
The Effect of Disease and Injury on Faecal Cortisol Metabolites, as an Indicator of Stress in Wild Hospitalised Koalas, Endangered Australian Marsupials

 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Koalas
2.1.1. AZ Koalas
2.1.2. ML Koalas
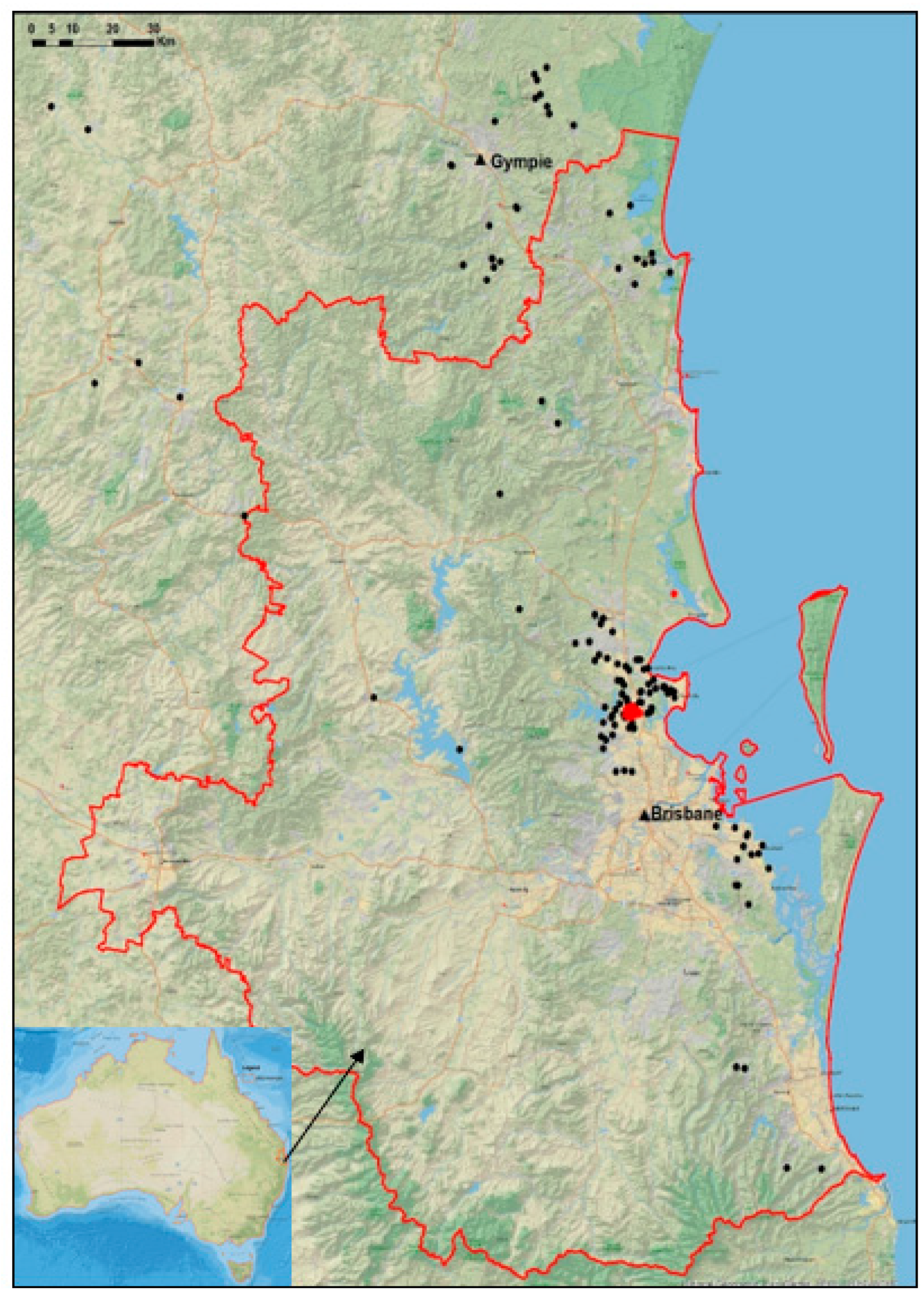
2.2. Location of Koalas
2.3. FCM Extraction and Analysis
2.4. Statistics
3. Results
3.1. FCM Values for the Control Koalas from AZ and ML
3.2. Longitudinal and 2-Day Scat Collection with and without GC Treatment
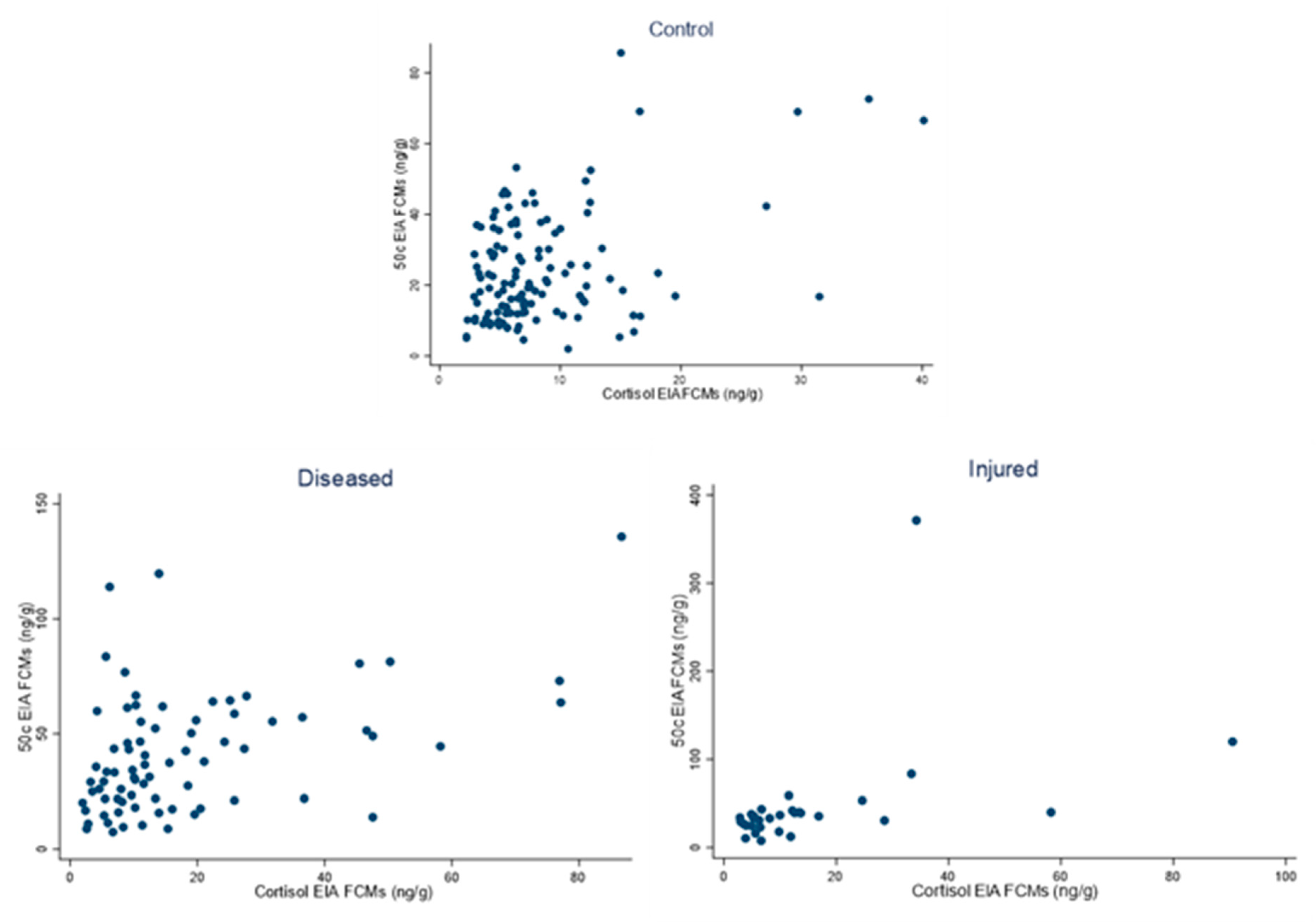
3.3. Correlation between FCM Values Analysed with the 50c and Cortisol EIAs in Day 1 Samples of Control, Diseased and Injured Conditions
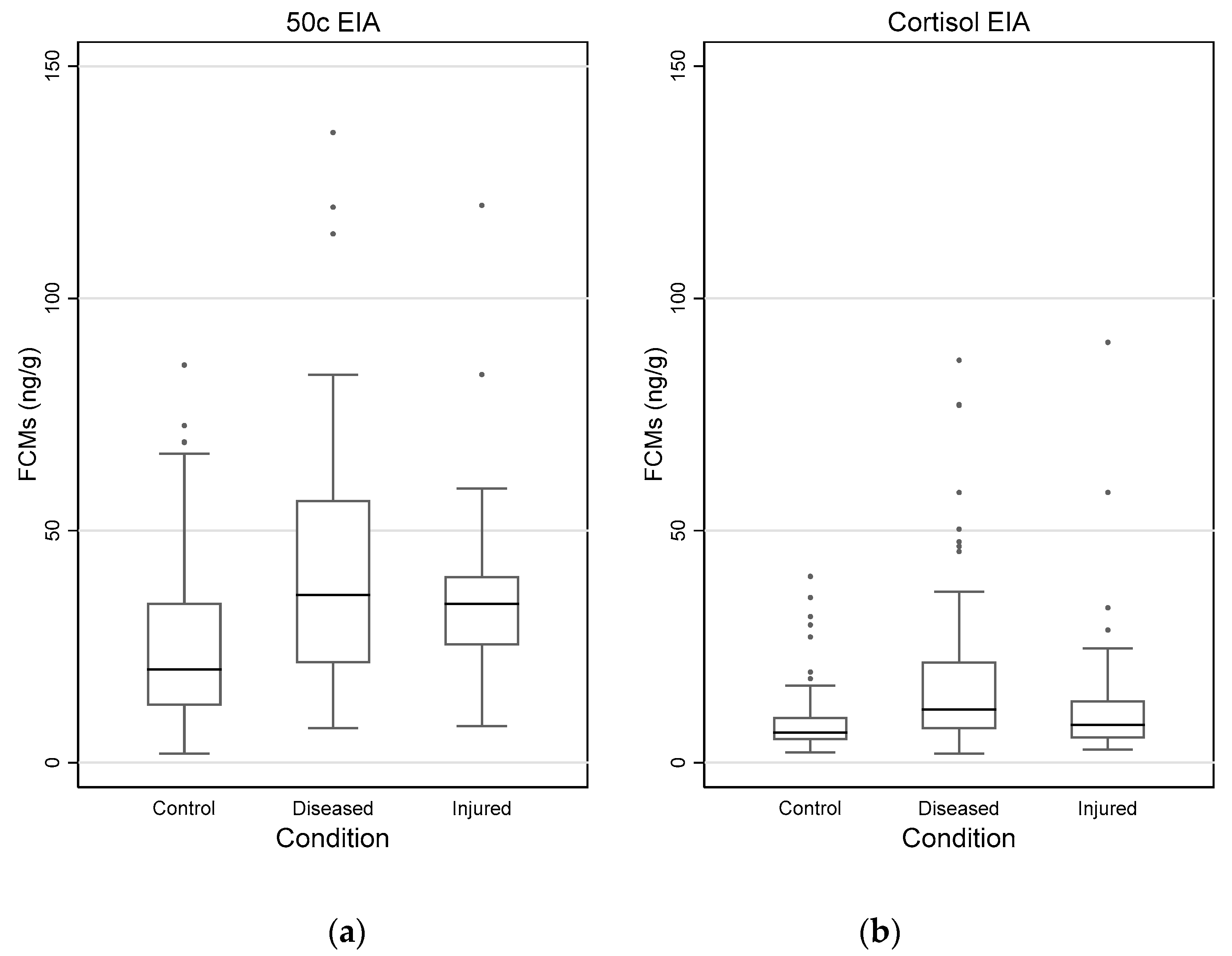
3.4. Impact of Disease and Injuries on FCM Values
3.5. Impact of Disease and Injuries on FCM Values by Sex
4. Discussion
4.1. FCM Values for the Control Koalas from AZ and ML
4.2. Longitudinal and 2-Day Scat Collection with and without GCs Treatment
4.3. Impact of Disease and Injuries on FCM Values
4.4. Impact of Disease and Injuries on FCM Values between Sexes
4.5. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | 50C EIA (ng/g) | Cortisol EIA (ng/g) | |
|---|---|---|---|
| AZ (N = 36) | Min. Max. Mean | 5.5 72.6 26.9 | 2.2 40.1 10.6 |
| ML (N = 88) | Min. Max. Mean | 1.9 85.6 23.2 | 2.2 19.5 7.5 |
| Graphs Reflecting FCM Values | Notes and Correlations |
|---|---|
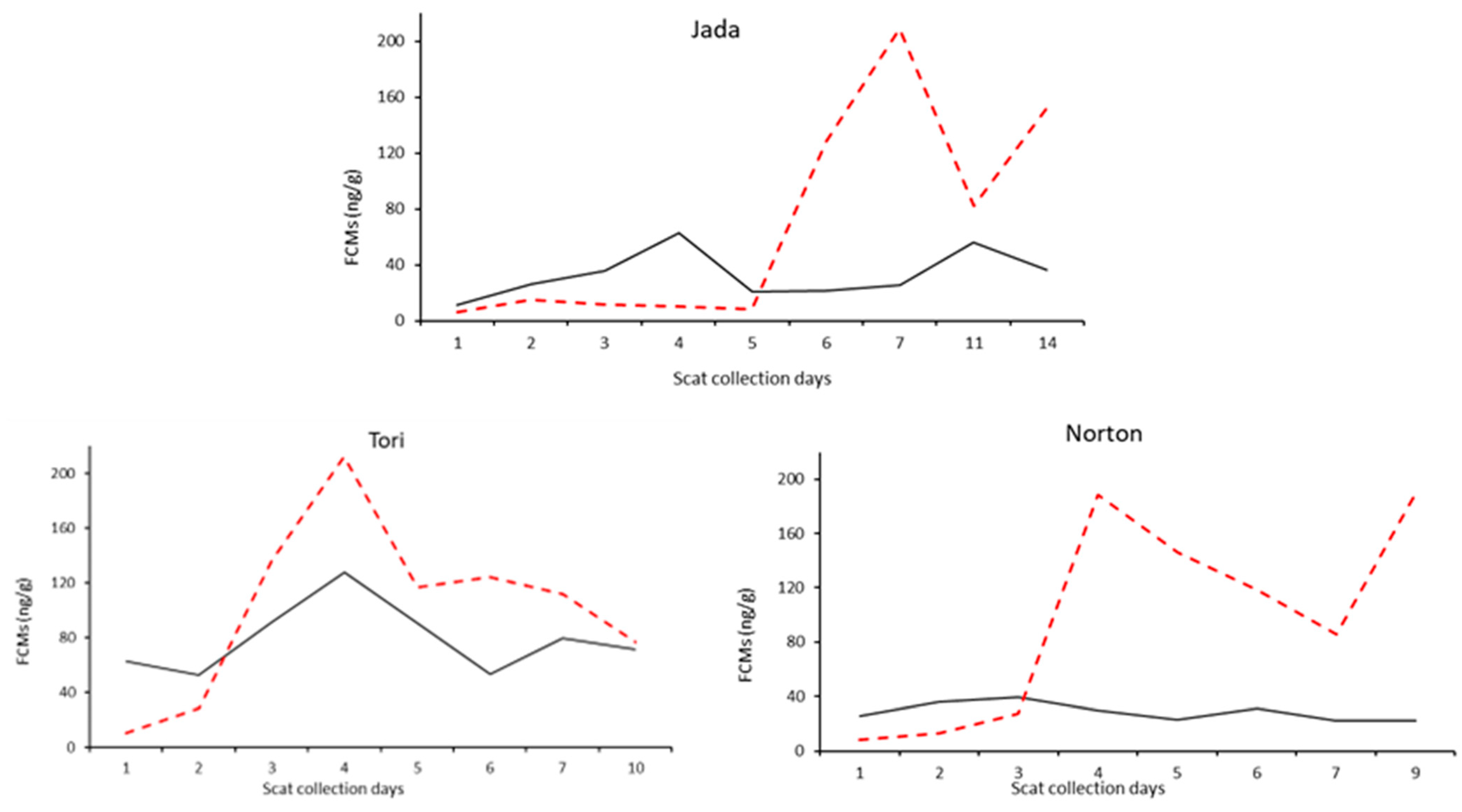
 (a) (a) | Locket (female): control The relatively high values on day 1 are likely related to the hospitalisation the afternoon before day 1. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = −0.0385 (p = 0.9347) |
 (b) (b) | Scratchy (male; orphan): control The high values on day 1 are likely related to the hospitalisation the day before day 1 after she was found alone on the side of a road. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = −0.18442 (p = 0.5818) |
 (c) (c) | Cloyna (male): diseased (cystitis, anaemia and kidney disease) He had extreme values of FCMs at admission due to illness. The spike on day 2 is likely related to the hospitalisation on day 1. He was euthanised after the last faeces collection. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = −0.0019 (p = 0.9962) |
