1. Introduction
Janus kinase (JAK) enzymes are involved in intracellular signaling pathways activated by cytokines dysregulated in various conditions ranging from cancer to a variety of inflammatory diseases such as allergies [
1]. The JAK pathway has been identified in recent years as an important target for the treatment of both inflammatory and autoimmune diseases in human medicine. In veterinary medicine, the main experience with JAK inhibitors has been with oclacitinib, which received FDA approval for the management of canine allergic skin diseases in 2013. Oclacitinib has been very successful in providing fast relief from pruritus and a high level of efficacy to decrease the severity of signs associated with canine atopic dermatitis [
2]. Oclacitinib is considered a selective JAK 1 inhibitor at labeled doses, thus primarily interfering with the signaling of allergic cytokines and minimally interfering with hematopoiesis and natural immunity. This is an important distinction between oclacitinib and many JAK inhibitors approved for use in human medicine that target other JAK pathways. Interestingly, despite the extensive use of oclacitinib in the past decade, no study has reported on the expression of JAKs in canine skin. Currently, the localization and expression of JAK1 and JAK3 in canine skin have never been described, and the effect of allergen exposure on the expression of JAKs in canine skin is unknown.
In normal human epidermis, JAK expression and distribution have been characterized by Nishio et al. [
3]. In that study, a strong immunoreactivity for JAK1 and JAK3 was described in the skin, and differential expression of various JAKS was reported in the epidermis, highlighting a likely role for JAKS in keratinocyte differentiation. A few years later, immunohistochemistry on healthy and lesional human skin was performed by Juczynska et al., showing the same staining patterns [
4]. JAK1 was reported to be cytoplasmic, and JAK3 was found to be perinuclear and nuclear in both studies [
3,
4]. JAK 1 was found throughout the epidermis, and the horny cell layer was not stained [
3]. The expression of JAK3 was found throughout the epidermis with the horny and granular layers being strongly stained by the antibody [
4].
The expression of the JAK-STAT pathway in the epidermis suggests its role in keratinocyte differentiation, and this was nicely demonstrated in a Japanese study in 2015 [
5]. In that study, the role of JAK-STAT signaling in skin barrier function was evaluated from the perspective of atopic dermatitis. Microarray analysis with a human skin equivalent model revealed that IL-4/IL-13 dominantly affected keratinocyte differentiation, presumably through JAK-STAT signaling. It was also found that the JAK inhibitor JTE-052 promoted filaggrin and loricrin protein production and that STAT3 is the key transcriptional factor in keratinocyte differentiation. These findings supported the notion that JAK-STAT3 signaling plays a key role in modulating keratinocyte differentiation [
5]. There is also evidence that the JAK/STAT pathway is essential to the normal functioning of the immune system [
6]. Stimulation of the JAK/STAT pathway facilitates intercellular communication and plays a significant role in cell processes such as proliferation, growth, differentiation, migration, and apoptosis [
7].
The objectives of our study were to compare localization and expression of JAK1 and JAK3 in the skin of dust mite-allergic atopic dogs before and after epicutaneous allergen exposure. Since patients with naturally occurring disease can have a combination of immunologically acute and chronic lesions, the availability of a research model in which the lesions can be triggered by allergen exposure and monitored over time allowing sequential biopsies is of great benefit. A colony of atopic beagles that naturally develop atopic dermatitis and develop clinical flares after exposure to dust mites has been validated as a model for canine atopic dermatitis [
8,
9,
10]. Dogs belonging to this colony were used for this study, and sequential allergen challenges were conducted over the course of 28 days [
8,
9,
10]. Thus, the aim of our study was to investigate the localization of JAKs in canine atopic skin before (Day 0) and after allergen exposure (Day 28) to see if changes in the expression and distribution of JAKs occur. The interest to look at changes in the course of allergen exposure was motivated by the fact that JAK1 and JAK3 are modulated by signaling molecules that play an important role in modulating pruritus and inflammation [
2,
11,
12]. JAK/STAT signaling is inhibited by suppressors of cytokine signaling proteins (SOCS) [
13]. The SOCS proteins regulate the balance between T helper cells with SOCS3 expressed in Th2 cells and atopic patients [
13]. Therefore, the hypothesis of our study was that JAK expression in lesional atopic skin after allergen exposure is decreased compared to non-lesional atopic skin. Documentation of the expression of JAKs in canine skin is important to build the foundation for the pursuit of topical JAK inhibitors in veterinary medicine. The modulation of JAK expression upon allergen exposure is of particular interest for future therapeutic purposes.
