


Storage Potential of the Cactus Pear (Opuntia ficus-indica) Fruit Juice and Its Biological and Chemical Evaluation during Fermentation into Cactus Pear Wine

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Selection of the Cactus Pear Fruits
2.2. Extraction of Cactus Pear Juice
2.3. Fermentation and Maturation
2.4. Sampling
2.5. Microbiological Analysis
2.6. Isolation of Yeast and Bacteria
2.7. Microbial Identification
2.8. Analysis of the Chemical Parameters
2.8.1. Sugar Content
2.8.2. Ethanol Concentration
2.8.3. Titratable Acidity
2.8.4. Juice and Wine pH
2.8.5. Tannin Content
2.8.6. Mineral Content
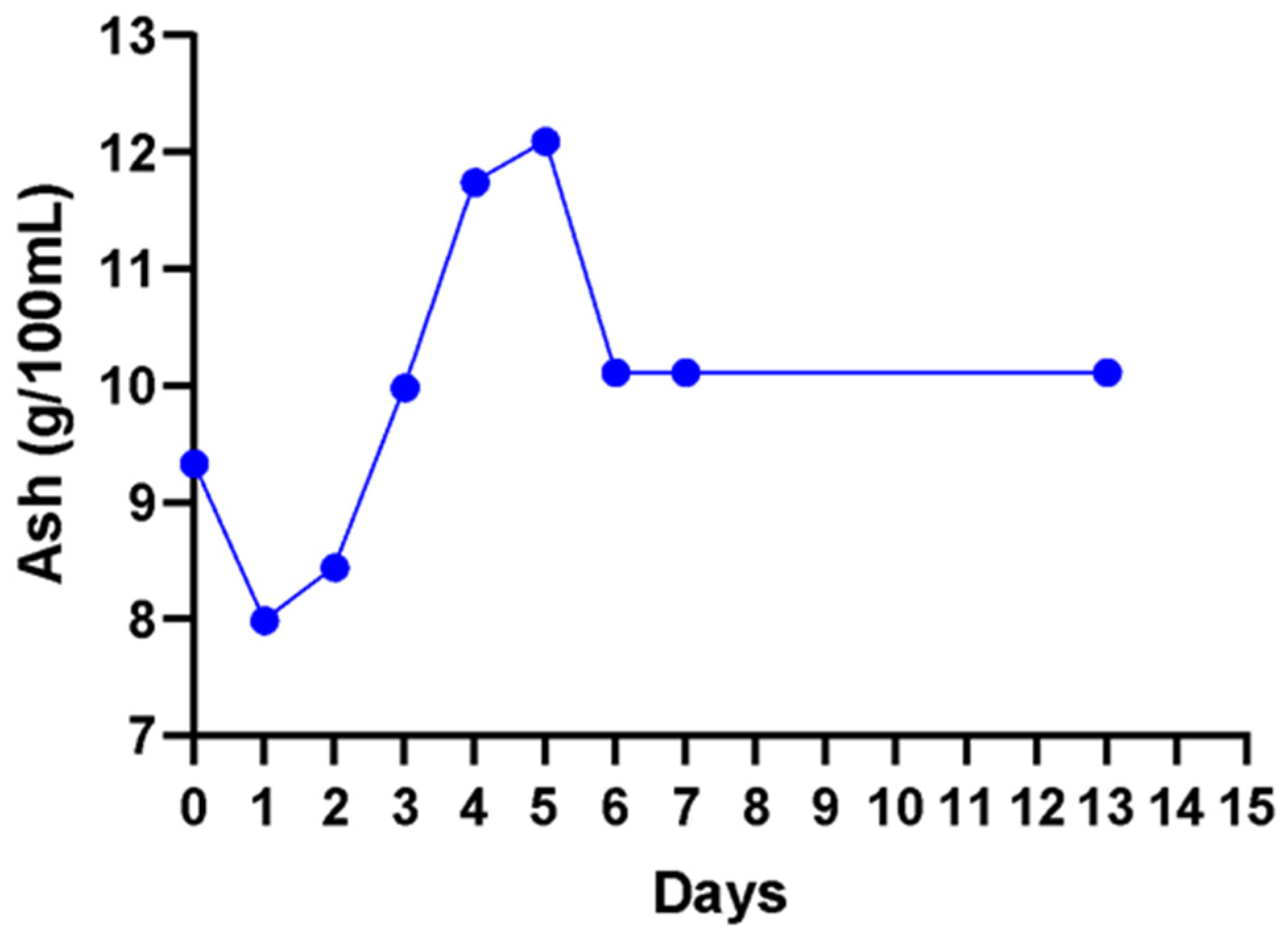
2.8.7. Ash Content
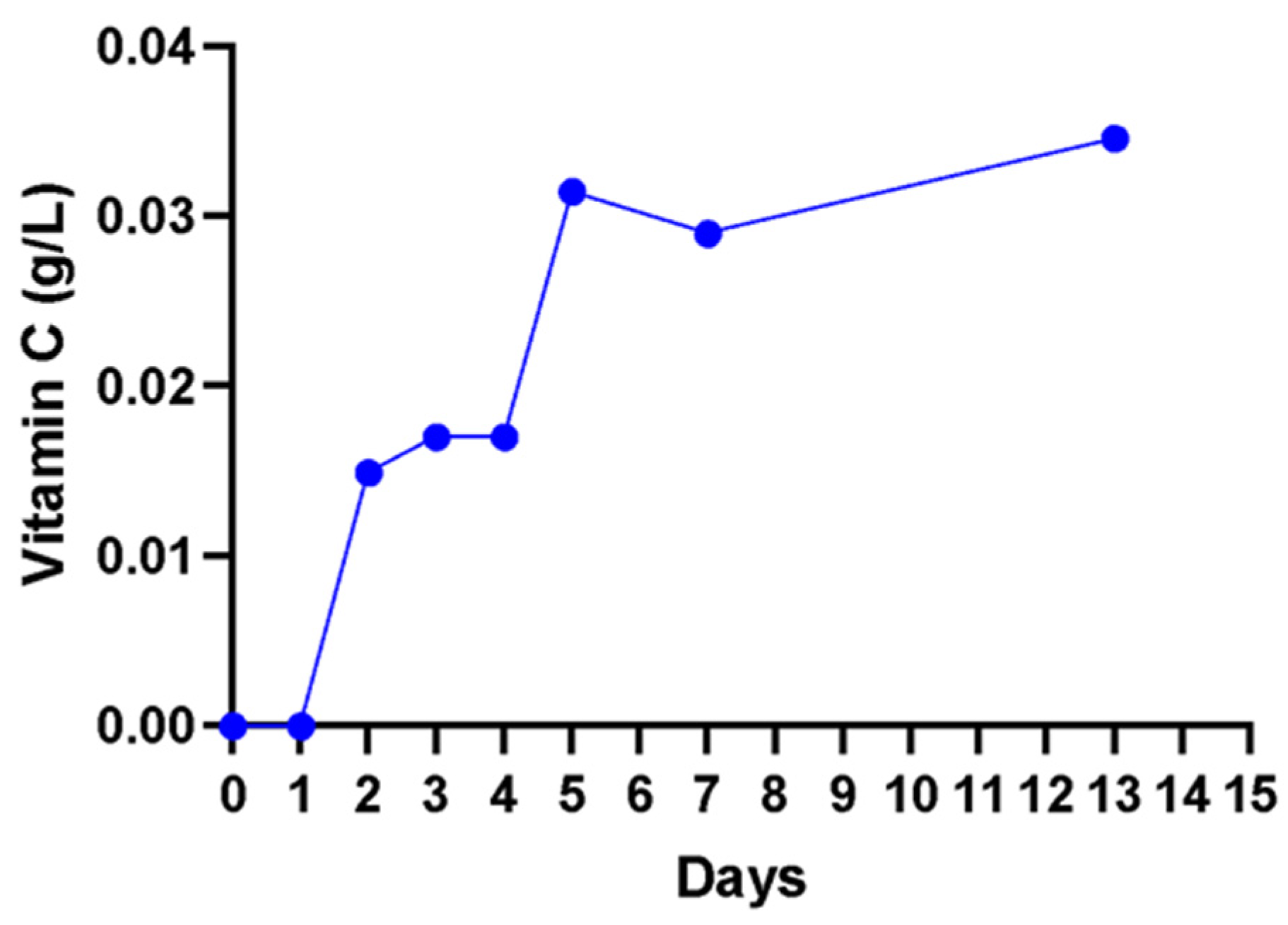
2.8.8. Determination of Vitamin C (Ascorbic Acid)
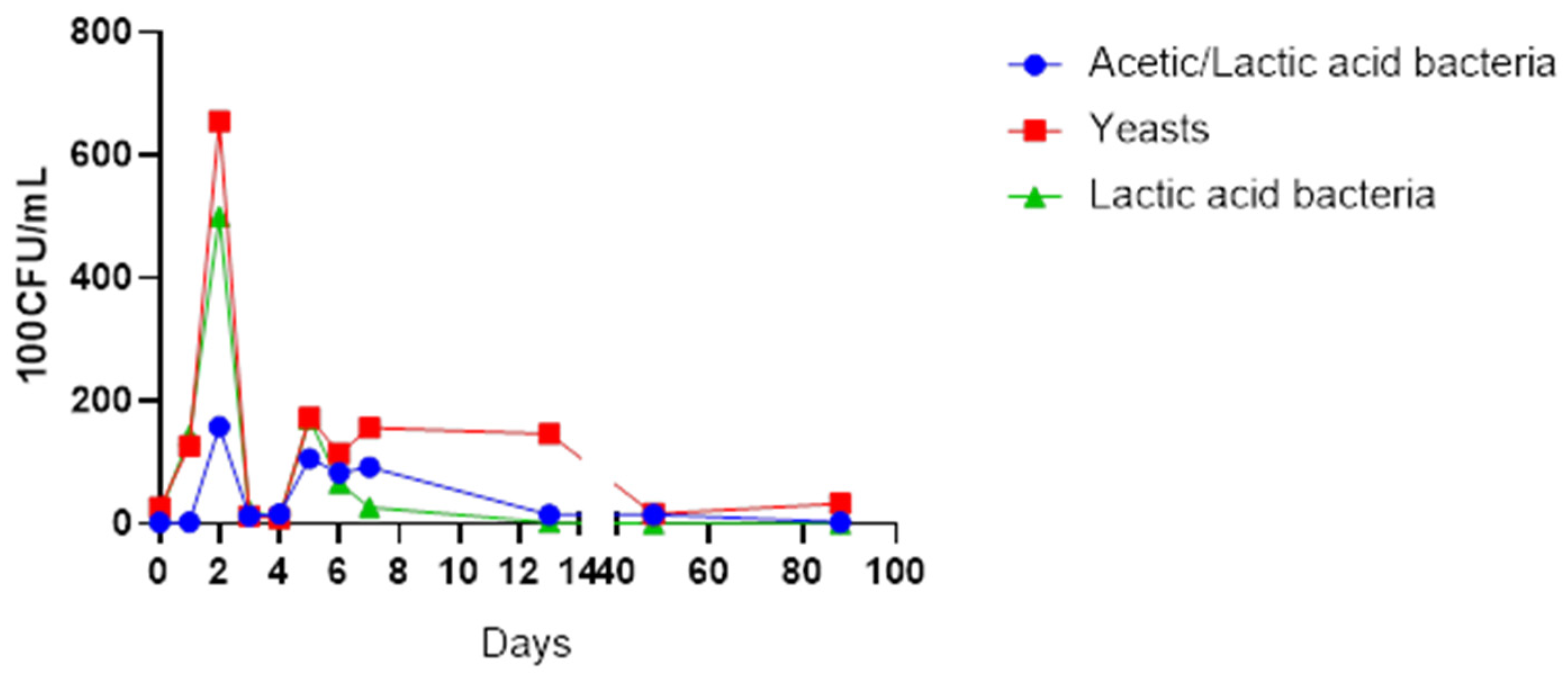
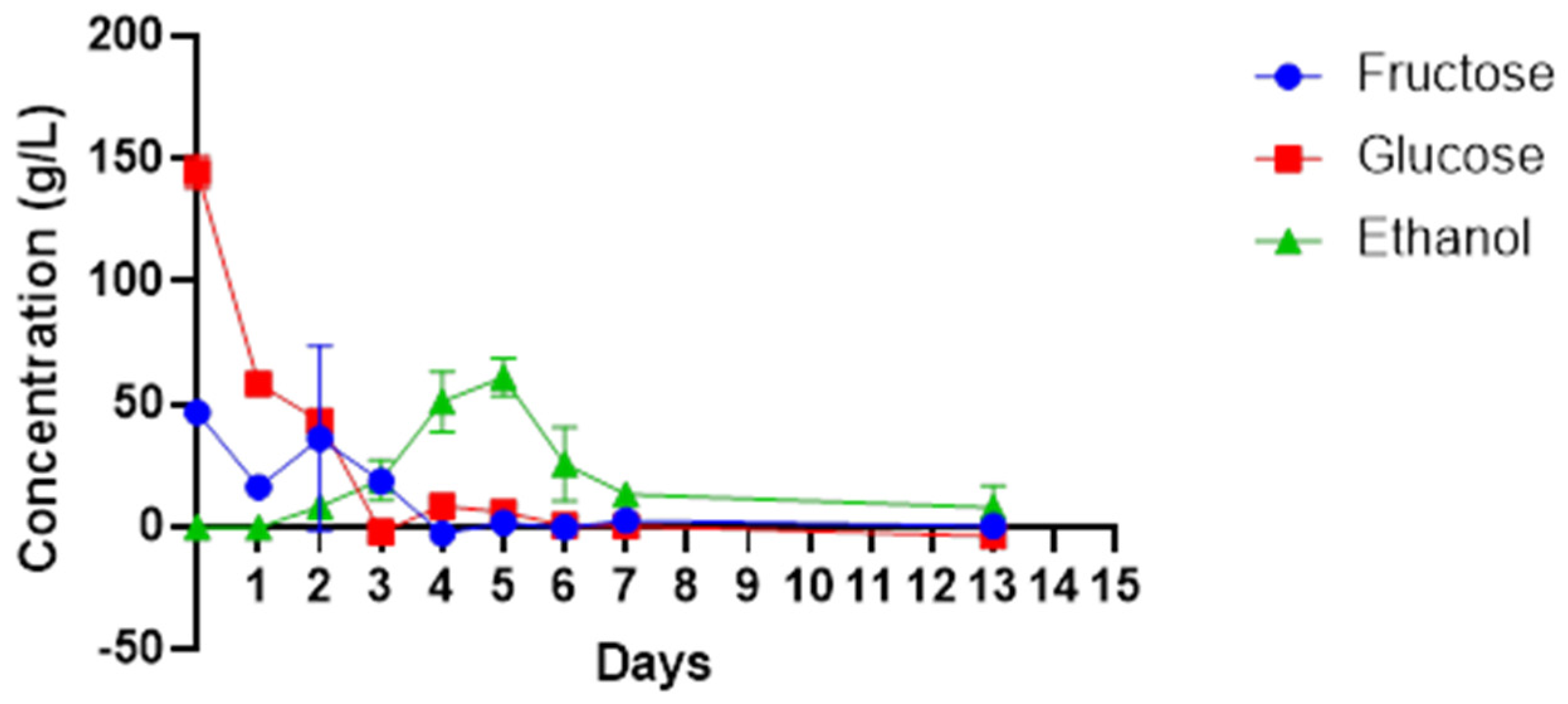
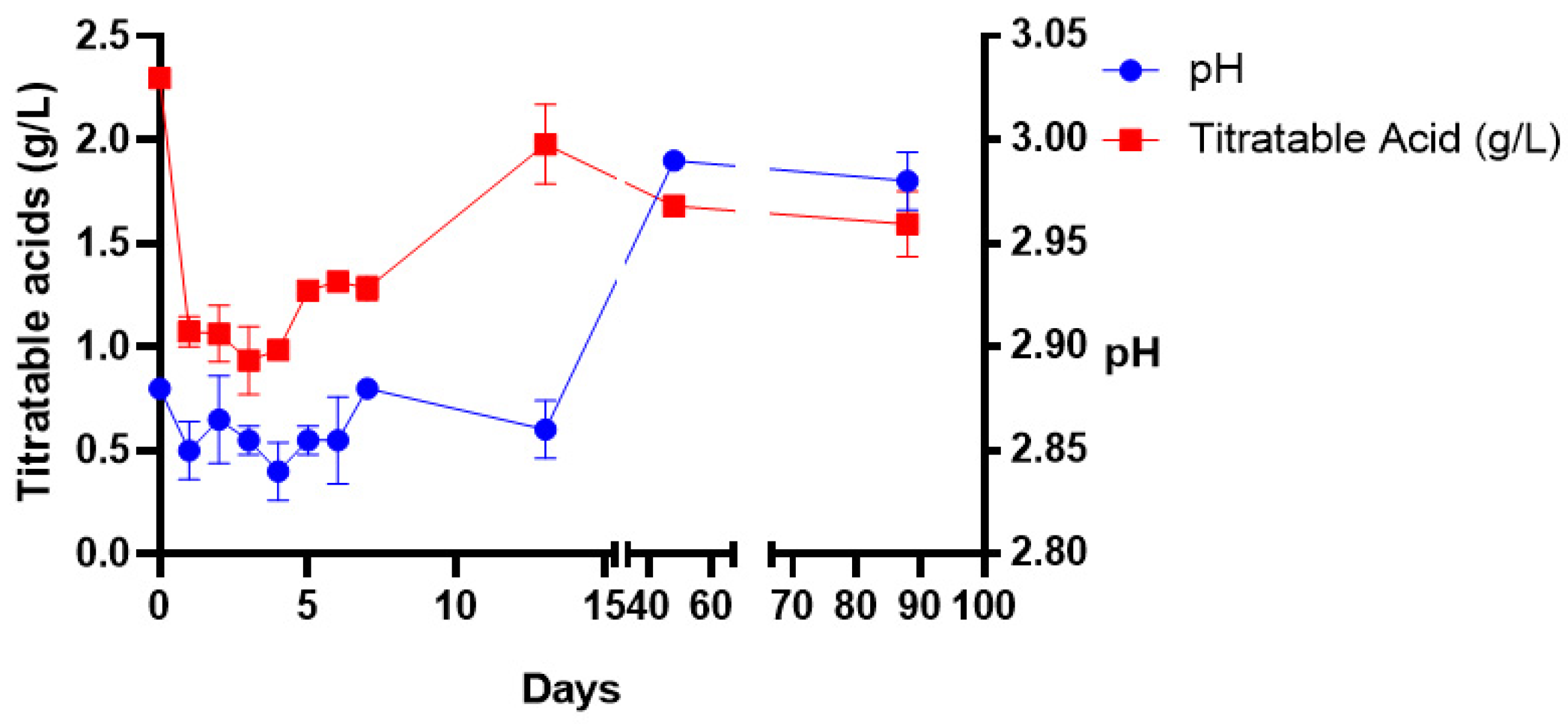
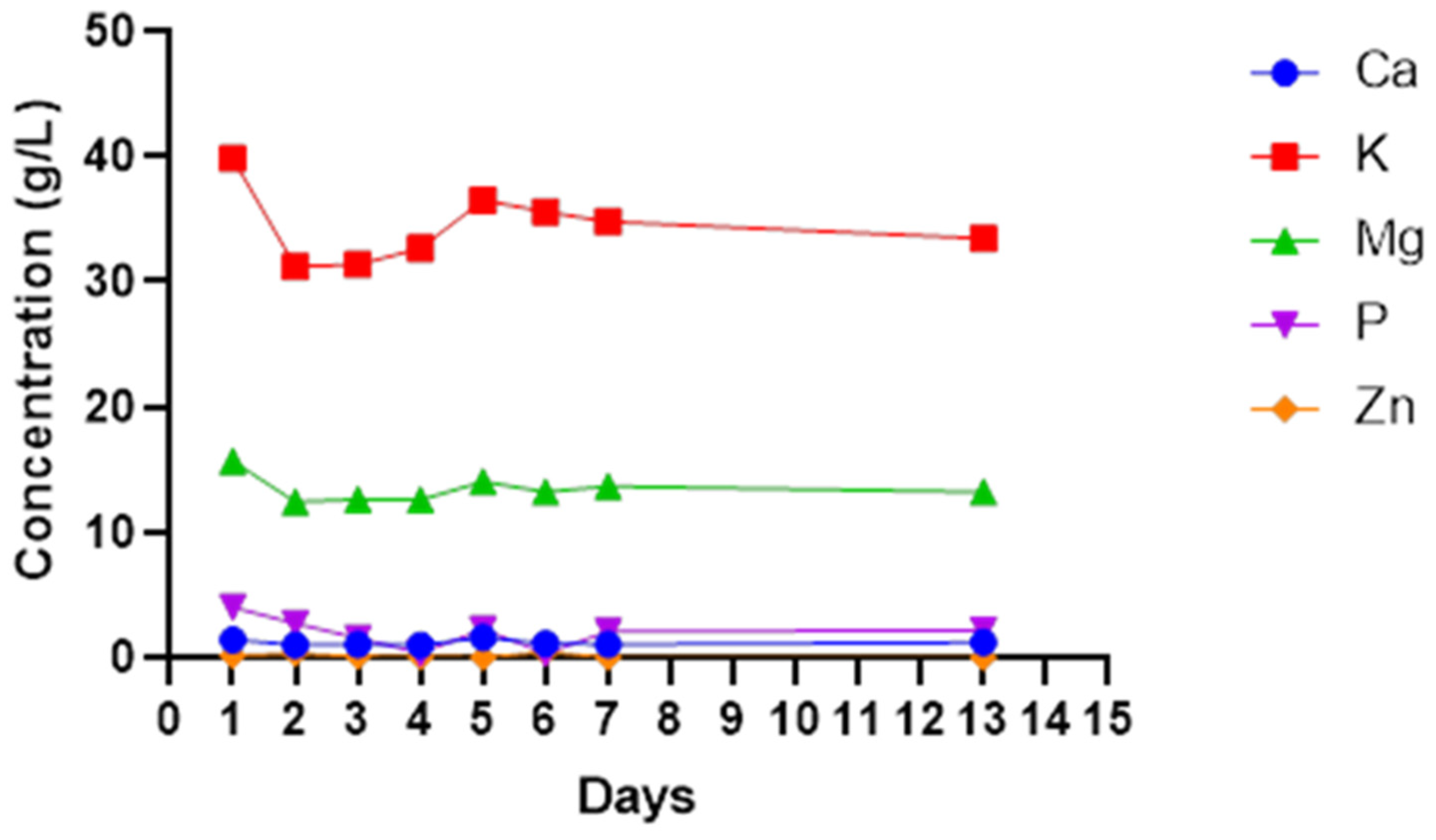
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrios, E.; Zannudo, J.; Yepez, E.; Nobel, P.S. Seasonal variation of net CO2 uptake for cactus pear (Opuntia ficus-indica) and pitayo (Stenocereus queretaroensis) in a semi-arid environment. J. Arid Environ. 2000, 44, 73–83. [Google Scholar] [CrossRef]
- Agriculture Research Council. Prickly Pear (Opuntia Ficus-Indica) (Cactaceae); ARC: Pretoria, South Africa, 2014; Available online: https://www.arc.agric.za/arc-ppri/Pages/Prickly-pear.aspx (accessed on 15 April 2022).
- Sáenz, C.; Sepúlveda, E. Cactus-Pear Juices. J. PACD 2001, 4, 3–10. [Google Scholar]
- Zimmermann, H.G.; Moran, C.V. Biological control of prickly pear, Opuntia ficusindica (Cactaceae), in South Africa. Agric. Ecosyst. Environ. 1991, 37, 29–35. [Google Scholar] [CrossRef]
- Oberoi, H.S.; Vandlani, P.V.; Nanjundaswamy, A.; Bansal, S.; Sigh, S.; Kaur, S.; Babbar, N. Enhanced ethanol production from Kinnow mandarin (Citrus reticulata) waste via a statistically optimized simultaneous saccharification and fermentation process. Bioresour. Technol. 2011, 102, 1593–1601. [Google Scholar] [CrossRef]