 (d) (d) | Cyril (female): diseased (Chlamydia; LAMP = 13.37 min) The spike on day 2 is likely related to the hospitalisation on day 1. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.1000 (p = 0.8311) |
 (e) (e) | Larissa (female): diseased (Chlamydia; LAMP = 3.56 min) The high values from day 7 are likely related to the start of the injection treatment. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = −0.6866 (p = 0.0600) |
 (f) (f) | Molly (female): diseased (Chlamydia; LAMP = 15.15 min) Treatment started on day 4. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.3144 (p = 0.6064) |
 (g) (g) | Golf (male): diseased (Chlamydia; LAMP = 14.4 min) The spikes on days 2 and 6 are likely related to the procedure for general anaesthetic carried out on days 1 and 5. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.4136 (p = 0.2348) |
 (h) (h) | Clementine (female): diseased (Chlamydia: LAMP = 8.51 min) No specific data were recorded but she was ultimately euthanised. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.8439 (p = 0.0084) |
 (j) (j) | Rarsta (male): diseased (kidney infection) He underwent injection treatment. Rarsta was euthanised 3 days after the last scat collection. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r= 0.4579 (p = 0.2539) |
 (k) (k) | Blake (male): diseased (chlamydial conjunctivitis) The spike on day 4 is likely related to day 3 injection. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r= 0.8270 (p = 0.0113) |
 (l) (l) | Spring (female): diseased (chlamydial conjunctivitis) She was recorded to be highly stressed in her enclosure on day 3. No notes were available to explain the increase on day 7. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.4835 (p = 0.2249) |
 (m) (m) | Wayne-o (male): injured (hit by car) The spike on day 2 is likely related to the hospitalisation on day 1. Procedure for general anaesthetic was carried out on day 5. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = −0.1518 (p = 0.7197) |
 (n) (n) | Firecracker (female): injured (hit by a car) The spike on day 2 is likely related to the procedure for general anaesthetic carried out on day 1. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.4041 (p = 0.2808) |
 (o) (o) | Milko (male): injured (hit by a car) Injection treatment started on day 1. Procedure for general anaesthetic was carried out on days 2 and 4. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.1905 (p = 0.6235) |
