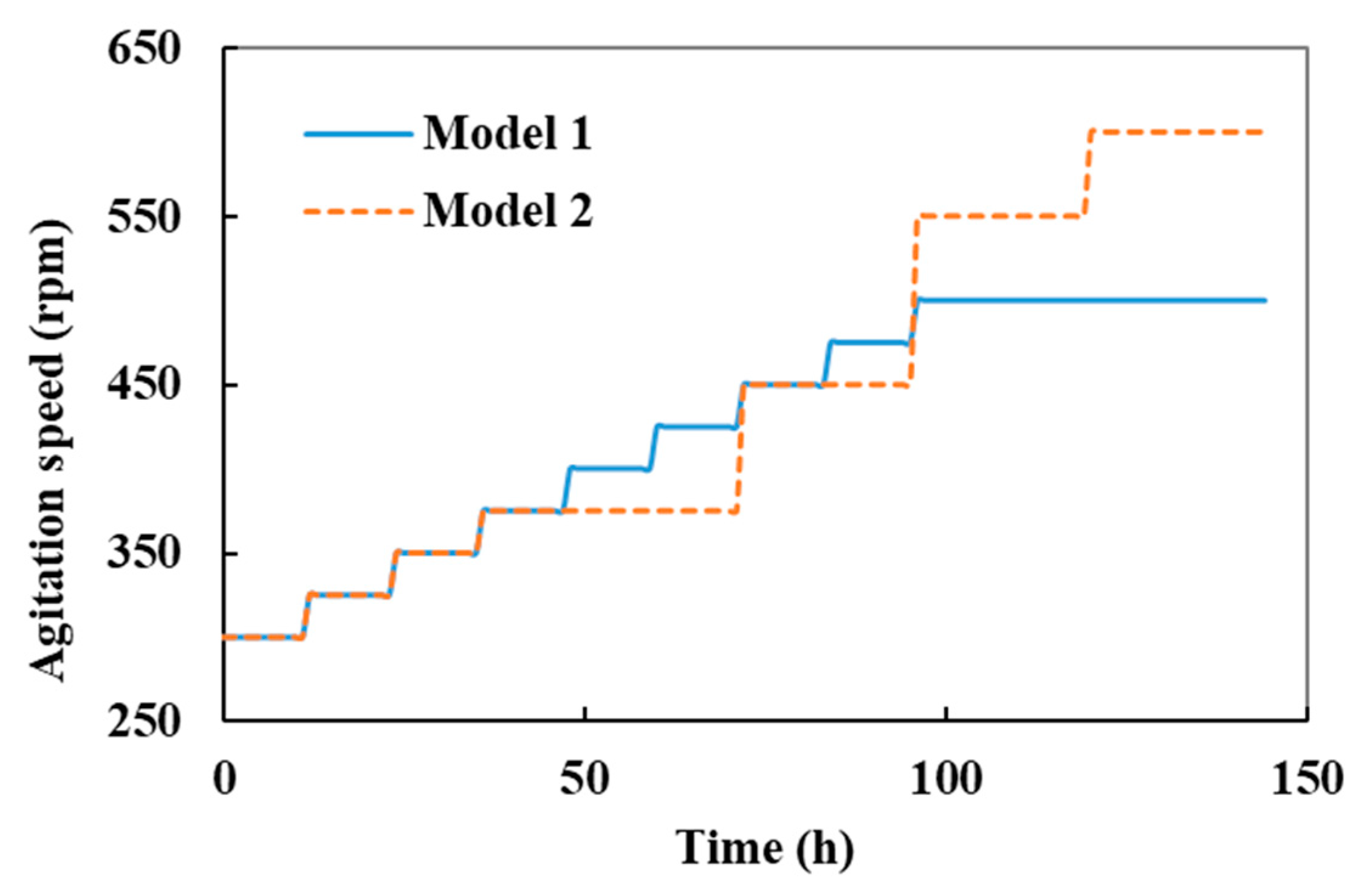
3.2.2. Morphology Control by Optimization of Agitation Strategy
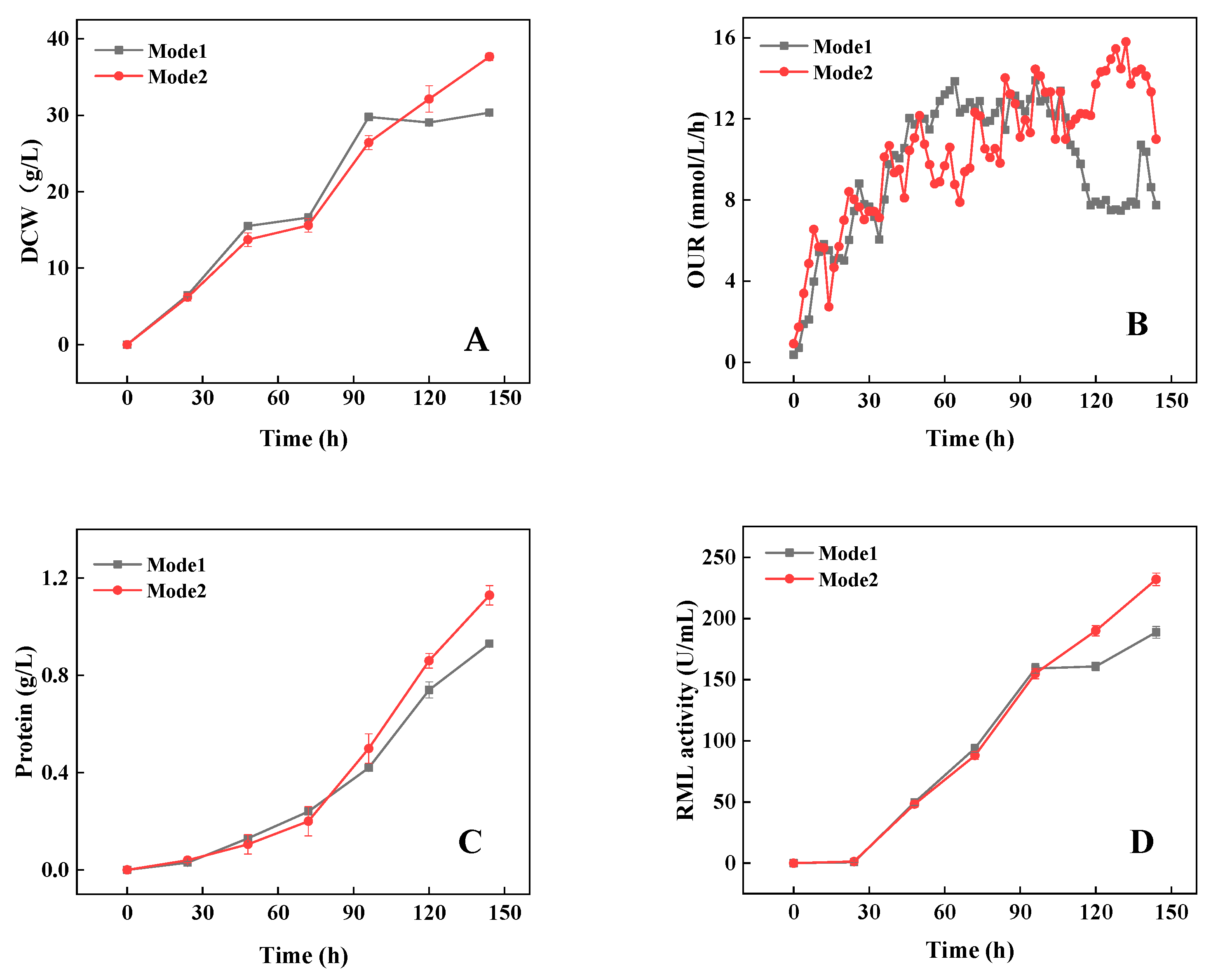
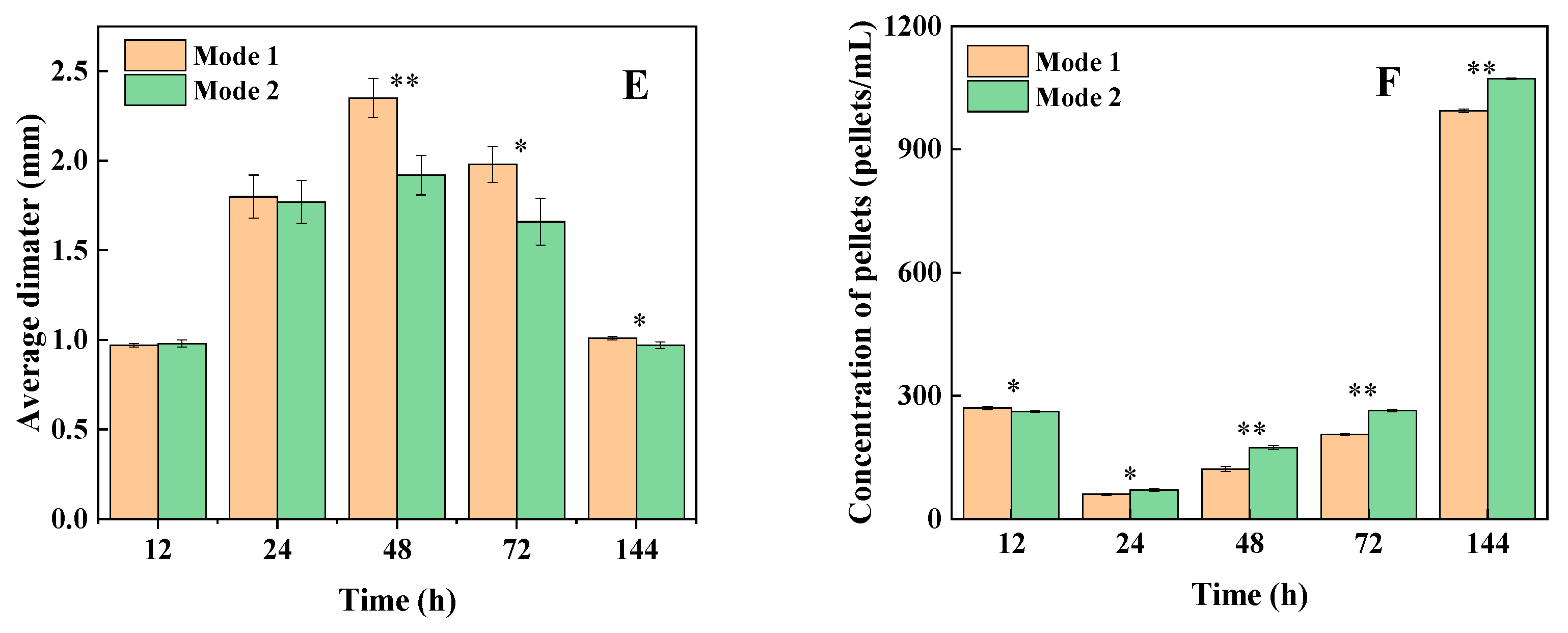
3.3. Strategy Verification Using a High-Yield Strain in a 5 L Bioreactor
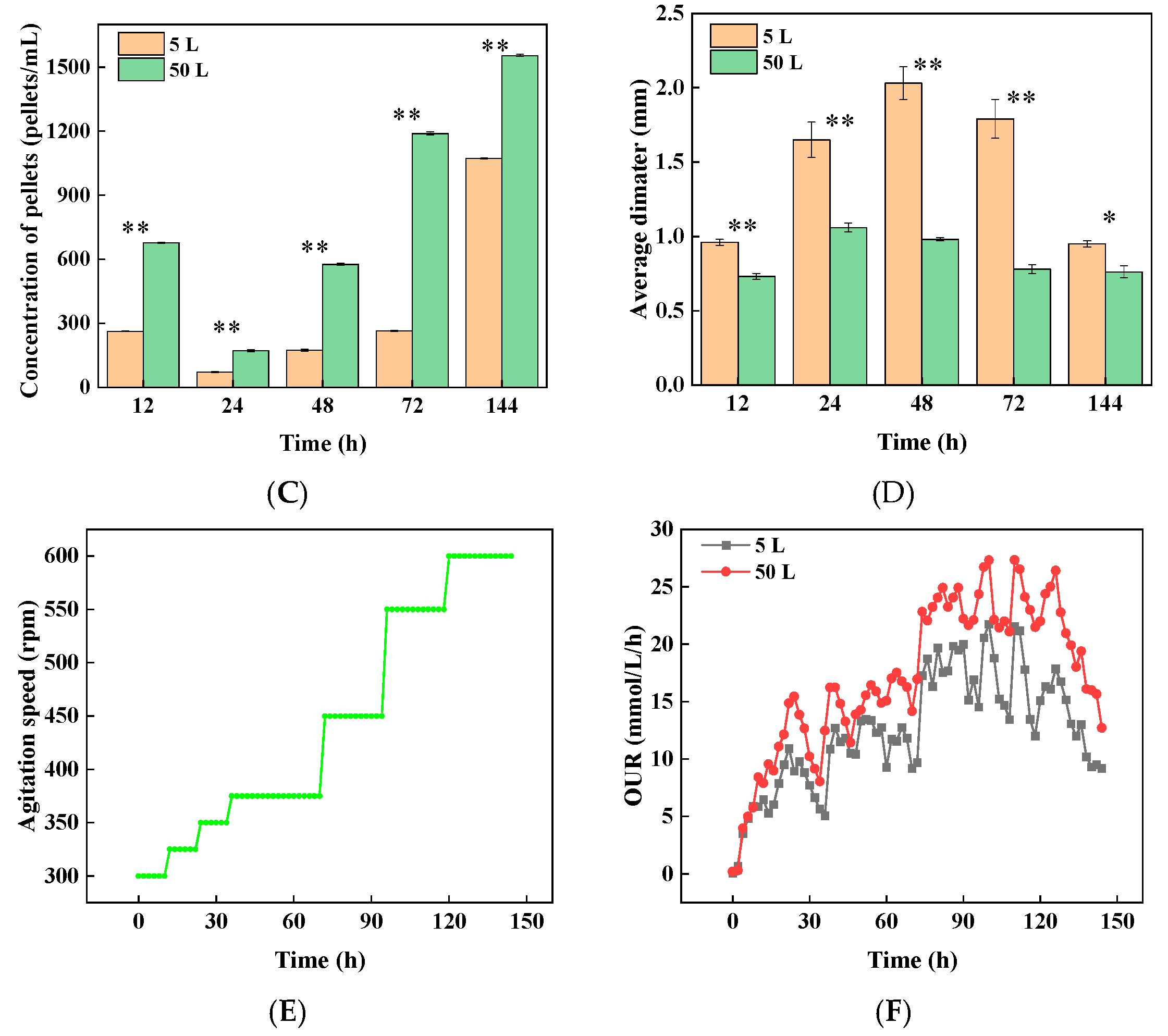
3.4. Strategy Verification and Scale-Up in a 50 L Bioreactor
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Liu, H.; Wang, P.; Gong, G.; Wang, L.; Zhao, G.; Zheng, Z. Morphology engineering of Penicillium chrysogenum by RNA silencing of chitin synthase gene. Biotechnol. Lett. 2013, 35, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Palla, C.A.; Pacheco, C.; Carrín, M.E. Production of structured lipids by acidolysis with immobilized Rhizomucor miehei lipases: Selection of suitable reaction conditions. J. Mol. Catal. B Enzym. 2012, 76, 106–115. [Google Scholar] [CrossRef]
- Gofferjé, G.; Stäbler, A.; Herfellner, T.; Schweiggert-Weisz, U.; Flöter, E. Kinetics of enzymatic esterification of glycerol and free fatty acids in crude Jatropha oil by immobilized lipase from Rhizomucor miehei. J. Mol. Catal. B Enzym. 2014, 107, 1–7. [Google Scholar] [CrossRef]
- Gumel, A.M.; Annuar, M.S.; Heidelberg, T.; Chisti, Y. Thermo-kinetics of lipase-catalyzed synthesis of 6-O-glucosyldecanoate. Bioresour. Technol. 2011, 102, 8727–8732. [Google Scholar] [CrossRef]
- He, D.; Luo, W.; Wang, Z.; Lv, P.; Yuan, Z.; Huang, S.; Xv, J. Establishment and application of a modified membrane-blot assay for Rhizomucor miehei lipases aimed at improving their methanol tolerance and thermostability. Enzym. Microb. Technol. 2017, 102, 35–40. [Google Scholar] [CrossRef]
