
Characterization of BoHV-1 gG-/tk-/gE- Mutant in Differential Protein Expression, Virulence, and Immunity

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture and Virus Strains
2.3. Construction of Triple Mutant Virus BoHV-1 Strain gG-/tk-/gE-
2.3.1. Construction of Transfer Vector pBoHV-1 gE-
2.3.2. Generation and Characterization of the Triple Mutant Virus
2.4. Viral Growth Kinetics and Plaque Size Determination
2.5. Label Free Quantitative Proteomics (LFQP)
2.5.1. Virions Purification
2.5.2. LC-MS/MS Analysis
2.5.3. Database Search and Bioinformatic Analysis
2.6. Cattle Experiments
2.6.1. Clinical Evaluation
2.6.2. Virus Isolation and Titration
2.6.3. Serological Investigation
2.6.4. Histo-Pathological Examination of Lungs
2.7. Statistical Analysis
3. Results
3.1. Construction and Characterization of the Recombinant BoHV-1 gG-/tk-/gE- Triple- Deleted Mutant
3.2. Comparative Viral Growth Kinetics and Plaque Size Estimation
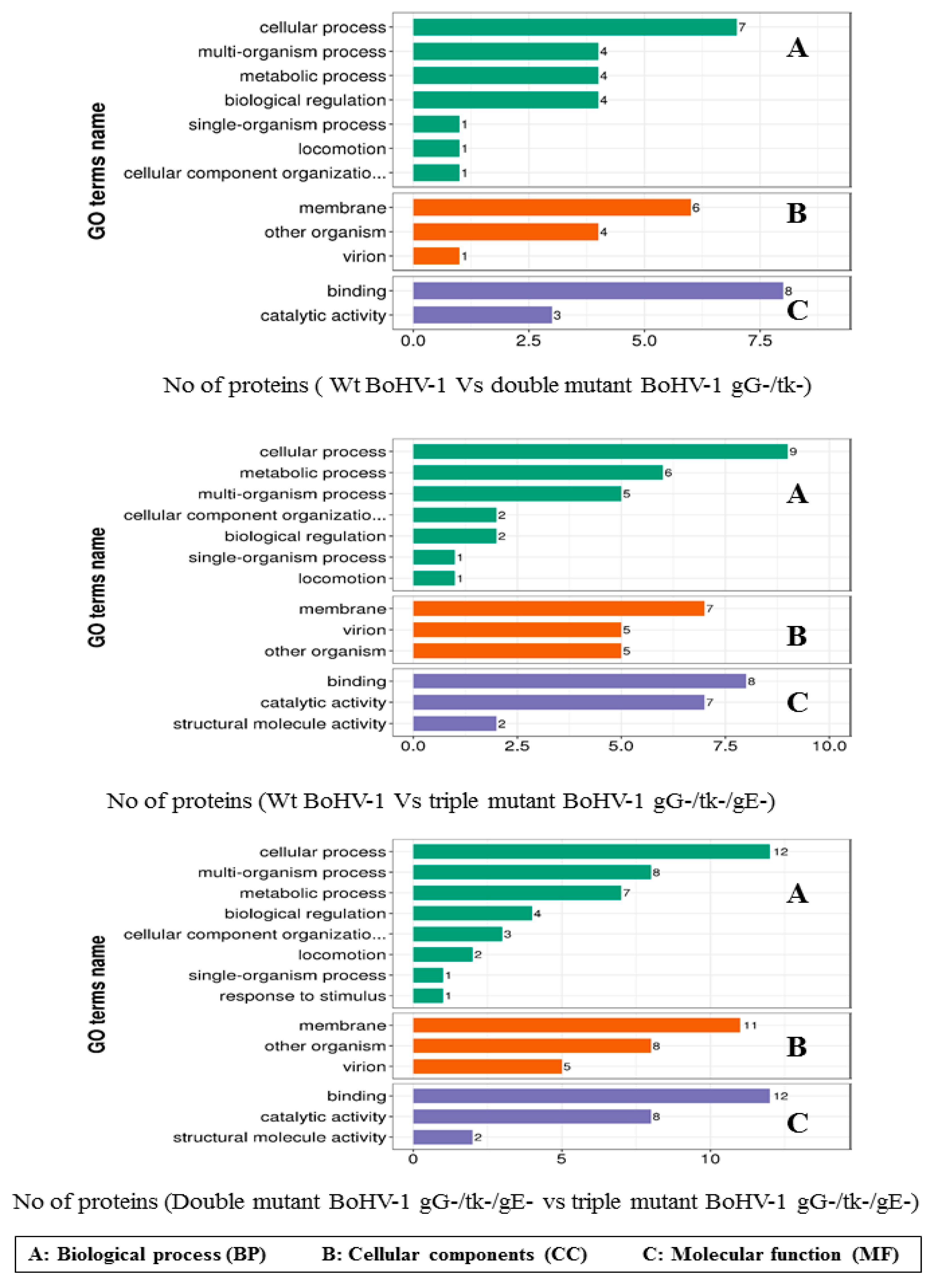
3.3. Comparative Proteomics of the Three BoHV-1 Strains
3.3.1. Differentially Expressed Proteins in Different Comparable Strains
3.3.2. GO Secondary Annotation Classification
3.4. In Vivo Experiment Findings
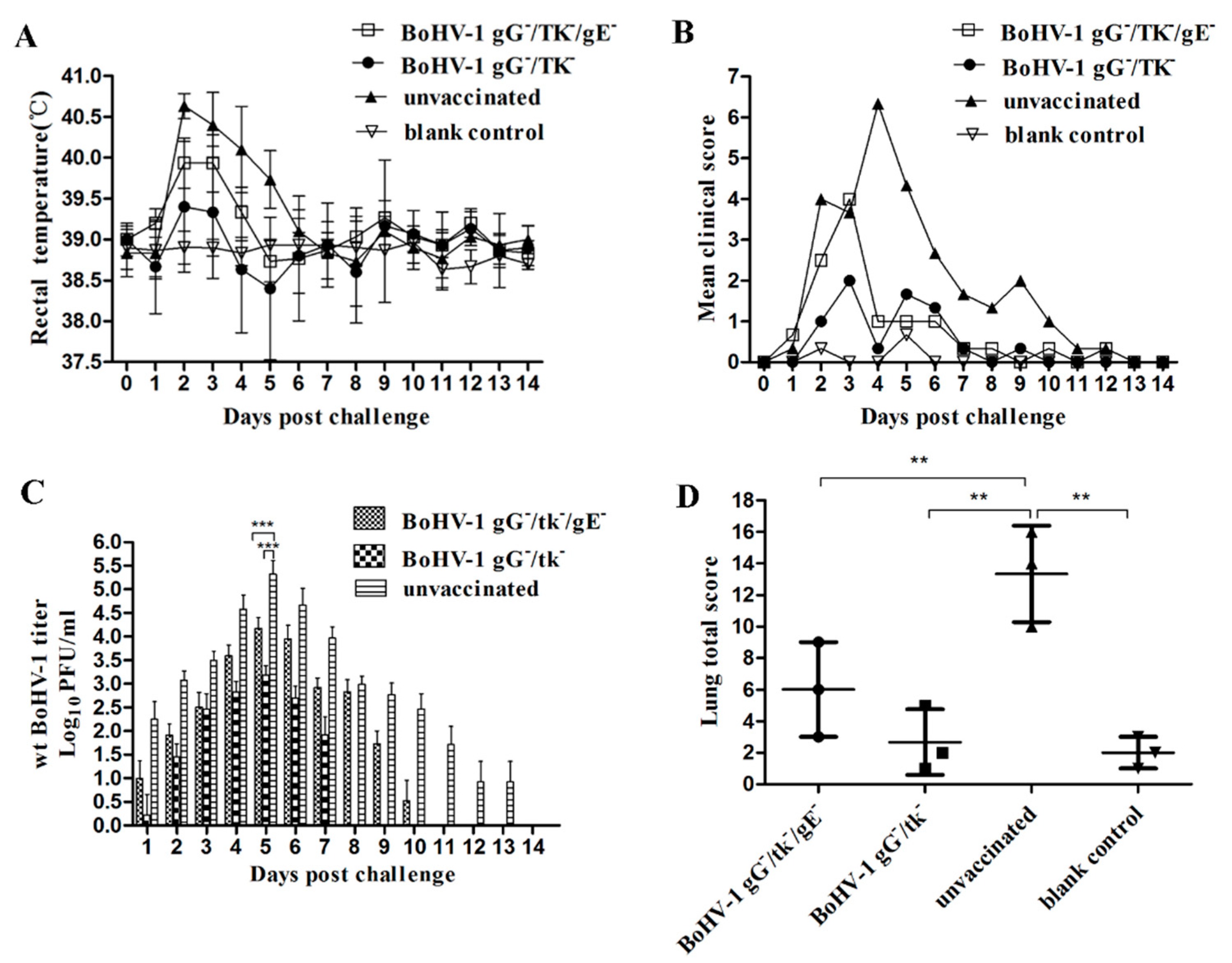
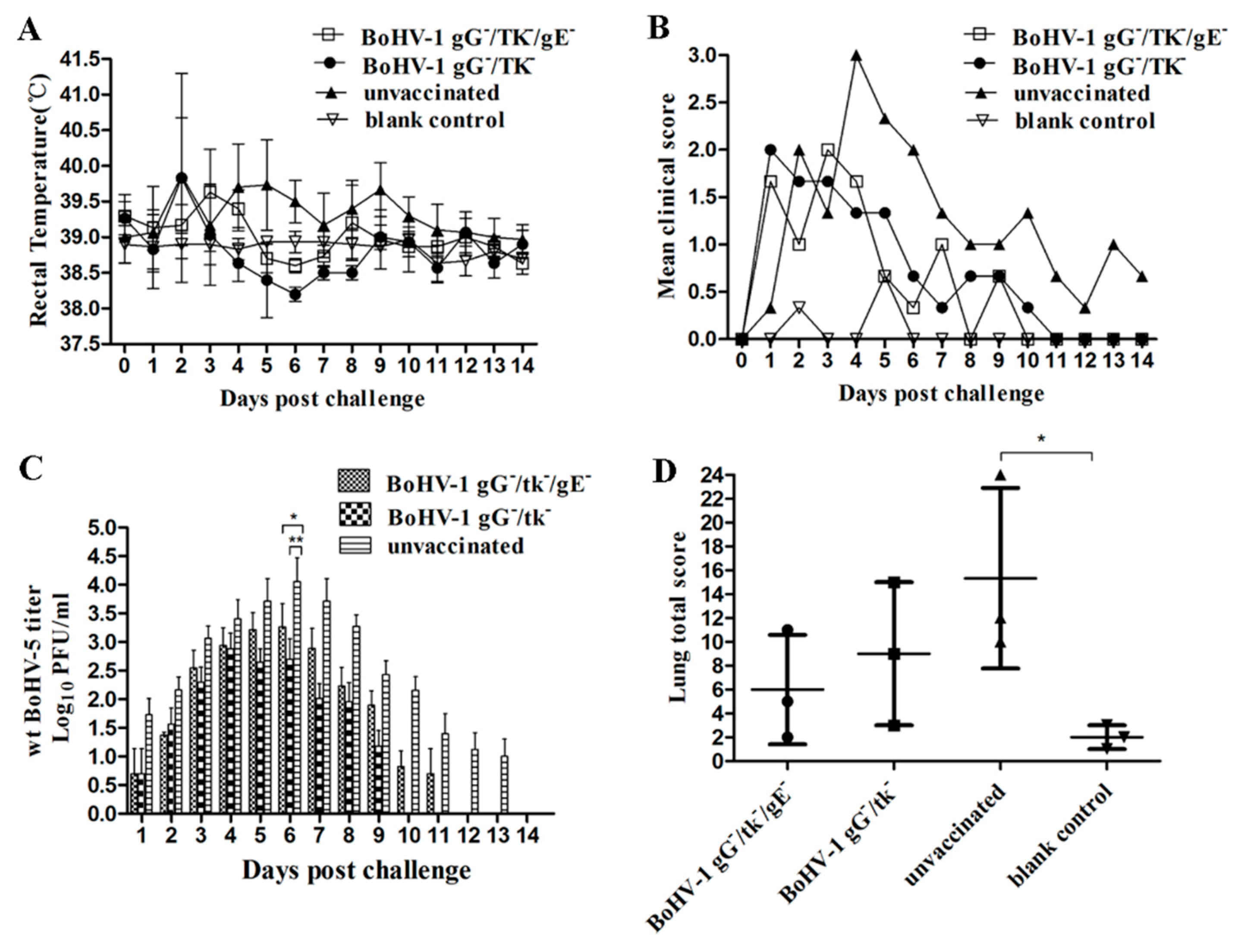
3.4.1. Virulence and Reactivation of BoHV-1 gG-/tk-/gE- in Calves
3.4.2. Protection of BoHV-1 gG-/tk-/gE- against wt BoHV-1 Challenge in Calves
3.4.3. Cross-Protection of BoHV-1 gG-/tk-/gE- against wt BoHV-5 Challenge in Calves
3.5. Serological Investigations
3.5.1. Humoral Immune Response in Calves
3.5.2. Cytokine Production in Calves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biswas, S.; Bandyopadhyay, S.; Dimri, U.; Patra, P.H. Bovine herpesvirus-1 (BHV-1)-a re-emerging concern in livestock: A revisit to its biology, epidemiology, diagnosis, and prophylaxis. Vet. Q. 2013, 33, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Engel, E.A.; Song, R.; Koyuncu, O.O.; Enquist, L.W. Investigating