 (p) (p) | Farrel (male): injured (abscess on the face and peritonitis) He was administered an injection of antibiotic on day one and carprofen, starting on day 2 for 5 days. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.2169 (p = 0.6403) |
 (q) (q) | Luca (male): injured The high values on day 1 are likely related to the hospitalisation in the afternoon before day 1. No data were recorded that may explain the spike on day 4. The higher values from day 7 are likely related to the procedure for general anaesthetic carried out on day 7. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.3763 (p = 0.3582) |
 (r) (r) | Noah (male): injured (dog attack) The spike on day 2 is likely related to the hospitalisation in the afternoon before day 1. Noah was kept in the ICU. No other information is available that could explain the spike on day 7. Correlation coefficient between FCM values measured by the cortisol and 50c EIA: Pearson’s r = 0.6886 (p = 0.0699) |
| Name | GCs Last Day Administration | Second Scat Collection |
|---|---|---|
| Paddy | 13 June | 15 June |
| Noodle | 21 October | 25 October |
| Florence | 11 March | 16 March |
| Nita | 31 August | 2 September |
| Deville | 2 October | 24 November |
| Group | 50C EIA (ng/g) | Cortisol EIA (ng/g) | |
|---|---|---|---|
| Control (N = 124) | Min. Max. Mean | 1.9 85.6 24.3 | 2.2 40.1 8.4 |
| Diseased (N = 76) | Min. Max. Mean | 7.4 135.7 41.2 | 1.9 86.6 18.2 |
| Injured (N = 30) | Min. Max. Mean | 7.9 371.4 47.9 | 2.8 90.5 15.2 |
| 50c EIA (ng/g) | Cortisol EIA (ng/g) | ||||
|---|---|---|---|---|---|
| Females | Males | Females | Males | ||
| Control (Females N = 63 Males N = 61) | Min. Max. Mean | 1.9 52.4 16.8 | 4.5 85.6 32.1 | 2.2 31.4 7.6 | 2.8 40.1 9.2 |
| Diseased (Females N = 42 Males N = 34) | Min. Max. Mean | 7.4 83.5 34.8 | 8.8 135.7 49.1 | 1.9 77.1 17.7 | 2.6 86.6 19.0 |
| Injured (Females N = 11 Males N = 19) | Min. Max. Mean | 7.9 371.4 62.0 | 10.6 120.0 39.7 | 2.9 58.2 19.0 | 2.8 90.5 13.1 |
Appendix B

| 50c EIA (ng/g) (95% CI) | Cortisol EIA (ng/g) (95% CI) | |||
|---|---|---|---|---|
| Breeding (N = 90) | Non-Breeding (N = 34) | Breeding (N = 90) | Non-Breeding (N = 34) | |
| Median (95% CI) | 20.6 (18.2, 27.3) | 17.2 (13.7, 22.4) | 6.4 (5.7, 7.2) | 6.8 (5.1, 10.6) |