- Luo, W.; He, D.; Fu, J.; Lv, P.; Yuan, Z.; Alam, M.A.; Wang, Z.; Huang, S. Effect of Propeptide Variation on Properties of Rhizomucor miehei Lipase. J. Biobased Mater. Bioenergy 2018, 12, 330–338. [Google Scholar] [CrossRef]
- Yildirim, D.; Baran, E.; Ates, S.; Yazici, B.; Tukel, S.S. Improvement of activity and stability of Rhizomucor miehei lipase by immobilization on nanoporous aluminium oxide and potassium sulfate microcrystals and their applications in the synthesis of aroma esters. Biocatal. Biotransform. 2018, 37, 210–223. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, Q.; Chen, L.; Zhang, C.; Bu, W.; Zhang, X.; Zhang, K.; Yang, Z. Improved production of recombinant Rhizomucor miehei lipase by coexpressing protein folding chaperones in Pichia pastoris, which triggered ER stress. Bioengineered 2020, 11, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Gögus, N.; Tari, C.; Oncü, S.; Unluturk, S.; Tokatli, F. Relationship between morphology, rheology and polygalacturonase production by Aspergillus sojae ATCC 20235 in submerged cultures. Biochem. Eng. J. 2006, 32, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Xu, Y.; Wang, D. Changes in morphology of Rhizopus chinensis in submerged fermentation and their effect on production of mycelium-bound lipase. Bioprocess. Biosyst. Eng. 2009, 32, 397–405. [Google Scholar] [CrossRef]
- Yu, L.; Chao, Y.; Wensel, P.; Chen, S. Hydrodynamic and kinetic study of cellulase production by Trichoderma reesei with pellet morphology. Biotechnol. Bioeng. 2012, 109, 1755–1768. [Google Scholar] [CrossRef] [PubMed]