- Peng, B.; Lei, Y.; Zhao, H.; Cui, L. Response surface methodology for optimization of fermentation process parameters for improving apple wine quality. J. Food Sci. Technol. 2015, 52, 513–751. [Google Scholar] [CrossRef]
- Kumar, S.Y.; Prakasam, R.S.; Reddy, O.V.S. Optimization of fermentation conditions for mango (Mangifera indica L.) wine production by employing response surface methodology. Int. J. Food Sci. 2009, 44, 2320–2327. [Google Scholar] [CrossRef]
- Navarrete-Bolanos, J.L. Improving traditional beverages: How to evolve from spontaneous to directed fermentation. Eng. life Sci. 2012, 12, 410–418. [Google Scholar] [CrossRef]
- Achi, O.K. The potential for upgrading traditional fermented foods through biotechnology. Afr. J. Biotechnol. 2005, 4, 375–380. [Google Scholar]
- Rodriguez-Lerma, G.K.; Gutiérrez-Moreno, K.; Cardenas-Manrıquez, M.; Botello-Álvarez, E.; Jiménez-Islas, H.; Rico-Martınez, R.; Navarrete-Bolanos, J.L. Microbial ecology studies of spontaneous fermentation: Starter culture selection for prickly pear wine production. J. Food Sci. 2011, 76, 346–352. [Google Scholar] [CrossRef]
- Navarrete-Bolaños, J.L.; Fato-Aldeco, E.; Gutiérrez-Moreno, K.; Botello-Álvarez, E.; Jiménez-Islas, H.; Rico-Martínez, R. A strategy to design efficient fermentation processes for traditional beverages production: Prickly pear wine. J. Food Sci. 2013, 78, 1560–1568. [Google Scholar] [CrossRef]
- Tsegay, Z.T.; Sathyanarayana, B.C.; Lemma, M.S. Optimization of cactus pear fruit fermentation process for wine production. Foods 2018, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kossori, E.L.; Villaume, R.L.; Boustani, E.L. The effect of variety and location on cactus pear (Opuntia ficus-indica) fruit quality. Plant Foods Hum. Nutr. 2004, 52, 263. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, W.N.; Khalil, J.K.; Al-Mohammad, M.M. Nutritive value of prickly pear seeds, Opuntia ficus-indica. Plant Foods Hum. Nutr. 1983, 33, 91–97. [Google Scholar] [CrossRef]
- Corrales-García, J.; Esparza- Torres, F.; Jiménez-Montoya, D.; Miranda-Romero, L.A.; del Rosario García-Mateos, M. Quality and safety of wine made from intact and peeled fruits of cactus pear (Opuntia spp.). Acta Hortic. 2015, 14, 1067. [Google Scholar] [CrossRef]