4. Discussion
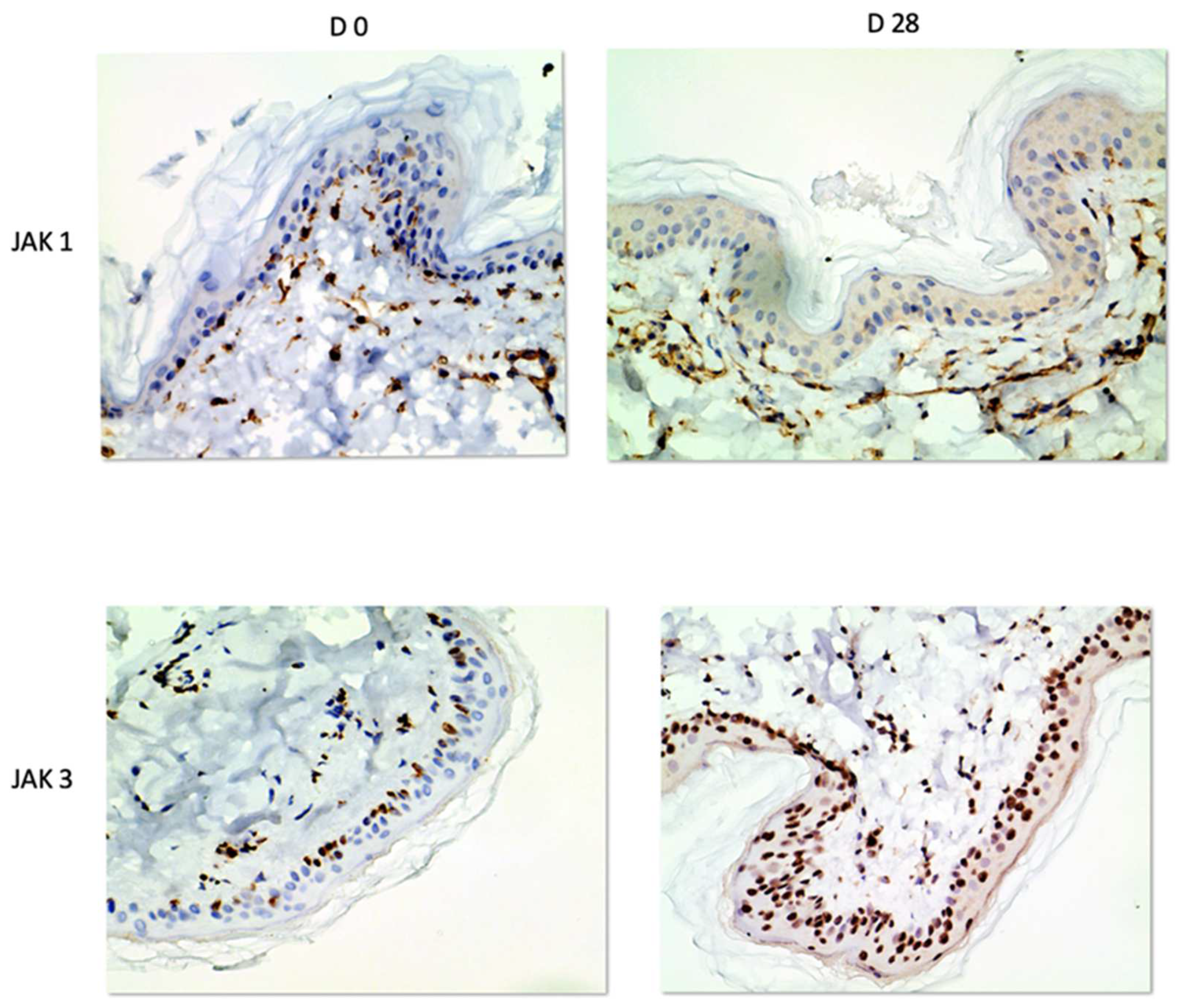
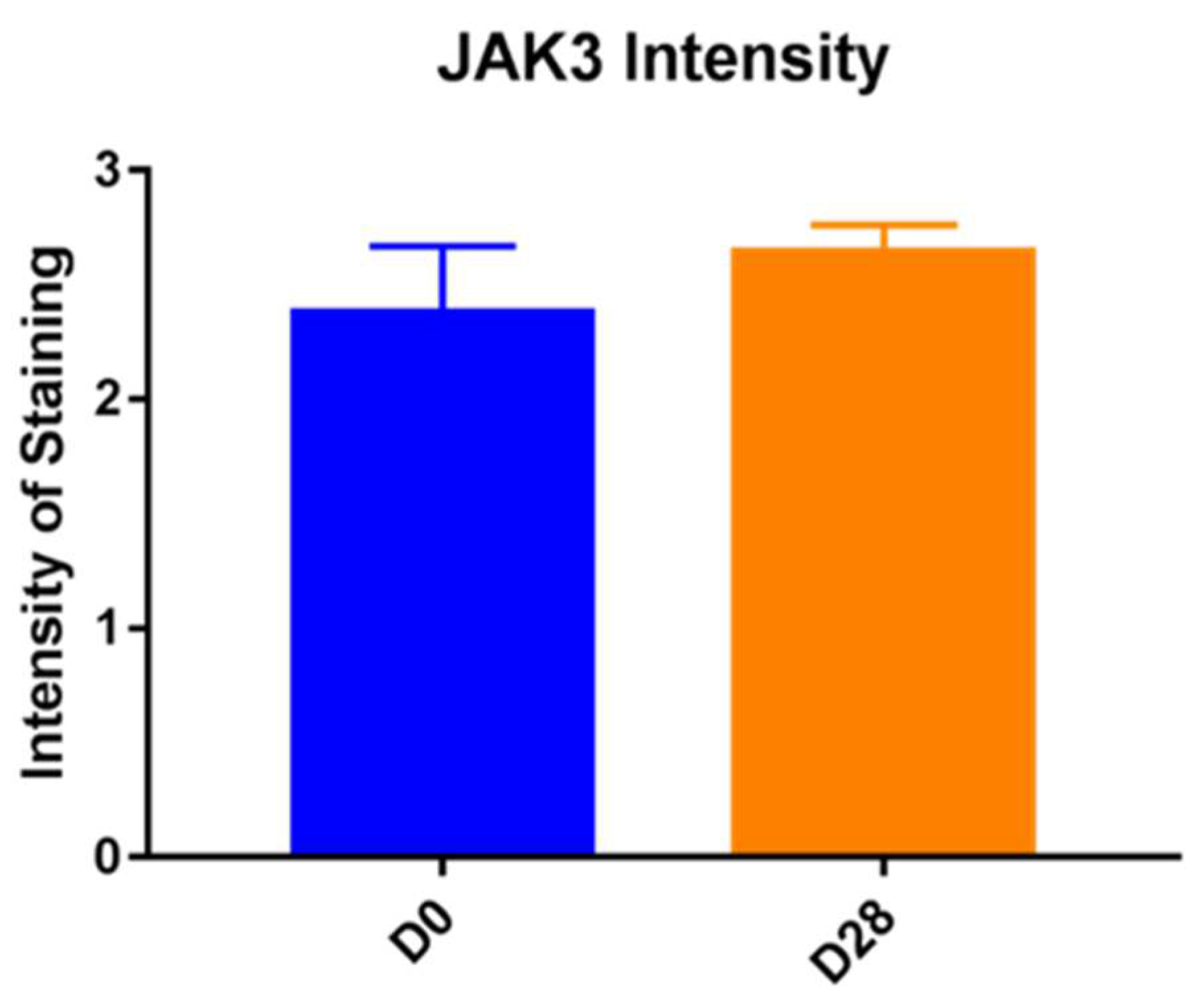
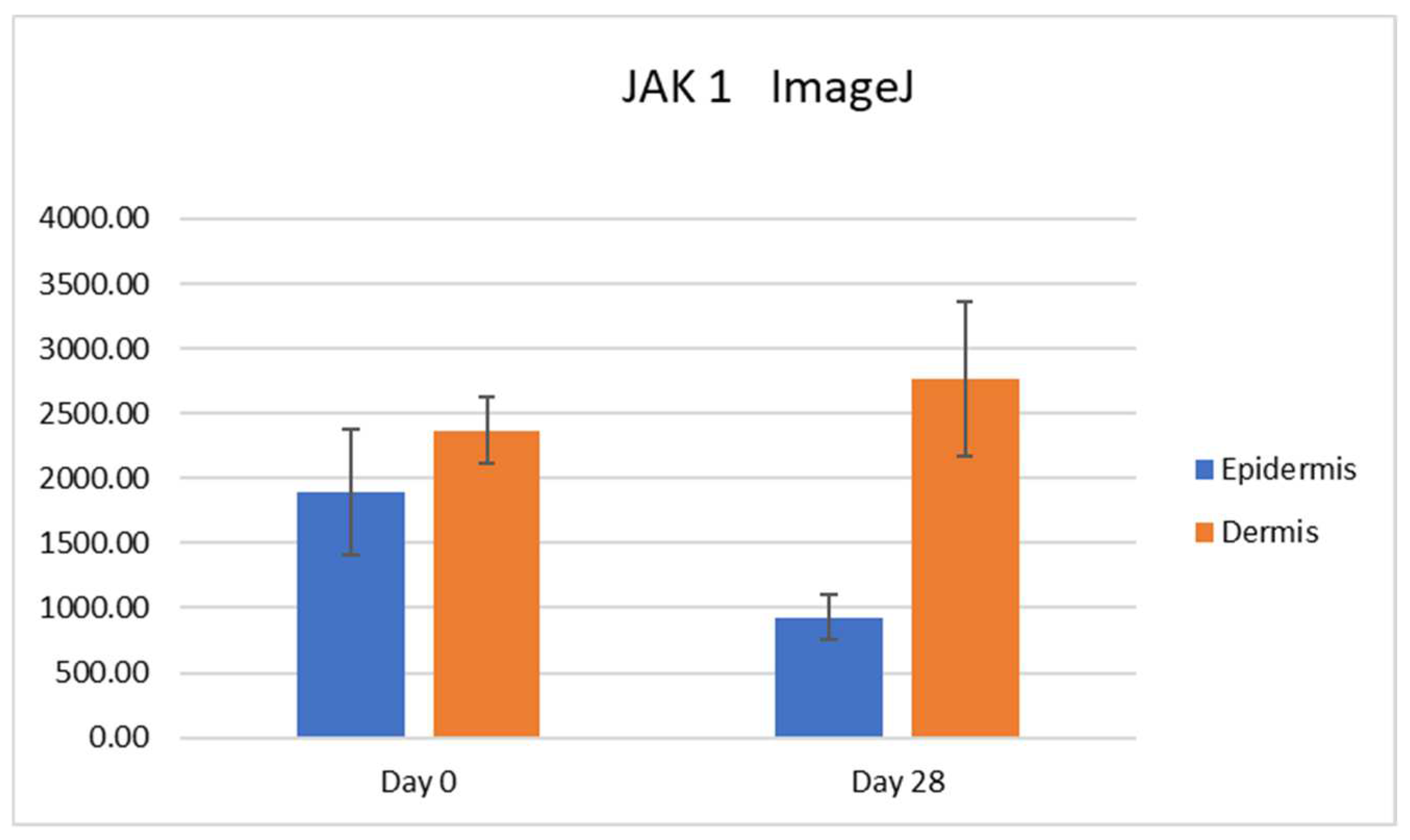
In our study, we were able to document the expression of JAK1 and JAK3 in canine atopic skin, and we found that there is a decreased expression of JAK1 in the epidermis and increased expression of JAK3 in the epidermis of house dust mite-allergic atopic dogs after allergen exposure. These changes are most likely due to inflammation, but it could also be due to the fact that the mites could have had a proteolytic effect, and it is possible that the results could also be induced by the mites rather than the allergic process. As we did not biopsy normal dogs challenged with dust mites, we are not able to sort out how much of that effect was linked to the allergic inflammation and how much was due to the presence of the mites on the skin. All biopsies used in our studies were taken from atopic dogs and even if the baseline biopsies were from non-lesional skin, atopic skin is always intrinsically different from normal skin.