- Metz, B.; Kossen, N.W.F. The growth of molds in the form of pellets–a literature review. Biotechnol. Bioeng. 1997, 19, 781–799. [Google Scholar] [CrossRef]
- Fang, X.; Zhao, G.; Dai, J.; Liu, H.; Wang, P.; Wang, L.; Song, J.; Zheng, Z. Macro-morphological characterization and kinetics of Mortierella alpine colonies during batch cultivation. PLoS ONE 2018, 13, 192803. [Google Scholar] [CrossRef] [PubMed]
- Veiter, L.; Kager, J.; Herwig, C. Optimal process design space to ensure maximum viability and productivity in Penicillium chrysogenum pellets during fed-batch cultivations through morphological and physiological control. Microb. Cell Fact. 2020, 19, 33. [Google Scholar] [CrossRef]
- El-Enshasy, H.; Kleine, J.; Rinas, U. Agitation effects on morphology and protein productive fractions of filamentous and pelleted growth forms of recombinant Aspergillus niger. Process. Biochem. 2006, 41, 2103–2112. [Google Scholar] [CrossRef]
- Xie, M.-h.; Xia, J.-y.; Zhou, Z.; Zhou, G.-z.; Chu, J.; Zhuang, Y.-p.; Zhang, S.-l.; Noorman, H. Power consumption, local and average volumetric mass transfer coefficient in multiple-impeller stirred bioreactors for xanthan gum solutions. Chem. Eng. Sci. 2014, 106, 144–156. [Google Scholar] [CrossRef]
- Driouch, H.; Roth, A.; Dersch, P.; Wittmann, C. Filamentous fungi in good shape: Microparticles for tailor-made fungal morphology and enhanced enzyme production. Bioeng. Bugs 2011, 2, 100–104. [Google Scholar] [CrossRef]
- Singh, B. Engineering fungal morphology for enhanced production of hydrolytic enzymes by Aspergillus oryzae SBS50 using microparticles. 3 Biotech 2018, 8, 283. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, Z.Y.; Zhu, L.; Zhan, X.B. Enhanced endo-β-1,3-glucanase production through morphology control of Trichoderma harzianum. J. Biol. 2016, 33, 6. [Google Scholar]