- Sáenz, C.; Berger, H.; Rodríguez-Félix, A.; Galletti, L.; García, J.C.; Sepúlveda, E.; Varnero, M.T.; de Cortázar, V.G.; García, R.C.; Arias, E.; et al. Agro-Industrial Utilization of Cactus Pear; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; pp. 1–124. [Google Scholar]
- Gebremedhin, K.; Hailu, Z.; Dargo, H.; Gebrehiwot, H. Preparation of wine from cactus fruit in Adigrat rural areas. Int. J. Eng. Res. Dev. 2017, 7, 292–298. [Google Scholar]
- Galati, E.M.; Mondello, M.R.; Giuffrida, D.; Dugo, G.; Miceli, N.; Pergolizzi, S.; Taviano, M.F. Chemical characterization and biological effects of Sicilian Opuntia ficus indica (L.) Mill. fruit juice: Antioxidant and antiulcerogenic activity. J. Agric. Food Chem. 2003, 51, 4903–4908. [Google Scholar] [CrossRef]
- Medina, E.D.; Rodriguez, E.R.; Romero, C.D. Chemical characterization of Opuntia dillenii and Opuntia ficusidica. Food Chem. 2007, 103, 38–45. [Google Scholar] [CrossRef]
- Akanni, G.; Ntuli, V.; du Preez, J.C. Cactus pear biomass, a potential lignocellulose raw material for single cell protein production (SCP): A review. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 171–197. [Google Scholar]
- Firew, T.; Peters, K.J.; Kijora, C. Current uses of cactus pear (Opuntia ficus-indica) as forage in Northern Ethiopia and farmers’ indigenous knowledge on its utilisation. Ethiop. J. Vet. Sci. Anim. Prod. 2008, 8, 10–21. [Google Scholar]
- Kelebek, H.; Selli, S.; Canbas, A.; Cabaroglu, T. HPLC determination of organic acids, sugars, phenolic compositions and antioxidant capacity of orange juice and orange wine made from a Turkish cv. Kozan. Microcheml. J. 2009, 91, 187–192. [Google Scholar] [CrossRef]
- Tambe, V.D.; Bhambar, R.S. Estimation of total phenol, tannin, alkaloid and flavonoid in Hibiscus tiliaceus linn. Wood extracts. Res. Rev. J. Pharmacogn. Phytochem. 2014, 2, 41–47. [Google Scholar]
