


Impact of Selected Bacterial and Viral Toll-like Receptor Agonists on the Phenotype and Function of Camel Blood Neutrophils
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Animals, Materials, and Methods
2.1. Animals and Sampling
2.2. Isolation of Neutrophils from Camel Blood
2.3. Reagents
2.4. In Vitro Stimulation of Purified Neutrophils with Synthetic Toll-like Receptor Ligands
2.5. Stimulation-Induced Neutrophil Cell Size Change
2.6. Impact of TLR Priming on Bacterial Phagocytosis by Camel Neutrophils
2.7. Generation of Reactive Oxygen Species by Neutrophils
2.8. Cell Surface Antigen Expression on Neutrophils
2.9. Neutrophils Vitality Assay
2.10. Statistical Analysis
3. Results
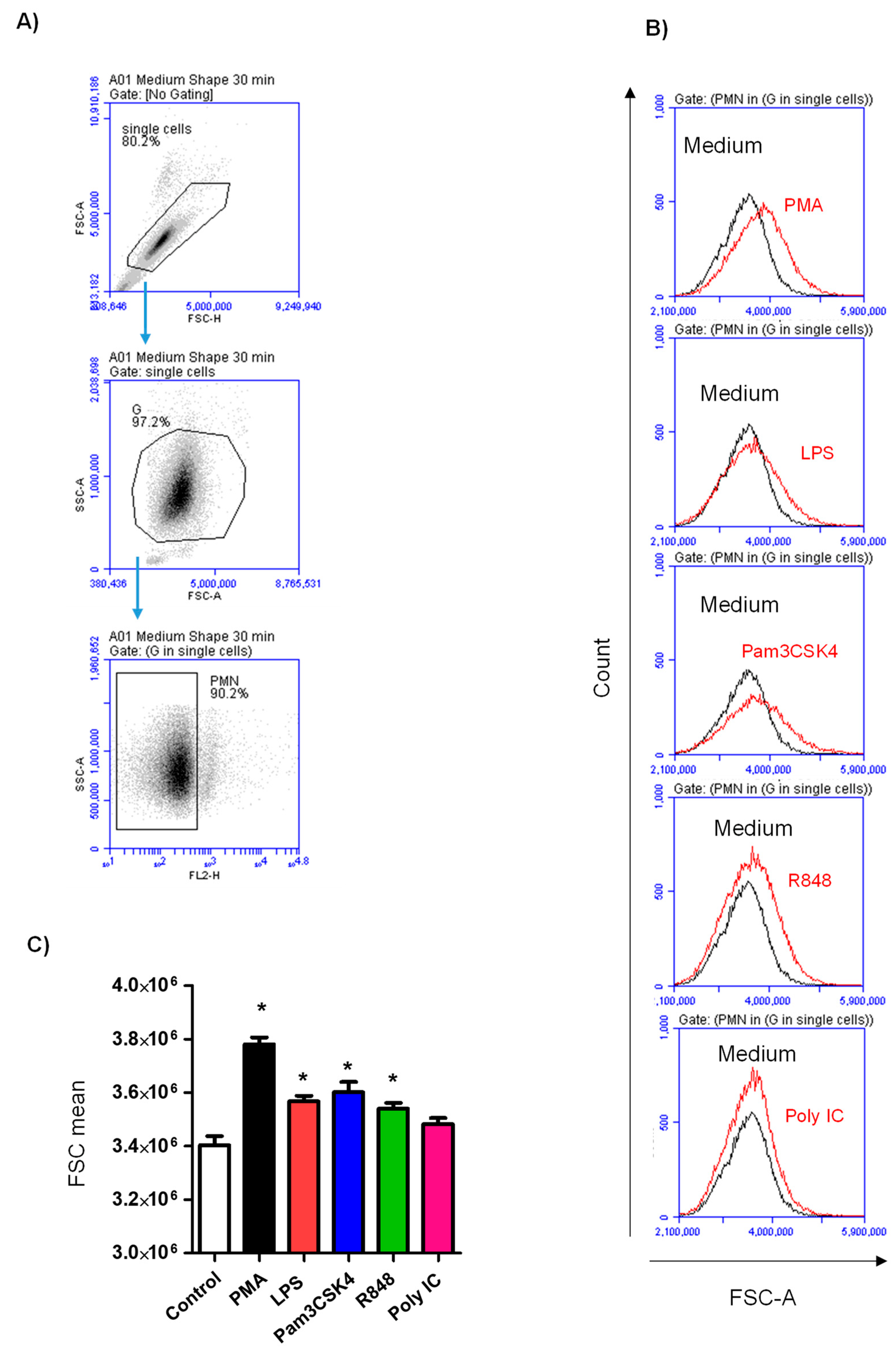
3.1. Impact of Toll-like Receptor Ligands on Cell Size of Stimulated Neutrophils
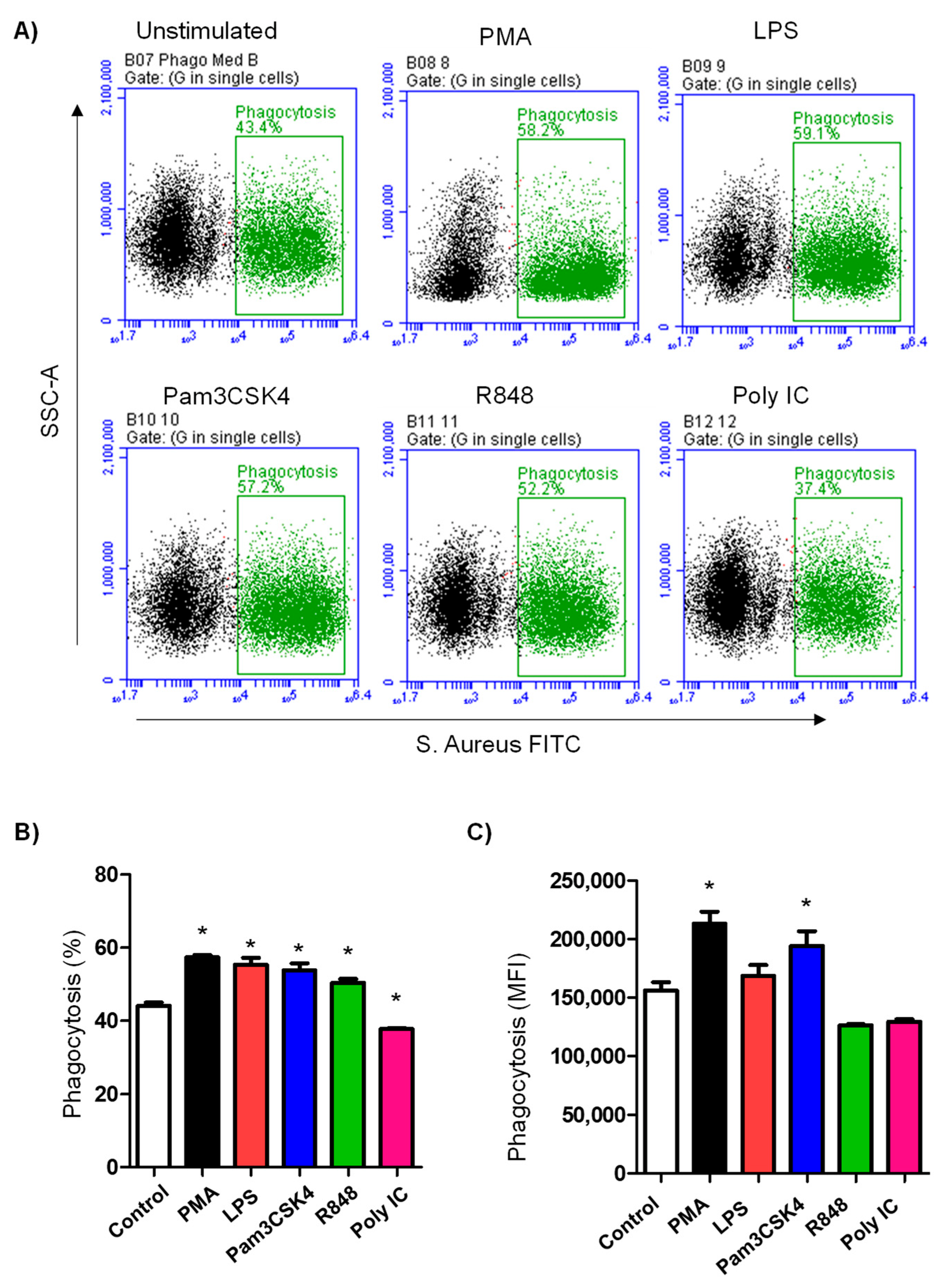
3.2. TLR Ligands Differently Modulate the Phagocytosis Activity of Camel Neutrophils
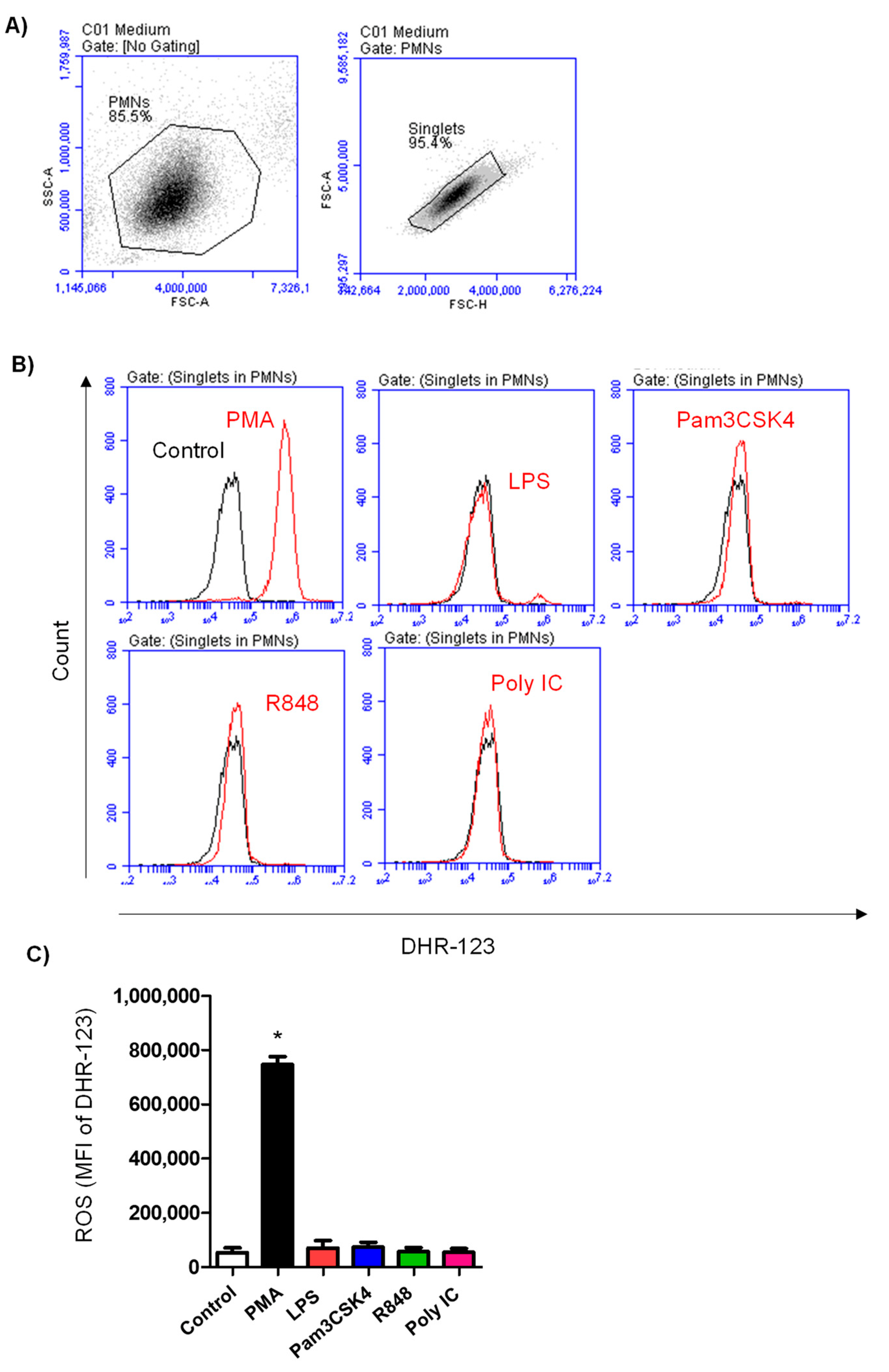
3.3. ROS Production in Neutrophils after TLR Stimulation
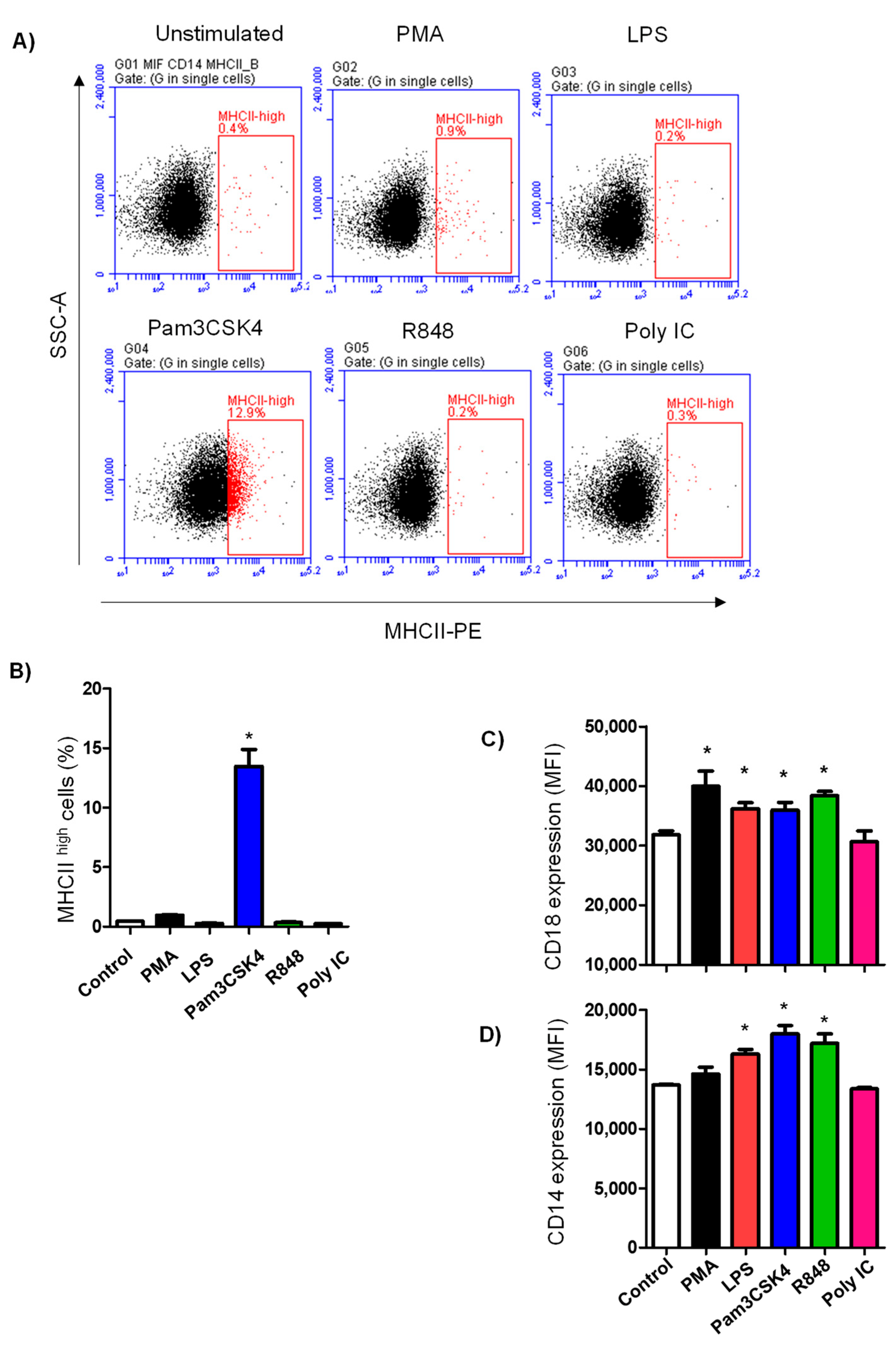
3.4. TLR Ligands Modulate the Expression of Some Neutrophil Cell-Surface Antigens
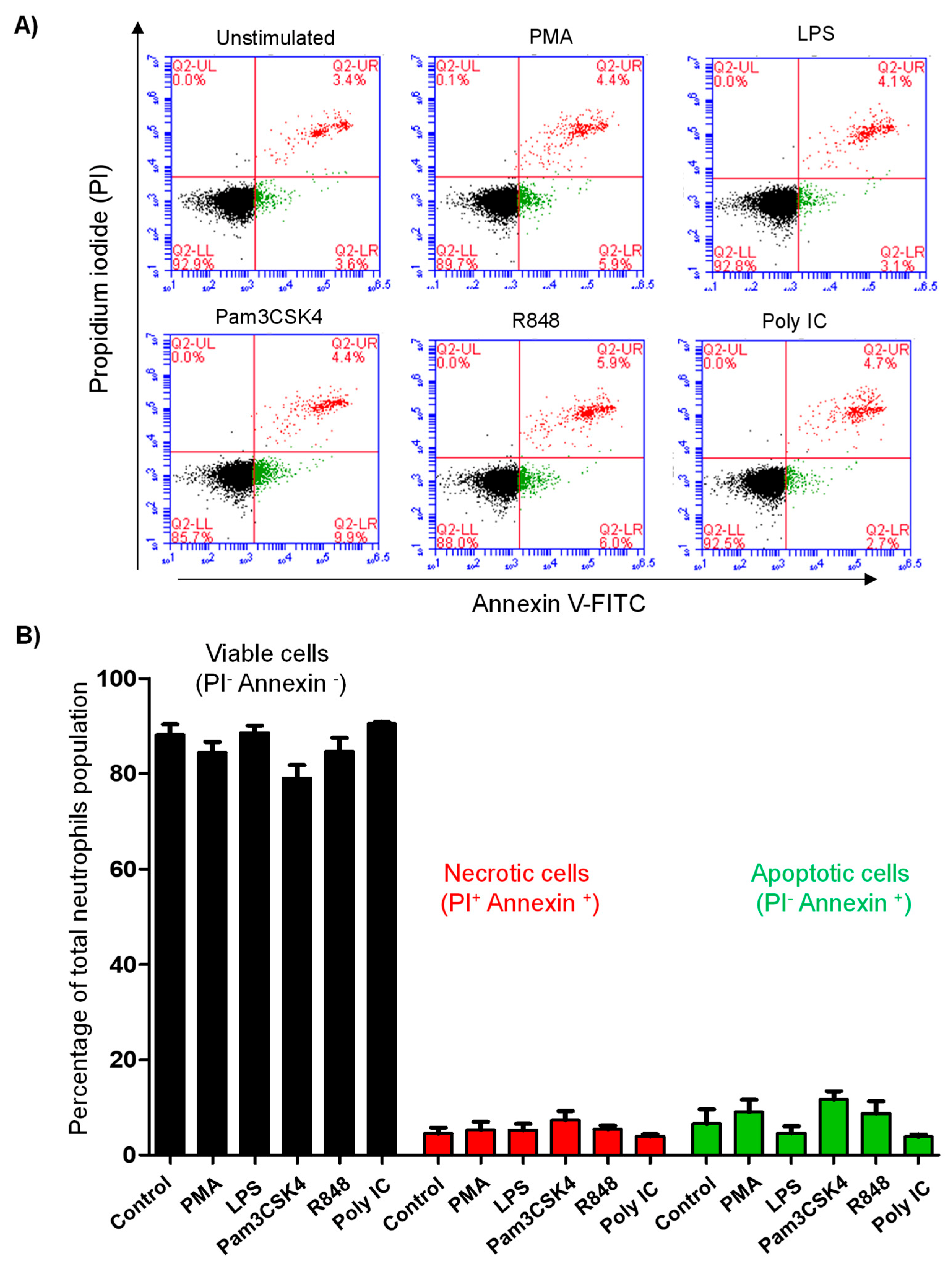
3.5. Stimulation with TLR Ligands Does Not Affect Cell Vitality of Camel Neutrophils