- Haack, M.B.; Olsson, L.; Hansen, K.; Eliasson Lantz, A. Change in hyphal morphology of Aspergillus oryzae during fed-batch cultivation. Appl. Microbiol. Biotechnol. 2006, 70, 482–487. [Google Scholar] [CrossRef]
- Resina, D.; Serrano, A.; Valero, F.; Ferrer, P. Expression of a Rhizopus oryzae lipase in Pichia pastoris under control of the nitrogen source-regulated formaldehyde dehydrogenase promoter. J. Biotechnol. 2004, 109, 103–113. [Google Scholar] [CrossRef]
- Xiong, Z.Y.; Xu, D.; Guo, Y.X.; Tian, X.W.; Chu, J. Establishment of a High-Throughput Screening Method for Rhizomucor miehei Lipase Production by A. oryzae. J. East. China Univ. Sci. Technol. 2022, 48, 519–525. [Google Scholar]
- Chen, Y.; Lin, Y.M.; Tian, X.W.; Li, Q.H.; Chu, J. Real-time dynamic analysis with low-field nuclear magnetic resonance of residual oil and sophorolipids concentrations in the fermentation process of Starmerella bombicola. J. Meth. 2019, 157, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. A New High-Throughput Screening Method for Lipase and the Immobilization and Application of Rhizomucor miehei Lipase. Master’s Thesis, Zhejiang University, Hangzhou, China, 2012. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Tang, W.; Xia, J.; Chu, J.; Zhuang, Y.; Zhang, S. Development and application of morphological analysis method in Aspergillus niger fermentation. Chin. J. Biotechnol. 2015, 31, 291–299. [Google Scholar]