The decreased JAK1 immunostaining after allergen exposure and flare of clinical signs could be explained by the fact that JAK/STAT signaling is inhibited by SOCS proteins [
1,
6,
13], overexpressed in atopic dermatitis [
13]. It is demonstrated in the literature that SOCS directly inhibits JAK1, JAK2, and TYK2, with similar affinity, but has no affinity for JAK3 [
16]; this could explain the JAK1/JAK3 imbalance at the end of the experiment, due to SOCS inhibition only on JAK1.
Stains for SOCS proteins were not carried out in our study, and therefore it is unknown if they are overexpressed also in atopic dogs. To the best of the author’s knowledge, the factors modulating the expression of JAK in dogs are not known. This is an important area of research moving forward as modulation of JAKs can play an important role in response to treatment. For example, there are some atopic dogs that simply are non-responsive to treatment with oclacitinib, and the exact reasons for that lack of response are not known. Similarly, there are dogs that are responsive in the early stages of treatment and then stop responding. Correlating the JAK1 expression and the clinical response to oclacitinib could be a very interesting approach to better understand the various clinical outcomes.
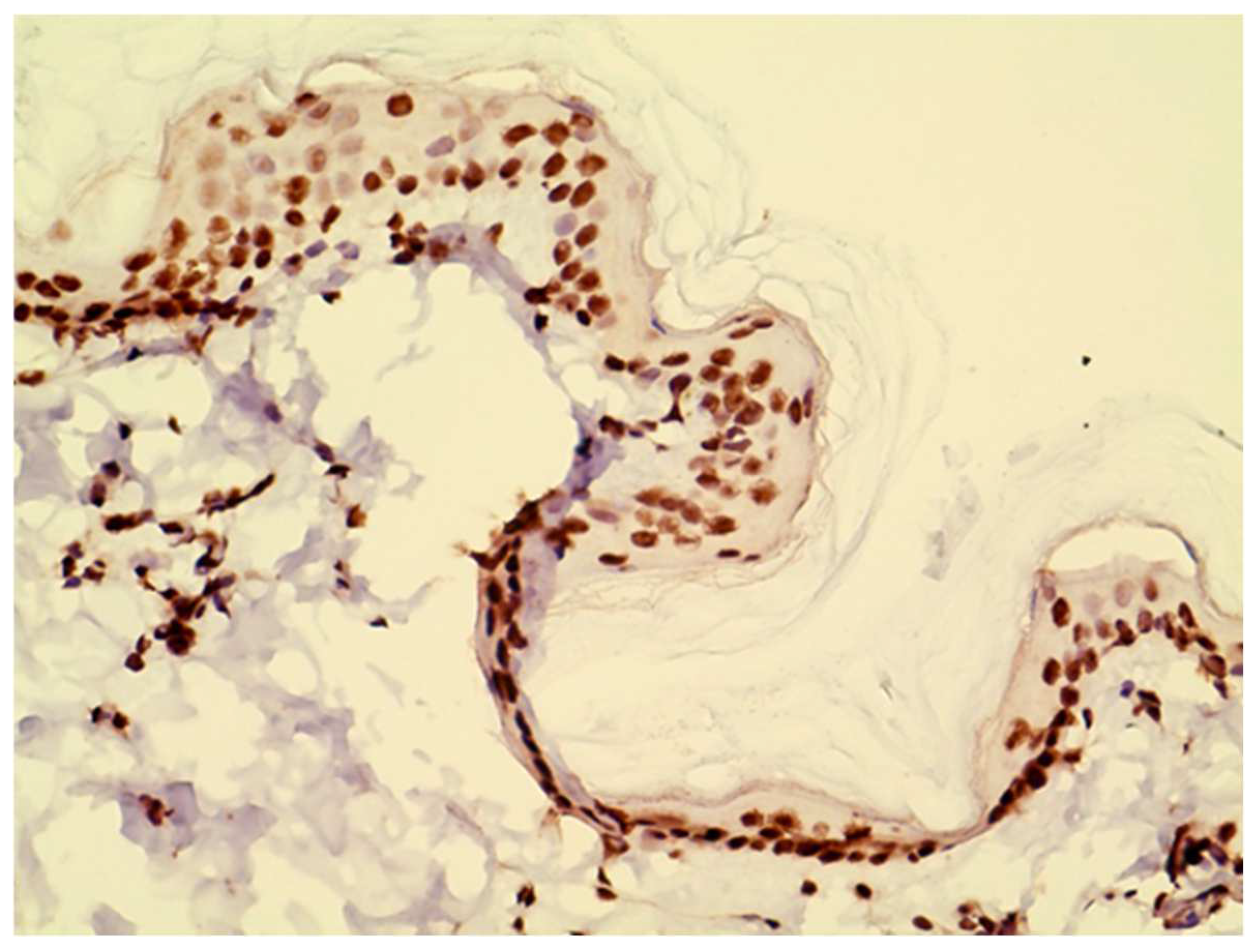
In our study, we found JAK1 and JAK3 positive cells in the dermal infiltrate of our biopsies. These cells were identified to be primarily lymphocytes and eosinophils based on Hematoxylin and Eosin staining. In hindsight, it would have been interesting to perform double staining to better characterize which cells express JAK1 versus JAK3 and how that changes during the course of allergen exposure.