| Mean | 25.4 | 21.5 | 8.1 | 9.1 |
| Min/max | 1.9/85.6 | 4.5/72.6 | 2.2/40.1 | 2.2/35.5 |
| 50c EIA | IRR 95%CI (Low, High) | p-Value | Wald Test p-Value |
|---|---|---|---|
| Sex | |||
| Female | 1 | ||
| Male | 1.9 (1.5, 2.3) | <0.001 | |
| Condition | |||
| Control | 1 | <0.001 | |
| Diseased | 2.0 (1.6, 2.6) | <0.001 | |
| Injured | 3.6 (1.4, 9.6) | 0.008 | |
| Sex # Condition | |||
| Male#Control vs. Female#Control | 1 | 0.0307 | |
| Male#Diseased vs. Female#Diseased | 0.7 (0.5, 1) | 0.074 | |
| Male#Injured vs. Female#Injured | 0.3 (0.1, 0.9) | 0.033 | |
| Cortisol EIA | IRR 95%CI (low, high) | p-value | Wald test p-value |
| Sex | |||
| Female | 1 | ||
| Male | 1.2 (0.9, 1.5) | 0.135 | |
| Condition | |||
| Control | 1 | <0.001 | |
| Diseased | 2.3 (1.6, 3.1) | 0 | |
| Injured | 2.5 (1.4, 4.2) | 0.001 | |
| Sex # Condition | |||
| Male#Control vs. Female#Control | 0.4221 | ||
| Male#Diseased vs. Female#Diseased | 0.8 (0.5, 1,4) | 0.636 | |
| Male#Injured vs. Female#Injured | 0.5 (0.2, 1.3) | 0.2 |
References
- McCormick, C.M.; Mathews, I.Z.; Thomas, C.; Waters, P. Investigations of HPA function and the enduring consequences of stressors in adolescence in animal models. Brain Cogn. 2010, 72, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Björntorp, P. International Textbook of Obesity; Wiley Online Library: Hoboken, NJ, USA, 2001. [Google Scholar]
- Palme, R.; Rettenbacher, S.; Touma, C.; El-Bahr, S.; Möstl, E. Stress hormones in mammals and birds: Comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Ann. N. Y. Acad. Sci. 2005, 1040, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiga, F.; Walker, J.J.; Terry, J.R.; Lightman, S.L. HPA axis-rhythms. Compr. Physiol. 2011, 4, 1273–1298. [Google Scholar]
- Möstl, E.; Rettenbacher, S.; Palme, R. Measurement of corticosterone metabolites in birds’ droppings: An analytical approach. Ann. N. Y. Acad. Sci. 2005, 1046, 17–34. [Google Scholar] [CrossRef]
- Hannibal, K.E.; Bishop, M.D. Chronic stress, cortisol dysfunction, and pain: A psychoneuroendocrine rationale for stress management in pain rehabilitation. Phys. Ther. 2014, 94, 1816–1825. [Google Scholar] [CrossRef]
- Guilliams, T.G.; Edwards, L. Chronic stress and the HPA axis. Standard 2010, 9, 1–12. [Google Scholar]
- Coutinho, A.E.; Chapman, K.E. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