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soehnlein, O.; Lindbom, L. Phagocyte partnership during the onset and resolution of inflammation. Nat. Rev. Immunol. 2010, 10, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C.; Demaurex, N.; Lowell, C.A.; Uribe-Querol, E. Neutrophils: Their Role in Innate and Adaptive Immunity. J. Immunol. Res. 2016, 2016, 1469780. [Google Scholar] [CrossRef]
- Tan, X.; Sun, L.; Chen, J.; Chen, Z.J. Detection of Microbial Infections Through Innate Immune Sensing of Nucleic Acids. Annu. Rev. Microbiol. 2018, 72, 447–478. [Google Scholar] [CrossRef]
- Akira, S. Pathogen recognition by innate immunity and its signaling. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 143–156. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Pathogen recognition or homeostasis? APC receptor functions in innate immunity. Comptes Rendus Biol. 2004, 327, 603–607. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Gordon, S. The role of scavenger receptors in pathogen recognition and innate immunity. Immunobiology 2004, 209, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pathogen recognition by innate immunity. Arerugi 2007, 56, 558–562. [Google Scholar]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between toll-like receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 13766–13771. [Google Scholar] [CrossRef]
- Beutler, B. Inferences, questions and possibilities in Toll-like receptor signalling. Nature 2004, 430, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.; Krook, H.; Goto, M.; Korsgren, O. MyD88-dependent toll-like receptor signalling is not a requirement for fetal islet xenograft rejection in mice. Xenotransplantation 2004, 11, 347–352. [Google Scholar] [CrossRef]