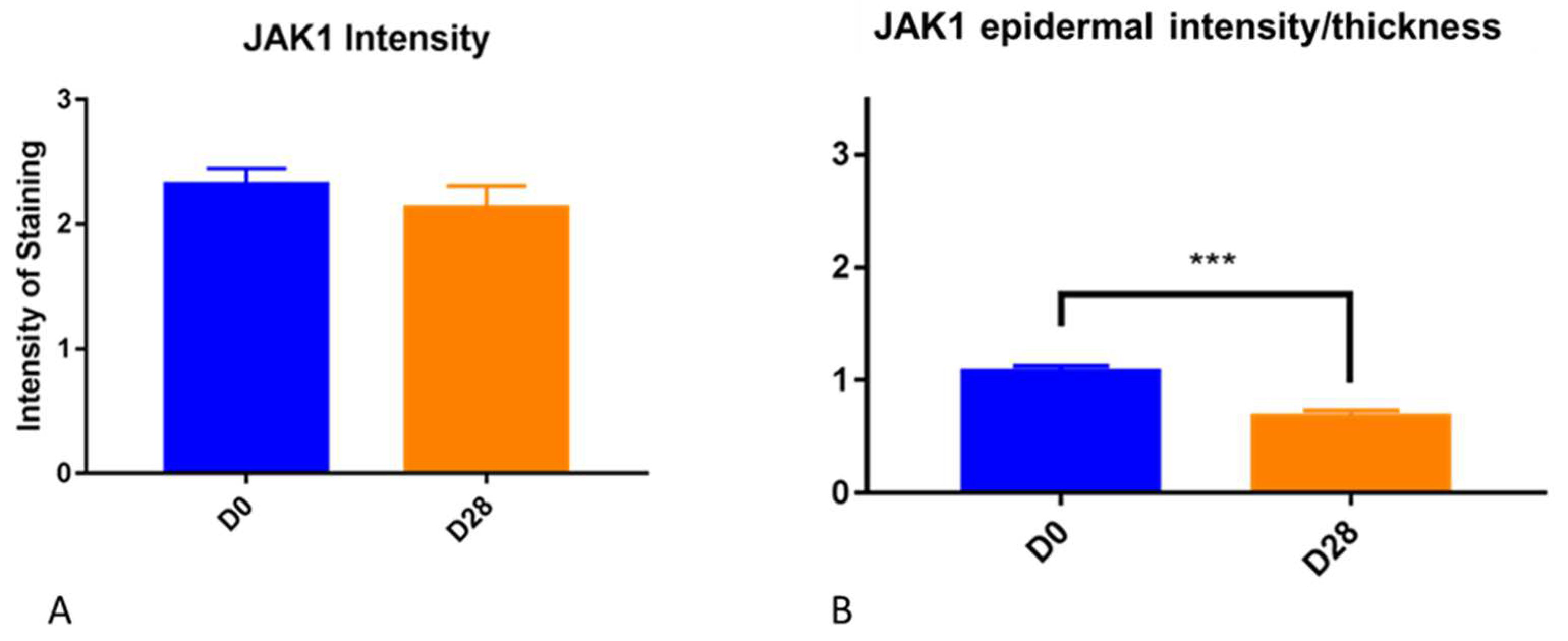
It is important to point out that the epidermis gets thicker as a result of allergen challenge and chronic inflammation. We have seen it in this experimental model for canine atopic dermatitis, and it is a common finding in people with atopic dermatitis. This factor needs to be taken into consideration when subjectively evaluating the intensity of immunohistochemistry in biopsy samples. The increased thickness of the epidermis may lead to increased intensity for subjective scores but when the scores are divided by the thickness, this effect is correct. Thus, we feel that the decrease in JAK1 intensity when the thickness factor was taken into consideration is a real finding.