- Caldji, C.; Liu, D.; Sharma, S.; Diorio, J.; Francis, D.; Meaney, M.J.; Plotsky, P.M. Development of individual differences in behavioral and endocrine responses to stress: Role of the postnatal environment. Compr. Physiol. 2010, 271–292. [Google Scholar]
- Romero, L.M. Physiological stress in ecology: Lessons from biomedical research. Trends Ecol. Evol. 2004, 19, 249–255. [Google Scholar] [CrossRef]
- Deem, S.L.; Karesh, W.B.; Weisman, W. Putting theory into practice: Wildlife health in conservation. Conserv. Biol. 2001, 15, 1224–1233. [Google Scholar] [CrossRef]
- Department of Agriculture Water and the Environment. EPBC Act Referral Guidelines for the Vulnerable Koala. Available online: https://www.awe.gov.au/environment/biodiversity/threatened/publications/epbc-act-referral-guidelines-vulnerable-koala#:~:text=On%2012%20February%202022%20the,a%20lower%20degree%20of%20endangerment (accessed on 8 August 2022).
- Gonzalez-Astudillo, V.; Allavena, R.; McKinnon, A.; Larkin, R.; Henning, J. Decline causes of Koalas in South East Queensland, Australia: A 17-year retrospective study of mortality and morbidity. Sci. Rep. 2017, 7, 42587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henning, J.; Hannon, C.; McKinnon, A.; Larkin, R.; Allavena, R. The causes and prognoses of different types of fractures in wild koalas submitted to wildlife hospitals. Prev. Vet. Med. 2015, 122, 371–378. [Google Scholar] [CrossRef] [PubMed]
- McAlpine, C. Relationships between human-induced habitat disturbance, stressors and disease in Koalas. In Proceedings of the Koala Research Network Disease Workshop, Brisbane, Australia, 9 June 2011. [Google Scholar]
- Schlagloth, R.; Santamaria, F.; Melzer, A.; Keatley, M.R.; Houston, W. Vehicle collisions and dog attacks on Victorian koalas as evidenced by a retrospective analysis of sightings and admission records 1997–2011. Aust. Zool. 2021, 42, 655–666. [Google Scholar] [CrossRef]
- Department of Environment and Heritage Protection. Koala Threats. Available online: https://environment.des.qld.gov.au/ (accessed on 27 August 2021).
- Department of Environment and Heritage Protection. Living with Wildlife-Koalas and Cars. Available online: https://environment.des.qld.gov.au/__data/assets/pdf_file/0025/86416/koalas-and-cars.pdf (accessed on 27 August 2021).