- Dlamini, N.R.; Dube, S. Studies on the physico-chemical, nutritional and microbiological changes during the traditional preparation of marula wine in Gwanda, Zimbabwe. Nutr. Food Sci. 2008, 38, 61–69. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Bamforth, C.W. The microbiology of malting and brewing. Microbiol. Mol. Biol. Rev. 2013, 77, 57–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Garcia, S.; Hernández-Bustos, C.; Merino, E.; Gosset, G.; Martinez, A. Homolactic fermentation from glucose and cellobiose using Bacillus subtilis. Microb. Cell Fact. 2009, 8, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Torija, M.J.; Rozès, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Yeast population dynamics in spontaneous fermentations: Comparison between two different wine-producing areas over a period of three years. Antonie Leeuw Int J. Gen. 2001, 79, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Amoa-Awa, W.K.A.; Tukobsen, K. The role of Bacillus species in the fermentation of cassava. J. Appl. Microbiol. 2008, 79, 250–256. [Google Scholar]
- Gao, T.; Wong, Y.; Ng, C.; Ho, K. L-Lactic acid production by Bacillus subtilis MUR1. Bioresour. Technol. 2009, 121, 105–110. [Google Scholar] [CrossRef]
- Rantsiou, K.; Englezos, V.; Torchio, F.; Risse, P.A.; Cravero, F.; Gerbi, V.; Rolle, L.; Cocolin, L. Modeling of the fermentation behavior of Starmerella bacillaris. Am. J. Enol. Vitic. 2017, 68, 378–385. [Google Scholar] [CrossRef]
- Jolly, N.; Mehlomakulu, N.N.; Nortje, S.; Beukes, L.; Hoff, J.; Booyse, M.; Erten, H. Non-Saccharomyces yeast for lowering wine alcohol levels: Partial aeration versus standard conditions. FEMS Yeast Res. 2022, 22, foac002. [Google Scholar] [CrossRef]
- Peres, C.M.; Alves, M.; Hernandez-Mendoza, A.; Moreira, L.; Silva, S.; Bronze, M.Z.; Vilas-Boas, L.; Peres, C.; Malcataef, F.X. Novel isolates of Lactobacilli from fermented Portuguese olive as potential probiotics. Food Sci. Technol. 2014, 59, 234–246. [Google Scholar] [CrossRef] [Green Version]