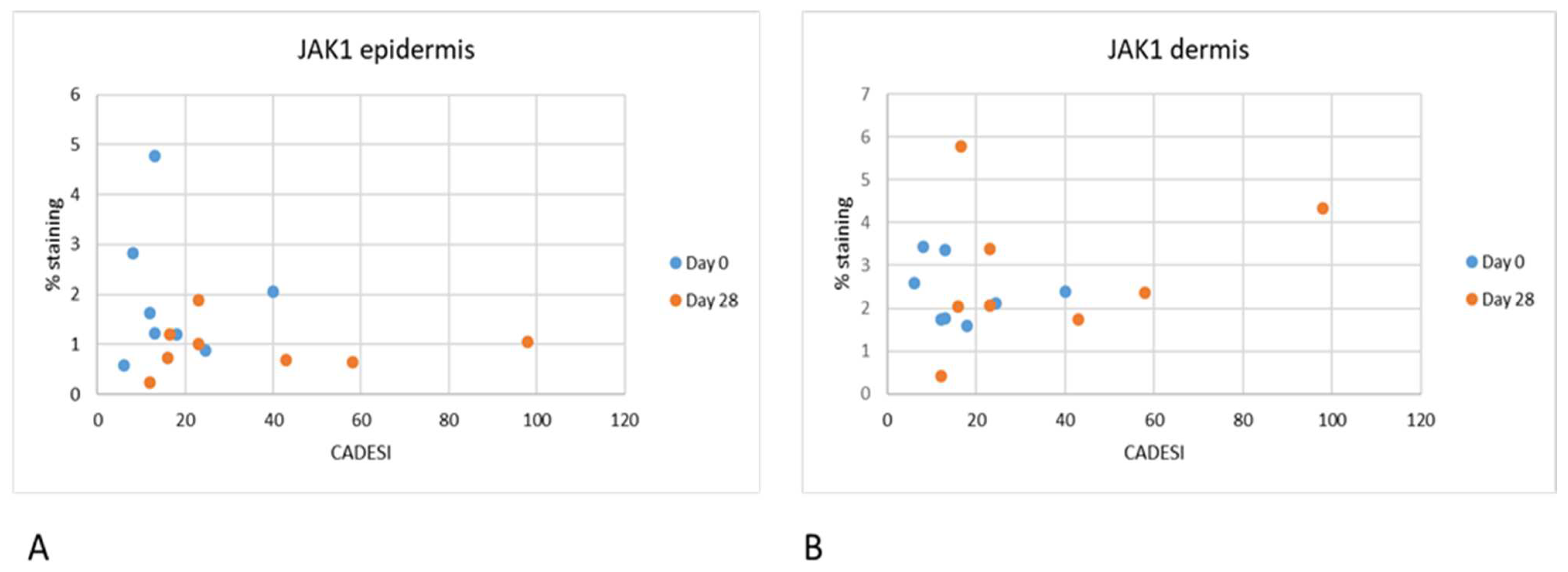
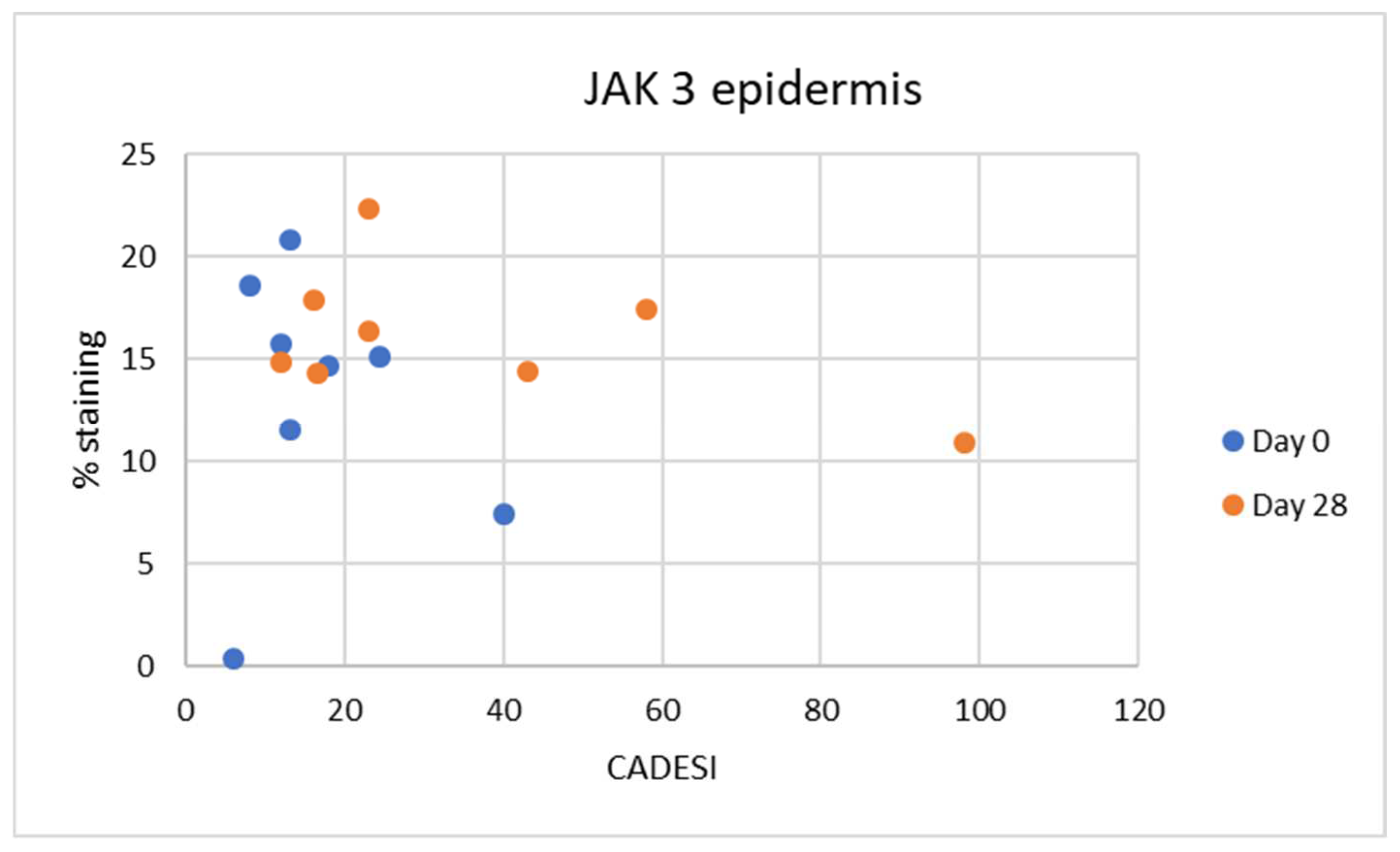
Interestingly in our study, we did not find any correlation between the severity of dermatitis scores and JAK expression. As our study was limited in size, it is possible that significant correlations can be found when examining a larger number of samples. It is also important to point out that the dermatitis scores used in this analysis were the scores for the whole body and not for the specific site that was biopsied. It is possible that with a larger number of samples and focusing on the clinical score of the specific site biopsied a correlation may be found.