- Gibbs, P.A.; Seviour, R.J.; Schmid, F. Growth of filamentous fungi in submerged culture: Problems and possible solutions. J. Crit. Rev. Biotechnol. 2000, 20, 17–48. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, W.; Yu, H. Enhanced activity of Rhizomucor miehei lipase by deglycosylation of its propeptide in Pichia pastoris. Curr. Microbiol. 2014, 68, 186–191. [Google Scholar] [CrossRef]
- Papagianni, M.; Mattey, M. Morphological development of Aspergillus niger in submerged citric acid fermentation as a function of the spore inoculum level. Microb. Cell Fact. 2006, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Ken, M.; Akira, Y.; Silai, Z.; Motoaki, S.; Mayumi, N.; Katsuya, G.; Keietsu, A. Increased enzyme production under liquid culture conditions in the industrial fungus Aspergillus oryzae by disruption of the genes encoding cell wall α-1,3-glucan synthase. Biosci. Biotechnol. Biochem. 2016, 9, 1853–1863. [Google Scholar]
- Kurakake, M.; Hirotsu, S.; Shibata, M. Relationship between pellet formation by Aspergillus oryzae strain KB and the production of β-fructofuranosidase with high transfructosylation activity. Fungal Biol. 2020, 8, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Hikaru, I.; Ken, M.; Keisuke, K.; Shunya, S.; Silai, Z.; Kiyoaki, M.; Ryutaro, O.; Ayumu, T.; Yuka, K.; Masahiro, H.; et al. Improved recombinant protein production in Aspergillus oryzae lacking both α-1,3-glucan and galactosaminogalactan in batch culture with a lab-scale bioreactor. J. Biosci. Bioeng. 2022, 1, 39–45. [Google Scholar]
- Gao, Q.; Liu, J.; Liu, L.M.; Chen, J. Effect of Morphology of Aspergillus terreus on Itaconic Acid Production Effciency. Chin. J. Process. Eng. 2013, 13, 281–286. [Google Scholar]