- Nissen, P.; Nielsin, D.; Arneborg, N. Viable Saccharomyces cerevisiae cells at high concentrations cause early growth arrest of non-Saccharomyces yeasts in mixed cultures by a cell-cell contact-mediated mechanism. Yeast 2003, 20, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisson, L.F.; Kunkee, R. Microbial interactions during wine production. In Mixed Cultures in Biotechnology; Zeikus, J.G., Johnson, E.A., Eds.; McGraw-Hill: New York, NY, USA, 1991; pp. 37–68. [Google Scholar]
- Lema, C.; Garcia-Jares, C.; Orriols, I.; Angulo, L. Contribution of Saccharomyces and non-Saccharomyces populations to the production of some compounds of Albarino wine aroma. Am. J. Enol. Vitic. 1996, 47, 206–216. [Google Scholar]
- Furlani, M.R.V.; Maturano1, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of non-Saccharomyces yeasts to be used in grape musts with high alcoholic potential: A strategy to obtain wines with reduced ethanol content. FEMS Yeast Res. 2017, 17, 1–10. [Google Scholar]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, W.J.; Pretorius, I.S. The occurrence, control and esoteric effect of acetic acid bacteria in winemaking. Ann. Microbiol. 2002, 52, 155–179. [Google Scholar]
- Priyodip, P.; Prakash, P.Y.; Balaji, S. Phytases of probiotic bacteria: Characteristics and beneficial aspects. Indian J. Microbiol. 2017, 57, 148–154. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P. The ‘buttery’ attribute of wine–diacetyl-desirability, spoilage and beyond. Int. J. Food Microbiol. 2004, 96, 235–252. [Google Scholar] [CrossRef]
- Plevri, N.; Demertzis, P.G.; Akrida-Demertzi, K. Microbiological evaluation of fruits of the prickly pear (Opuntia ficus-indica L. Miller) Collected in Greece. IOSR-JAVS 2017, 10, 31–38. [Google Scholar]
- Veron, H.E.; Di Risio, H.D.; Isla, M.I.; Torres, S. Isolation and selection of potential probiotic lactic acid bacteria from Opuntia ficus-indica fruits that grow in Northwest Argentina. LWT-Food Sci. 2017, 10, 231–240. [Google Scholar] [CrossRef]
- Carpena, M.; Fraga-Corral, M.; Otero, P.; Nogueira, R.A.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Secondary a roma: Influence of wine microorganisms in their aroma profile. Foods 2020, 10, 51. [Google Scholar] [CrossRef]