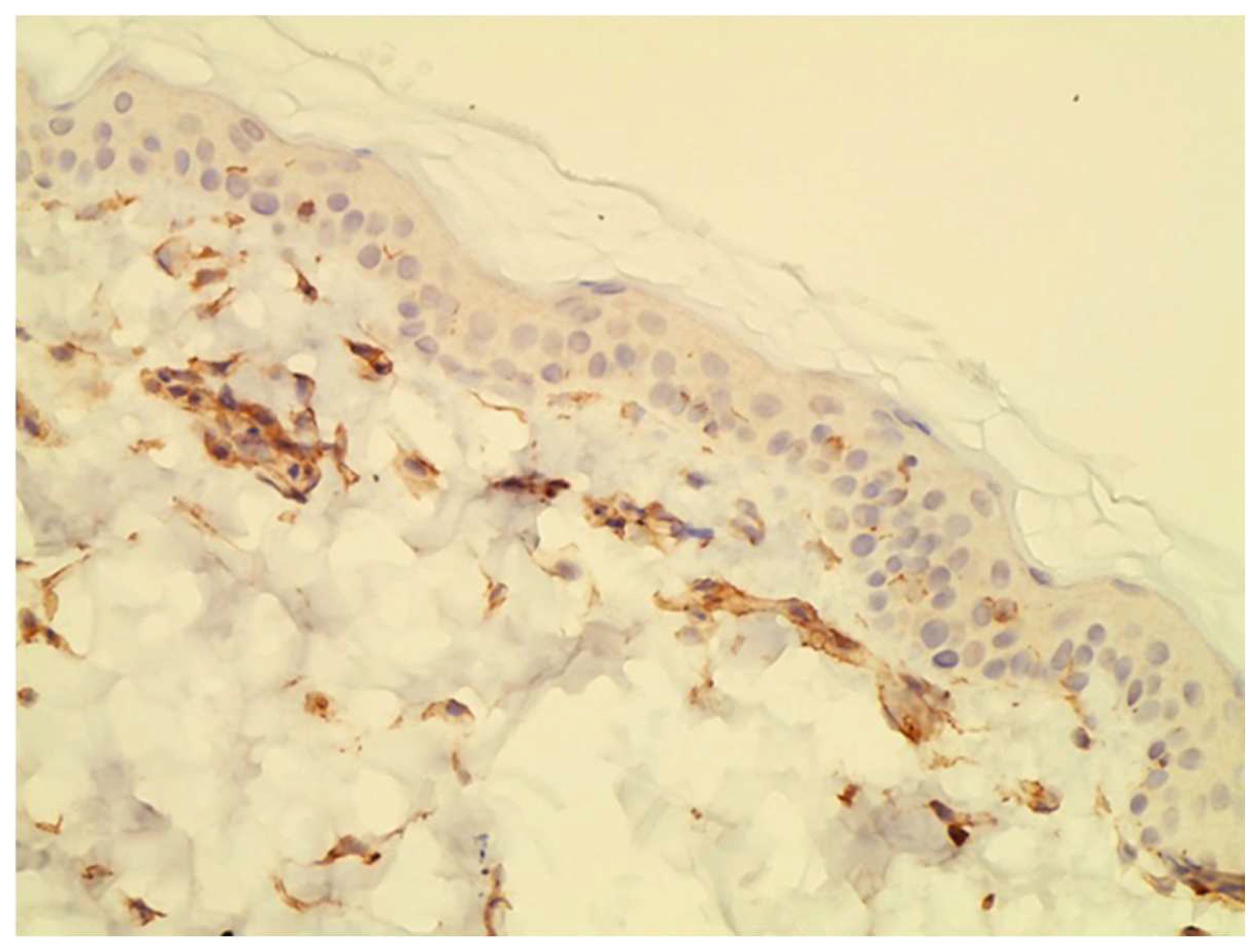
As far as immunolocalization is concerned, JAK1 was found in the cytoplasm, and this is the normal localization of JAK family enzymes [
6,
7]. In fact, after the attachment of the signaling molecule to its transmembrane receptor, activation of the enzymes associated with the cytoplasmic domain of the receptor JAK takes place. When activated, JAKs phosphorylate cytokine receptors, which enables STAT monomers present in the cytoplasm to bind to the complex and form homo- and heterodimers due to tyrosine phosphorylation. Then, activated STATs translocate to the cell nucleus and bind to DNA, enabling the transcription of target genes [
6].