- Davies, N.; Gillett, A.; McAlpine, C.; Seabrook, L.; Baxter, G.; Lunney, D.; Bradley, A. The effect of ACTH upon faecal glucocorticoid excretion in the koala. J. Endocrinol. 2013, 219, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burach, F.; Pospischil, A.; Hanger, J.; Loader, J.; Pillonel, T.; Greub, G.; Borel, N. Chlamydiaceae and Chlamydia-like organisms in the koala (Phascolarctos cinereus)–organ distribution and histopathological findings. Vet. Microbiol. 2014, 172, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, F.; Schlagloth, R. The effect of Chlamydia on translocated Chlamydia-naïve koalas; a case study. Aust. Zool. 2016, 38, 192–202. [Google Scholar] [CrossRef]
- Scheun, J.; Ludynia, K.; Snyman, A.; Ganswindt, A. Non-invasive hormone monitoring as a robust method for determining adrenocortical activity in injured, emaciated and oil-contaminated African penguins undergoing rehabilitation. Gen. Comp. Endocrinol. 2021, 303, 113703. [Google Scholar] [CrossRef]
- Rolland, R.M.; McLellan, W.A.; Moore, M.J.; Harms, C.A.; Burgess, E.A.; Hunt, K.E. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 2017, 34, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Wolf, T.E.; Valades, G.B.; Simelane, P.; Bennett, N.C.; Ganswindt, A. The relationship between physical injury, body condition and stress-related hormone concentrations in free-ranging giraffes. Wildl. Biol. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Merl, S.; Scherzer, S.; Palme, R.; Möstl, E. Pain causes increased concentrations of glucocorticoid metabolites in horse feces. J. Equine Vet. Sci. 2000, 20, 586–590. [Google Scholar] [CrossRef]
- Palme, R.; Möstl, E. Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Int. J. Mamm. Biol. 1997, 62, 192–197. [Google Scholar]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Narayan, E. Physiological stress levels in wild koala sub-populations facing anthropogenic induced environmental trauma and disease. Sci. Rep. 2019, 9, 6031. [Google Scholar] [CrossRef] [PubMed]
- Ganswindt, A.; Münscher, S.; Henley, M.; Palme, R.; Thompson, P.; Bertschinger, H. Concentrations of faecal glucocorticoid metabolites in physically injured free-ranging African elephants Loxodonta africana. Wildl. Biol. 2010, 16, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossey, B.; Chimimba, C.; Du Plessis, C.; Hall, G.; Ganswindt, A. Using faecal glucocorticoid metabolite analyses to elucidate stressors of African wild dogs Lycaon pictus from South Africa. Wildl. Biol. 2020, 2020, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, F.; Barlow, C.K.; Schlagloth, R.; Schittenhelm, R.; Palme, R.; Henning, J. Identification of koala (Phascolarctos cinereus) faecal cortisol metabolites using liquid chromatography-mass spectrometry and enzyme immunoassays. Metabolites 2021, 11, 393. [Google Scholar] [CrossRef]
- Santamaria, F.; Palme, R.; Schlagloth, R.; Klobetz-Rassam, E.; Henning, J. Seasonal variations of faecal cortisol metabolites in koalas in South East Queensland. Animals 2021, 11, 1622. [Google Scholar] [CrossRef]
- Santamaria, F.; Schlagloth, R.; Palme, R.; Henning, J. Over time decay of cortisol metabolites in faecal pellets of koalas in Central Queensland. Animals 2021, 11, 3376. [Google Scholar] [CrossRef]
- McCallum, H.; Kerlin, D.H.; Ellis, W.; Carrick, F. Assessing the significance of endemic disease in conservation–koalas, chlamydia and koala retrovirus as a case study. Conserv. Lett. 2018, 11, e12425. [Google Scholar] [CrossRef]
- Hulse, L.S.; McDonald, S.; Johnston, S.D.; Beagley, K.W. Rapid point-of-care diagnostics for the detection of Chlamydia pecorum in koalas (Phascolarctos cinereus) using loop-mediated isothermal amplification without nucleic acid purification. MicrobiologyOpen 2019, 8, e916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quillfeldt, P.; Möstl, E. Resource allocation in Wilson’s storm-petrels Oceanites oceanicus determined by measurement of glucocorticoid excretion. Acta Ethol. 2003, 5, 115–122. [Google Scholar] [CrossRef]
- Mood, A.; Graybill, F. Introduction to the Theory of Statistics, 2nd ed.; McGraw-Hill: New York, NY, USA, 1963. [Google Scholar]
- Dinno, A. Nonparametric pairwise multiple comparisons in independent groups using Dunn’s test. Stata J. 2015, 15, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.C.; Trivedi, P.K. Microeconometrics Using Stata; Stata Press: College Station, TX, USA, 2010; Volume 2. [Google Scholar]
- Cope, H.R.; Keeley, T.; Keong, J.; Smith, D.; Silva, F.R.; McArthur, C.; Webster, K.N.; Mella, V.S.; Herbert, C.A. Validation of an enzyme immunoassay to measure faecal glucocorticoid metabolites in common brushtail possums (Trichosurus vulpecula) to evaluate responses to rehabilitation. Animals 2022, 12, 1627. [Google Scholar] [CrossRef] [PubMed]