- Ayed, L.; Hamdi, M. Manufacture of a beverage from cactus pear juice using “tea fungus” fermentation. Ann. Microbiol. 2015, 65, 2293–2299. [Google Scholar] [CrossRef]
- Gurrier, S.; Miceli, L.; Lanza, C.M.; Tomaselli, F.; Bonomo, R.P.; Rizzarelli, E. Chemical characterization of Sicilian prickly pear (Opuntia ficus indica) and perspectives for the storage of its juice. J. Agric. Food Chem. 2000, 48, 5424–5431. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Christmann, M. The use of dextrose in winemaking. EDP Sci. 2016, 31, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J. Ethanol from sugar: What are the prospects for US sugar crops? Rural Coop. 2006, 73, 1–46. [Google Scholar]
- Tsegay, Z.T.; Gebremedhin, M.K. Physicochemical and sensory properties of wine produced from blended cactus pear (Opuntia ficus-indica) and Lantana camara (L. camara) Fruits. J. Food Qual. 2019, 2019, 6834946. [Google Scholar] [CrossRef] [Green Version]
- Alba-Lois, L.; Segal-Kischinevzky, C. Beer & Wine Makers. Nat. Sci. Educ. 2010, 3, 1–17. [Google Scholar]
- Lund, P.A.; Biase, D.D.; Liran, O.; Scheler, O.; Mira, N.P.; Cetecioglu, Z.; Fernandez, E.N.; Bover-Cid, S.; Hall, R.; Sauer, M.; et al. Understanding how microorganisms respond to acid pH is central to their control and successful exploitation. Front. Microbiol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Rajkovic, M.B.; Novakovic, I.D.; Petrovic, A. Determination of titratable acidity in white wine. J. Agric. Sci. 2007, 52, 169–184. [Google Scholar] [CrossRef]
- Barnett, J.A.; Lichtenthaler, F.W. A history of research on yeast 3: Emil Fischer, Eduard Buchner and their contemporaries. Yeast 2001, 18, 363–388. [Google Scholar] [CrossRef]
- Hillman, Z.; Mizrahi, Y.; Beit-Yannai, E. Evaluation of valuable nutrients in selected genotypes of marula (Sclerocarya birrea ssp. caffra). Sci. Hortic. 2008, 117, 321–328. [Google Scholar] [CrossRef]
- Moganedi, K.L.M.; Glober, J.P.; Goyvaerts, E.; Sibara, M. An assessment of genetic diversity among marula populations using the amplified fragment length polymorphism (AFLP) technique. Afr. J. Agric. Res. 2011, 6, 790–797. [Google Scholar]