- Radoshevich, L.; Dussurget, O. Cytosolic Innate Immune Sensing and Signaling upon Infection. Front. Microbiol. 2016, 7, 313. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, H.; Kudo, K.; Mori, K.; Nagai, F.; Hatsugaya, A.; Tajima, M.; Tamura, K.; Hoshi, F.; Koiwa, M.; Kawamura, S. Acute phase response in naturally occurring coliform mastitis. J. Vet. Med. Sci. 2001, 63, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Johnzon, C.F.; Dahlberg, J.; Gustafson, A.M.; Waern, I.; Moazzami, A.A.; Ostensson, K.; Pejler, G. The Effect of Lipopolysaccharide-Induced Experimental Bovine Mastitis on Clinical Parameters, Inflammatory Markers, and the Metabolome: A Kinetic Approach. Front. Immunol. 2018, 9, 1487. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K. Innate recognition of lipopolysaccharide by Toll-like receptor 4-MD-2. Trends Microbiol. 2004, 12, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K. Innate recognition of lipopolysaccharide by CD14 and toll-like receptor 4-MD-2: Unique roles for MD-2. Int. Immunopharmacol. 2003, 3, 119–128. [Google Scholar] [CrossRef]
- Miyake, K.; Ogata, H.; Nagai, Y.; Akashi, S.; Kimoto, M. Innate recognition of lipopolysaccharide by Toll-like receptor 4/MD-2 and RP105/MD-1. J. Endotoxin Res. 2000, 6, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Mintz, M.; Mintz, D.; Ezra-Elia, R.; Shpigel, N.Y. Pam3CSK4/TLR2 signaling elicits neutrophil recruitment and restricts invasion of Escherichia coli P4 into mammary gland epithelial cells in a murine mastitis model. Vet. Immunol. Immunopathol. 2013, 152, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.; Beynon, C.; Kennedy, E.; O’Farrelly, C.; Meade, K.G. Bovine innate immune phenotyping via a standardized whole blood stimulation assay. Sci. Rep. 2021, 11, 17227. [Google Scholar] [CrossRef]
- Prince, L.R.; Whyte, M.K.; Sabroe, I.; Parker, L.C. The role of TLRs in neutrophil activation. Curr. Opin. Pharm. 2011, 11, 397–403. [Google Scholar] [CrossRef]
- Hussen, J.; Koy, M.; Petzl, W.; Schuberth, H.J. Neutrophil degranulation differentially modulates phenotype and function of bovine monocyte subsets. Innate Immun. 2016, 22, 124–137. [Google Scholar] [CrossRef]
- Conejeros, I.; Gibson, A.J.; Werling, D.; Munoz-Caro, T.; Hermosilla, C.; Taubert, A.; Burgos, R.A. Effect of the synthetic Toll-like receptor ligands LPS, Pam3CSK4, HKLM and FSL-1 in the function of bovine polymorphonuclear neutrophils. Dev. Comp. Immunol. 2015, 52, 215–225. [Google Scholar] [CrossRef]