- McCallum, H.; Dobson, A. Disease, habitat fragmentation and conservation. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 2041–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, N.; Gramotnev, G.; McAlpine, C.; Seabrook, L.; Baxter, G.; Lunney, D.; Rhodes, J.R.; Bradley, A. Physiological stress in koala populations near the arid edge of their distribution. PLoS ONE 2013, 8, e79136. [Google Scholar] [CrossRef]
- Acsai, A.; Drexler, T.; Evans, N.; McCafferty, D. Low levels of faecal cortisol in bank voles (Myodes glareolus) in response to live-trapping. Glasg. Nat. 2021, 27, 19–27. [Google Scholar] [CrossRef]
- Champagne, C.D.; Kellar, N.M.; Crocker, D.E.; Wasser, S.K.; Booth, R.K.; Trego, M.L.; Houser, D.S. Blubber cortisol qualitatively reflects circulating cortisol concentrations in bottlenose dolphins. Mar. Mammal Sci. 2017, 33, 134–153. [Google Scholar] [CrossRef]
- Fanson, K.V.; Best, E.C.; Bunce, A.; Fanson, B.G.; Hogan, L.A.; Keeley, T.; Narayan, E.J.; Palme, R.; Parrott, M.L.; Sharp, T.M. One size does not fit all: Monitoring faecal glucocorticoid metabolites in marsupials. Gen. Comp. Endocrinol. 2015, 244, 146–156. [Google Scholar] [CrossRef]
- Pahuja, H.K.; Narayan, E.J. Comparing the agreement of a commercial cortisol kit with a biologically validated assay in evaluating faecal cortisol metabolite levels in koala joeys. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2022, 276, 111353. [Google Scholar] [CrossRef]
- Anderwald, P.; Campell Andri, S.; Palme, R. Reflections of ecological differences? Stress responses of sympatric Alpine chamois and red deer to weather, forage quality, and human disturbance. Ecol. Evol. 2021, 11, 15740–15753. [Google Scholar] [CrossRef] [PubMed]
- Dupoué, A.; Rutschmann, A.; Le Galliard, J.F.; Clobert, J.; Blaimont, P.; Sinervo, B.; Miles, D.B.; Haussy, C.; Meylan, S. Reduction in baseline corticosterone secretion correlates with climate warming and drying across wild lizard populations. J. Anim. Ecol. 2018, 87, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Brown, J.; Swedell, L. Anthropogenic effects on the physiology and behaviour of chacma baboons in the Cape Peninsula of South Africa. Conserv. Physiol. 2020, 8, coaa066. [Google Scholar] [CrossRef] [PubMed]
- Young, K.; Walker, S.; Lanthier, C.; Waddell, W.; Monfort, S.L.; Brown, J.L. Noninvasive monitoring of adrenocortical activity in carnivores by fecal glucocorticoid analyses. Gen. Comp. Endocrinol. 2004, 137, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Salaberger, T.; Millard, M.; El Makarem, S.; Möstl, E.; Grünberger, V.; Krametter-Frötscher, R.; Wittek, T.; Palme, R. Influence of external factors on hair cortisol concentrations. Gen. Comp. Endocrinol. 2016, 233, 73–78. [Google Scholar] [CrossRef]
- Yadav, V.; Gaisford, S.; Merchant, H.A.; Basit, A.W. Colonic bacterial metabolism of corticosteroids. Int. J. Pharm. 2013, 457, 268–274. [Google Scholar] [CrossRef]
- Watson, R.; Munro, C.; Edwards, K.L.; Norton, V.; Brown, J.L.; Walker, S.L. Development of a versatile enzyme immunoassay for non-invasive assessment of glucocorticoid metabolites in a diversity of taxonomic species. Gen. Comp. Endocrinol. 2013, 186, 16–24. [Google Scholar] [CrossRef]
- Dehnhard, M.; Clauss, M.; Lechner-Doll, M.; Meyer, H.; Palme, R. Noninvasive monitoring of adrenocortical activity in roe deer (Capreolus capreolus) by measurement of fecal cortisol metabolites. Gen. Comp. Endocrinol. 2001, 123, 111–120. [Google Scholar] [CrossRef]
- Vera, F.; Antenucci, C.D.; Zenuto, R.R. Different regulation of cortisol and corticosterone in the subterranean rodent Ctenomys talarum: Responses to dexamethasone, angiotensin II, potassium, and diet. Gen. Comp. Endocrinol. 2019, 273, 108–117. [Google Scholar] [CrossRef]
- Lepschy, M.; Touma, C.; Hruby, R.; Palme, R. Non-invasive measurement of adrenocortical activity in male and female rats. Lab. Anim. 2007, 41, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Palme, R.; Robia, C.; Messmann, S.; Hofer, J.; Mostl, E. Measurement of faecal cortisol metabolites in ruminants: A non-invasive parameter of adrenocortical function. Wien. Tierärztliche Mon. 1999, 86, 237–241. [Google Scholar]
- Dehnhard, M.; Schreer, A.; Krone, O.; Jewgenow, K.; Krause, M.; Grossmann, R. Measurement of plasma corticosterone and fecal glucocorticoid metabolites in the chicken (Gallus domesticus), the great cormorant (Phalacrocorax carbo), and the goshawk (Accipiter gentilis). Gen. Comp. Endocrinol. 2003, 131, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Bamford, N.; Harris, P.; Bailey, S. Circannual variation in plasma adrenocorticotropic hormone concentrations and dexamethasone suppression test results in Standardbred horses, Andalusian horses and mixed-breed ponies. Aust. Vet. J. 2020, 98, 616–621. [Google Scholar] [CrossRef]