- Harbertson, J.F.; Hodgins, R.E.; Thurston, L.N.; Schaffer, L.J.; Reid, M.S.; Landon, J.L.; Ross, C.F.; Adams, D.O. Variability of tannins concentration in red wines. Am. J. Enol. Vitic. 2008, 59, 210–214. [Google Scholar]
- Shang, A.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Tang, G.Y.; Corke, H.; Mavumengwana, V.; Li, H.B. Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiwilepo-van Hal, P.; Bosschaart, C.; Van Twisk, C.; Verkerk, R.; Dekker, M. Kinetics of thermal degradation of vitamin C in marula fruit (Sclerocarya birrea subsp. caffra) as compared to other selected tropical fruits. Food Sci. Technol. 2012, 49, 188–191. [Google Scholar] [CrossRef]
- Mabotja, F.S.; Metcalfe, D.J.A.; Adebo, O.A. South African consumers’ interpretation of nutritional labelling systems of food products. Trans. R. Soc. S. Afr. 2021, 76, 41–51. [Google Scholar] [CrossRef]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Serna-Sald´ıvar, S.O. Phenolic composition, antioxidant capacity and in vitro cancer cell cytotoxicity of nine prickly pear (Opuntia spp.) juices. Plant Foods Hum. Nutr. 2009, 64, 146–152. [Google Scholar] [CrossRef]
- Buitendag, R.A. Yield and Fruit Quality Assessment of Cactus Pear (Opuntia ficus-indica (L.) Mill.) Treated with Natural Bio-Stimulants; University of the Free State: Bloemfontein, South Africa, 2004; pp. 6–24. Available online: https://scholar.ufs.ac.za/bitstream/handle/11660/7688/BuitendagRA.pdf?sequence=1&isAllowed=y (accessed on 27 April 2022).
- Tirna, P.; Sangeeta, P. Formulation and nutrient analysis of vitamin c enriched red wine using roselle (Hibiscus sabdariffa) and peppermint (Mentha piperita L.). Agric. Food Secur. 2018, 1, 58–71. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days | Identified Microbes | Dominant Yeast and Bacteria spp. |
|---|---|---|
| 0 | Aureobasidium pullulans Bacillus amyloliquefaciens Gluconobacter sp. Gluconobacter Kondonii Aureobasidium leucospermi Gluconobacter albidus Starmerella bacillaris Bacillus pumilus | Starmerella and Gluconobacter spp. |