In the epidermal layer, JAK3 was overexpressed on Day 28, indicating its crucial role in inflammatory keratinocytes as previously demonstrated [
11]. The epidermal expression levels of JAK3 suggest that its inhibition could be a promising approach for topical application [
11]. The expression patterns of JAKs have been evaluated in various skin diseases in people, including atopic dermatitis [
11]. In particular, phosphorylated and non-phosphorylated forms of JAK/STAT pathway members were analyzed in atopic dermatitis, psoriasis, lichen planus, cutaneous lupus erythematosus, pyoderma gangrenosum, and alopecia areata versus healthy controls. As far as JAK3 and pJAK3 expression is concerned, in the epidermis, it showed a higher expression in all inflammatory diseases except in cutaneous lupus erythematosus. JAK1 and phosphorylated JAK1 were only significantly overexpressed in pyoderma gangrenosum. No significantly different expression was seen for JAK2 and phosphorylated JAK2 in any of the investigated diseases. JAK3 was strongly expressed also in alopecia areata; JAK1 and JAK2 were to a lesser extent also elevated. The resulting downstream cascade seems to be based on the expression levels mainly conducted by phosphorylated STAT2. [
11] This is remarkable because alopecia areata is considered to be an IFN-γ driven disease which would result in pan-JAK activation. In this regard, it is important to notice that JAK3 was found to be the only JAK overexpressed in human alopecia areata compared to controls. In veterinary medicine, alopecia areata is also described. An investigation of JAK/STAT pathway members’ expression in this disease would be interesting in order to demonstrate the role of topical or systemic JAK inhibitors in disease regression, as anecdotally reported.
Compared to healthy skin, in the above-mentioned study [
11], the expression of both phosphorylated JAK1 and phosphorylated JAK3 was significantly enhanced in the dermal inflammatory cells of most diseases. This overexpression was more pronounced than in the epidermal keratinocytes. None of the inflammatory skin diseases is characterized by an increased expression of JAK2; phosphorylated JAK2 was overexpressed in atopic dermatitis and pyoderma gangrenosum. An interesting finding of the study [
11] is that the JAK3 pathway was most upregulated, followed by JAK1 and to a lesser extent JAK2. In the epidermal layer, JAK3 was overexpressed in all diseases (except cutaneous lupus erythematosus), indicating its crucial role in inflammatory keratinocytes. This suggests that selective JAK3 inhibition is the most promising approach for topical application in inflammatory skin diseases, as it can directly reverse the JAK profile of keratinocytes.
The expression of JAKs in the skin is of particular interest as it opens the possibility for topical treatments targeting these pathways. This approach allows a focus on where the suppression is needed, eliminating the need for systemic treatment and the risk of adverse effects. Studies regarding topical JAK inhibitors (tofacitinib, ruxolitinib, brepocitinib) to treat atopic dermatitis in humans are ongoing [
17,
18]. Ruxolitinib cream has met all primary and secondary endpoints in phase 3 clinical trials for mild-to-moderate atopic dermatitis with minimal treatment-emergent adverse events. Delgocitinib is an approved topical pan-JAK inhibitor for treating atopic dermatitis in adults and children [
17]. Based on the structural and functional characteristics of JAKs, as well as specific tissue distribution, JAK3, in particular, has emerged as an ideal target for the treatment of inflammatory or autoimmune diseases [
17] as previously demonstrated. To date, no selective JAK3 inhibitor has yet been approved for the treatment of inflammatory and autoimmune diseases [
17]. Topical JAK inhibitors are found to be effective and safe in the treatment of atopic dermatitis, and they were observed to significantly improve clinical signs, pain, and pruritus scores compared to placebo in human patients [
19].
Moreover, topical JAK inhibitors have a lower risk of adverse effects compared to systemic JAK inhibitors or other immunomodulatory drugs used to treat the disease. The majority of reactions are localized to the application site, and treatment discontinuation is not needed, making this type of treatment appealing. Other topical options for atopic dermatitis are glucocorticoids and calcineurin inhibitors. Topical glucocorticoids have the undesirable adverse effect of cutaneous atrophy, and topical calcineurin inhibitors frequently cause a stinging sensation, at least at the beginning of therapy.
In terms of comparing the efficacy of topical and systemic JAK inhibitors in the treatment of atopic dermatitis, it has been reported that topical forms are significantly more effective than systemic. In particular tofacitinib, ruxolitinib, and delgocitinib had higher achievement in the investigator’s global assessment response compared to topical tacrolimus and phosphodiesterase-4 inhibitors. [
17]. Meanwhile, tofacitinib was reported to possess the highest response among the rest of the included medications [
17].
JAK3 had the same nuclear staining pattern as reported in previous studies [
3,
4,
10] possibly reflecting nuclear translocation [
18]. In a previous study [
11], all JAKs and their phosphorylated forms were positive in the epidermis and dermis of inflammatory skin diseases, and they showed a cytoplasmic expression along the cell membrane of keratinocytes, with the exception of pJAK2, which was nuclear. TYK2 and pTYK2 were expressed in the nucleus and in the cytoplasm, with pTYK2 nuclear expression being more pronounced than the cytoplasmatic one. There is evidence that these kinases phosphorylate not only other transcription factors besides the STATs, but that the JAKs function as epigenetic regulators of gene expression. Emerging evidence indicates that the latter role may be of particular significance under physiological and pathological conditions of heightened cellular growth [
19]. This was confirmed in a recent publication that reported on JAK3 expression in the nucleus of malignant T cells [
20].
Limitations of our study included the small number of dogs and the lack of a normal dog population similarly challenged with the allergen. An additional potential limitation is that an antibody raised against human JAKs was used rather than a specific anti-canine antibody. To the best of the authors’ knowledge, such an antibody is not commercially available. The antibody used is advertised to react to canine JAK, but it is possible that it could bind and cross-react with other proteins. In our study, we did not perform investigations also on phosphorylated forms of JAKs as reported in previous studies [
11]. Further studies are needed in order to better determine the phosphorylation of these enzymes and thus their activation or inactivation status.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