- Silva, V.M.; Souza, M.T.; Blagitz, M.G.; Souza, F.N.; Batista, C.F.; Alves, A.J.; Fernandes, A.C.C.; Sanchez, E.M.R.; Ordinola-Ramirez, C.M.; da Costa, L.; et al. Milk lymphocyte profile and macrophage functions: New insights into the immunity of the mammary gland in quarters infected with Corynebacterium bovis. BMC Vet. Res. 2021, 17, 282. [Google Scholar] [CrossRef]
- Mosaad, A.A.; Elbagory, A.R.; Khalid, A.M.; Waters, W.; Tibary, A.; Hamilton, M.J.; Davis, W.C. Identification of monoclonal antibody reagents for use in the study of the immune response to infectious agents in camel and water buffalo. J. Camel Pract. Res. 2006, 13, 91–101. [Google Scholar]
- Al-Mubarak, A.I.A. Differential expression of the coronavirus (Mers-cov) Receptor, Dipeptidyl Peptidase 4, on normal and stimulated leukocytes of dromedary camels. J. Camel. Pract. Res. 2018, 25, 249. [Google Scholar] [CrossRef]
- Hussen, J.; Shawaf, T.; Al-herz, A.I.; Alturaifi, H.R.; Al khamees, M.; Alluwaimi, A.M. Expression Patterns of Cell Adhesion Molecules on CD4+ T Cells and WC1+ T Cells in the Peripheral Blood of Dromedary Camels. Pak. Vet. J. 2018, 38, 231–236. [Google Scholar] [CrossRef]
- Hussen, J.; Shawaf, T.; Al-herz, A.I.; Alturaifi, H.R.; Alluwaimi, A.M. Reactivity of commercially available monoclonal antibodies to human CD antigens with peripheral blood leucocytes of dromedary camels (Camelus dromedarius. Open Vet. J. 2017, 7, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Hussen, J.; Shawaf, T.; Al-Mubarak, A.I.A.; Al Humam, N.A.; Almathen, F.; Schuberth, H.J. Dromedary camel CD14(high) MHCII(high) monocytes display inflammatory properties and are reduced in newborn camel calves. BMC Vet. Res. 2020, 16, 62. [Google Scholar] [CrossRef]