- Yu, X.-W.; Lu, X.; Zhao, L.-S.; Xu, Y. Impact of NH4+ nitrogen source on the production of Rhizopus oryzae lipase in Pichia pastoris. Process. Biochem. 2013, 48, 1462–1468. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spore Concentration (spores/Ml) | Residual Sugar (g/L) | DCW (g/L) | Morphology |
|---|---|---|---|
| 1 × 104 | 19.9 ± 0.3 | 7.8 ± 0.3 | Larger pellets (S1) |
| 1 × 105 | 18.0 ± 0.5 | 8.8 ± 0.2 | Radiate mycelium pellets (S2) |
| 1 × 106 | 12.6 ± 0.4 | 13.3 ± 0.2 | Clustered mycelium (S3) |
| 1 × 107 | 10.4 ± 0.2 | 14.2 ± 0.2 | Dispersed mycelia (S4) |
| Spore Concentration (spores/mL) | RML Activity (U/mL) | Residual Sugar (g/L) | DCW (g/L) | Concentration of Mycelium Pellets (pellets/mL) | Average Diameter (mm) |
|---|---|---|---|---|---|
| 1 × 104 | 106.9 ± 1.3 | 1.9 ± 0.1 | 18.1 ± 0.4 | 22 ± 2.2 | 2.99 ± 0.17 |
| 1 × 105 | 209.0 ± 0.7 | 1.7 ± 0.1 | 26.7 ± 0.4 | 1495 ± 5.0 | 0.79 ± 0.02 |
| 1 × 106 | 181.1 ± 0.9 | 1.5 ± 0.1 | 27.7 ± 0.3 | 1874 ± 14.7 | 0.64 ± 0.02 |
| 1 × 107 | 151.7 ± 0.9 | 1.4 ± 0.1 | 28.3 ± 0.1 | 2145 ± 77.2 | 0.54 ± 0.13 |
| Spore Concentration (spores/mL) | Average Diameter (mm) | Concentration of Mycelium Pellets (pellets/mL) | RML Activity (U/mL) |
|---|---|---|---|
| 7 × 104 | 1.05 ± 0.1 | 1030 ± 9.1 | 164.6 ± 1.0 |
| 9 × 104 | 0.81 ± 0.1 | 1232 ± 43.0 | 166.4 ± 0.7 |
| 3 × 105 | 0.77 ± 0.1 | 1516 ± 45.5 | 217.5 ± 1.1 |
| 5 × 105 | 0.72 ± 0.1 | 1605 ± 10.1 | 204.4 ± 1.2 |
| Mode 1 | Mode 2 | |||
|---|---|---|---|---|
| Time (h) | Pellet Concentrations (pellets/mL) | Average Diameter (mm) | Pellet Concentrations (pellets/mL) | Average Diameter (mm) |