| 1 | Gluconobacter sp. Gluconobacter kondonii Gluconobacter albidus Starmerella bacillaris Staphylococcus epidermidis Starmerella bacillaris Bacillus amyloliquefaciens Bacillus sp. | Starmerella and Gluconobacter spp. |
| 2 | Hanseniaspora opuntiae Hanseniaspora uvarum Pichia kluyveri Pichia fermentans | Hanseniaspora sp. |
| 3 | Starmerella bacillaris Hanseniaspora opuntiae Pichia fermentans Hanseniaspora uvarum Bacillus pumilus Bacillus megaterium Bacillus aryabhattai Bacillus subtilis Bacillus sp. | Hanseniaspora and Bacillus spp. |
| 4 | Hanseniaspora opuntiae Pichia fermentans Pichia kluyveri Hanseniaspora guilliermondi | Hanseniaspora and Pichia spp. |
| 5 | Bacillus subtilis Bacillus amyloliquefaciens | Bacillus sp. |
| 6 | Bacillus thuringiensis Rothia dentocariosa Paenibacillus taichungensis Saccharomyces cerevisiae Bacillus spp. Bacillus velezensis Bacillus subtilis | Saccharomyces sp. |
| 13 | Bacillus subtilis Saccharomyces cerevisiae | Saccharomyces sp. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramoba, L.; Monyama, M.C.; Moganedi, K. Storage Potential of the Cactus Pear (Opuntia ficus-indica) Fruit Juice and Its Biological and Chemical Evaluation during Fermentation into Cactus Pear Wine. Beverages 2022, 8, 67. https://doi.org/10.3390/beverages8040067
Ramoba L, Monyama MC, Moganedi K. Storage Potential of the Cactus Pear (Opuntia ficus-indica) Fruit Juice and Its Biological and Chemical Evaluation during Fermentation into Cactus Pear Wine. Beverages. 2022; 8(4):67. https://doi.org/10.3390/beverages8040067
Chicago/Turabian StyleRamoba, Louis, Maropeng Charles Monyama, and Kgabo Moganedi. 2022. "Storage Potential of the Cactus Pear (Opuntia ficus-indica) Fruit Juice and Its Biological and Chemical Evaluation during Fermentation into Cactus Pear Wine" Beverages 8, no. 4: 67. https://doi.org/10.3390/beverages8040067
APA StyleRamoba, L., Monyama, M. C., & Moganedi, K. (2022). Storage Potential of the Cactus Pear (Opuntia ficus-indica) Fruit Juice and Its Biological and Chemical Evaluation during Fermentation into Cactus Pear Wine. Beverages, 8(4), 67. https://doi.org/10.3390/beverages8040067