- Borer-Weir, K.; Menzies-Gow, N.; Bailey, S.; Harris, P.; Elliott, J. Seasonal and annual influence on insulin and cortisol results from overnight dexamethasone suppression tests in normal ponies and ponies predisposed to laminitis. Equine Vet. J. 2013, 45, 688–693. [Google Scholar] [CrossRef]
- Dickens, M.J.; Delehanty, D.J.; Romero, L.M. Stress and translocation: Alterations in the stress physiology of translocated birds. Proc. Biol. Sci. R. Soc. 2009, 276, 2051–2056. [Google Scholar] [CrossRef]
- Ceulemans, D.L.; Westenberg, H.G.; Van Praag, H.M. The effect of stress on the dexamethasone suppression test. Psychiatry Res. 1985, 14, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Padgett, D.A.; Glaser, R. How stress influences the immune response. Trends Immunol. 2003, 24, 444–448. [Google Scholar] [CrossRef]
- McLaren, G.; Bonacic, C.; Rowan, A. Animal Welfare and Conservation: Measuring Stress in the Wild; Blackwell Publishing: Melbourne, VIC, Australia, 2007. [Google Scholar]
- Hofer, H.; East, M.L. Biological conservation and stress. Adv. Study Behav. 1998, 27, 405–525. [Google Scholar]
- Mesotten, D.; Vanhorebeek, I.; Van den Berghe, G. The altered adrenal axis and treatment with glucocorticoids during critical illness. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 496–505. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Kumar, V.; Chandrasekhar, M.; Malviya, M.; Ganswindt, A.; Ramesh, K.; Sankar, K.; Umapathy, G. Glucocorticoid stress responses of reintroduced tigers in relation to anthropogenic disturbance in Sariska Tiger Reserve in India. PLoS ONE 2015, 10, e0127626. [Google Scholar] [CrossRef] [Green Version]
- Touma, C.; Sachser, N.; Möstl, E.; Palme, R. Effect of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen. Comp. Endocrinol. 2003, 130, 267–278. [Google Scholar] [CrossRef] [PubMed]







| AZ | ML | |||
|---|---|---|---|---|
| Diseased 1 | Injured 2 | Diseased + Injured | Control 3 | External control 4 |
| 76 (42F + 34M) | 30 (11F + 19M) | 4 (2F + 2M) | 36 (19F + 17M) | 88 (44F + 44M) |
| Non-breeding season N = 74 Breeding season N = 72 | Non-breeding season N = 76; Breeding season N = 12 | |||
| Condition | N | Sampling Point | Mean Days between Samplings | Median 50c EIA (ng/g) (95% CI) | 50c EIA p-Value * | Median Cortisol EIA (ng/g) (95% CI) | Cortisol EIA p-Value * |
|---|---|---|---|---|---|---|---|
| Control | 7 | 1 | 19.7 (13.2, 32.3) | 9.7 (3.1, 25.4) | |||
| 2 | 12.2 | 12.4 (8.8, 30.2) | 0.125 | 3.3 (2.2, 11.6) | 0.4531 | ||
| Diseased | 32 | 1 | 33.5 (26.0, 50.2) | 9.8 (6.3, 14.3) | |||
| 2 | 13.2 | 36.2 (25.0, 52.3) | 0.4869 | 9.3 (5.7, 19.4) | 0.8601 | ||
| Injured | 13 | 1 | 34.2 (26.2, 38.8) | 9.8 (5.2, 15.5) | |||
| 2 | 12.9 | 30.0 (20.9, 42.2) | 0.5811 | 8.8 (4.5, 11.0) | 0.0225 |
| 50c EIA (ng/g) (95% CI) | Cortisol EIA (ng/g) (95% CI) | |||
|---|---|---|---|---|
| Females | Males | Females | Males | |
| Control (Females N = 63 Males N = 61) | 14.4 (12.1, 17.2) | 29.4 (23.3, 36.5) | 6.4 (5.5, 7.2) | 6.8 (6.0, 8.1) |
| Diseased (Females N = 42 Males N = 34) | 29.2 (20.6, 43.1) | 44.7 (33.4, 57.4) | 11.4 (9.3, 17.8) | 11.0 (8.9, 16.2) |
| Injured (Females N = 11 Males N = 19) | 29.5 (16.6, 52.6) | 35.7 (32.7, 40.7) | 11.8 (6.0, 33.6) | 6.6 (5.1, 12.5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamaria, F.; Schlagloth, R.; Valenza, L.; Palme, R.; de Villiers, D.; Henning, J. The Effect of Disease and Injury on Faecal Cortisol Metabolites, as an Indicator of Stress in Wild Hospitalised Koalas, Endangered Australian Marsupials. Vet. Sci. 2023, 10, 65. https://doi.org/10.3390/vetsci10010065
Santamaria F, Schlagloth R, Valenza L, Palme R, de Villiers D, Henning J. The Effect of Disease and Injury on Faecal Cortisol Metabolites, as an Indicator of Stress in Wild Hospitalised Koalas, Endangered Australian Marsupials. Veterinary Sciences. 2023; 10(1):65. https://doi.org/10.3390/vetsci10010065
Chicago/Turabian StyleSantamaria, Flavia, Rolf Schlagloth, Ludovica Valenza, Rupert Palme, Deidre de Villiers, and Joerg Henning. 2023. "The Effect of Disease and Injury on Faecal Cortisol Metabolites, as an Indicator of Stress in Wild Hospitalised Koalas, Endangered Australian Marsupials" Veterinary Sciences 10, no. 1: 65. https://doi.org/10.3390/vetsci10010065
APA StyleSantamaria, F., Schlagloth, R., Valenza, L., Palme, R., de Villiers, D., & Henning, J. (2023). The Effect of Disease and Injury on Faecal Cortisol Metabolites, as an Indicator of Stress in Wild Hospitalised Koalas, Endangered Australian Marsupials. Veterinary Sciences, 10(1), 65. https://doi.org/10.3390/vetsci10010065