- Burn, G.L.; Foti, A.; Marsman, G.; Patel, D.F.; Zychlinsky, A. The Neutrophil. Immunity 2021, 54, 1377–1391. [Google Scholar] [CrossRef]
- Malech, H.L.; DeLeo, F.R.; Quinn, M.T. The Role of Neutrophils in the Immune System: An Overview. Methods Mol. Biol. 2020, 2087, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. 2020, 15, 493–518. [Google Scholar] [CrossRef]
- Vijay, K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int. Immunopharmacol. 2018, 59, 391–412. [Google Scholar] [CrossRef]
- Parker, L.C.; Whyte, M.K.; Dower, S.K.; Sabroe, I. The expression and roles of Toll-like receptors in the biology of the human neutrophil. J. Leukoc. Biol. 2005, 77, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.J.; Hollers, J.C.; Crockett-Torabi, E.; Smith, C.W. Recruitment of CD11b/CD18 to the neutrophil surface and adherence-dependent cell locomotion. J. Clin. Investig. 1992, 90, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Means, T.K.; Luster, A.D. Toll-like receptors stimulate human neutrophil function. Blood 2003, 102, 2660–2669. [Google Scholar] [CrossRef] [PubMed]
- Aomatsu, K.; Kato, T.; Fujita, H.; Hato, F.; Oshitani, N.; Kamata, N.; Tamura, T.; Arakawa, T.; Kitagawa, S. Toll-like receptor agonists stimulate human neutrophil migration via activation of mitogen-activated protein kinases. Immunology 2008, 123, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Conrath, K.E.; Wernery, U.; Muyldermans, S.; Nguyen, V.K. Emergence and evolution of functional heavy-chain antibodies in Camelidae. Dev. Comp. Immunol. 2003, 27, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Mol, S.; Hafkamp, F.M.J.; Varela, L.; Simkhada, N.; Taanman-Kueter, E.W.; Tas, S.W.; Wauben, M.H.M.; Groot Kormelink, T.; de Jong, E.C. Efficient Neutrophil Activation Requires Two Simultaneous Activating Stimuli. Int. J. Mol. Sci. 2021, 22, 10106. [Google Scholar] [CrossRef] [PubMed]
- Vono, M.; Lin, A.; Norrby-Teglund, A.; Koup, R.A.; Liang, F.; Lore, K. Neutrophils acquire the capacity for antigen presentation to memory CD4(+) T cells in vitro and ex vivo. Blood 2017, 129, 1991–2001. [Google Scholar] [CrossRef] [PubMed]
- Culshaw, S.; Millington, O.R.; Brewer, J.M.; McInnes, I.B. Murine neutrophils present Class II restricted antigen. Immunol. Lett. 2008, 118, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Rambault, M.; Borkute, R.; Doz-Deblauwe, E.; Le-Vern, Y.; Winter, N.; Dorhoi, A.; Remot, A. Isolation of Bovine Neutrophils by Fluorescence- and Magnetic-Activated Cell Sorting. Methods Mol. Biol. 2021, 2236, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Rambault, M.; Gilbert, F.B.; Roussel, P.; Tessier, A.; David, V.; Germon, P.; Winter, N.; Remot, A. MHC-II expressing neutrophils circulate in blood and milk during mastitis and show high microbicidal activity. bioRxiv 2022. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussen, J.; Alkuwayti, M.A.; Falemban, B.; Alhojaily, S.M.; Adwani, S.A.; Hassan, E.A.E.; Al-Mubarak, A.I. Impact of Selected Bacterial and Viral Toll-like Receptor Agonists on the Phenotype and Function of Camel Blood Neutrophils. Vet. Sci. 2023, 10, 154. https://doi.org/10.3390/vetsci10020154
Hussen J, Alkuwayti MA, Falemban B, Alhojaily SM, Adwani SA, Hassan EAE, Al-Mubarak AI. Impact of Selected Bacterial and Viral Toll-like Receptor Agonists on the Phenotype and Function of Camel Blood Neutrophils. Veterinary Sciences. 2023; 10(2):154. https://doi.org/10.3390/vetsci10020154
Chicago/Turabian StyleHussen, Jamal, Mayyadah Abdullah Alkuwayti, Baraa Falemban, Sameer M. Alhojaily, Salma Al Adwani, El Awad El Hassan, and Abdullah IA Al-Mubarak. 2023. "Impact of Selected Bacterial and Viral Toll-like Receptor Agonists on the Phenotype and Function of Camel Blood Neutrophils" Veterinary Sciences 10, no. 2: 154. https://doi.org/10.3390/vetsci10020154
APA StyleHussen, J., Alkuwayti, M. A., Falemban, B., Alhojaily, S. M., Adwani, S. A., Hassan, E. A. E., & Al-Mubarak, A. I. (2023). Impact of Selected Bacterial and Viral Toll-like Receptor Agonists on the Phenotype and Function of Camel Blood Neutrophils. Veterinary Sciences, 10(2), 154. https://doi.org/10.3390/vetsci10020154