| 12 | 270 ± 4 | 0.97 ± 0.01 | 262 ± 2 | 0.98 ± 0.02 |
| 24 | 60 ± 2 | 1.80 ± 0.12 | 71 ± 3 | 1.77 ± 0.12 |
| 48 | 122 ± 6 | 2.35 ± 0.11 | 174 ± 5 | 1.92 ± 0.11 |
| 72 | 206 ± 2 | 1.98 ± 0.10 | 264 ± 3 | 1.66 ± 0.13 |
| 144 | 994 ± 4 | 1.01 ± 0.01 | 1072 ± 2 | 0.97 ± 0.02 |
| Total RML activity (U) | 377,466 ± 1793 | 487,179 ± 2812 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Xu, D.; Xiong, Z.; Yang, Y.; Tian, G.; Wu, X.; Wang, Y.; Zhuang, Y.; Chu, J.; Tian, X. Optimization of the Fermentative Production of Rhizomucor miehei Lipase in Aspergillus oryzae by Controlling Morphology. Bioengineering 2022, 9, 610. https://doi.org/10.3390/bioengineering9110610
Li C, Xu D, Xiong Z, Yang Y, Tian G, Wu X, Wang Y, Zhuang Y, Chu J, Tian X. Optimization of the Fermentative Production of Rhizomucor miehei Lipase in Aspergillus oryzae by Controlling Morphology. Bioengineering. 2022; 9(11):610. https://doi.org/10.3390/bioengineering9110610
Chicago/Turabian StyleLi, Chao, Dou Xu, Zhiyue Xiong, Yiming Yang, Guiwei Tian, Xuezhi Wu, Yonghong Wang, Yingping Zhuang, Ju Chu, and Xiwei Tian. 2022. "Optimization of the Fermentative Production of Rhizomucor miehei Lipase in Aspergillus oryzae by Controlling Morphology" Bioengineering 9, no. 11: 610. https://doi.org/10.3390/bioengineering9110610
APA StyleLi, C., Xu, D., Xiong, Z., Yang, Y., Tian, G., Wu, X., Wang, Y., Zhuang, Y., Chu, J., & Tian, X. (2022). Optimization of the Fermentative Production of Rhizomucor miehei Lipase in Aspergillus oryzae by Controlling Morphology. Bioengineering, 9(11), 610. https://doi.org/10.3390/bioengineering9110610