the biology of alpha herpesviruses with MS-based proteomics. Proteomics 2015, 15, 1943–1956. [Google Scholar] [CrossRef] [Green Version]
- Muylkens, B.; Thiry, J.; Kirten, P.; Schynts, F.; Thiry, E. Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet. Res. 2007, 38, 181–209. [Google Scholar] [CrossRef] [Green Version]
- D’Arce, R.C.; Almeida, R.S.; Silva, T.C.; Franco, A.C.; Spilki, F.; Roehe, P.M.; Arns, C.W. Restriction endonuclease and monoclonal antibody analysis of Brazilian isolates of bovine herpesviruses types 1 and 5. Vet. Microbiol. 2002, 88, 315–324. [Google Scholar] [CrossRef]
- Jones, C. Bovine Herpesvirus 1 Counteracts Immune Responses and Immune-Surveillance to Enhance Pathogenesis and Virus Transmission. Front. Immunol. 2019, 10, 1008. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, C.; Bratanich, A. The latency related gene of bovine herpesvirus types 1 and 5 and its modulation of cellular processes. Arch. Virol. 2016, 161, 3299–3308. [Google Scholar] [CrossRef]
- Guo, J.; Li, Q.; Jones, C. The bovine herpesvirus 1 regulatory proteins, bICP4 and bICP22, are expressed during the escape from latency. J. Neurovirol. 2019, 25, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Petrini, S.; Iscaro, C.; Righi, C. Antibody Responses to Bovine Alphaherpesvirus 1 (BoHV-1) in Passively Immunized Calves. Viruses 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.E.; Meers, J.; Gravel, J.L.; McCarthy, F.M.; Mahony, T.J. The essential and non-essential genes of Bovine herpesvirus 1. J. Gen. Virol. 2008, 89, 2851–2863. [Google Scholar] [CrossRef]
- Van Engelenburg, F.A.; Kaashoek, M.J.; van Oirschot, J.T.; Rijsewijk, F.A. A glycoprotein E deletion mutant of bovine herpesvirus 1 infects the same limited number of tissues in calves as wild-type virus, but for a shorter period. J. Gen. Virol. 1995, 76 Pt 9, 2387–2392. [Google Scholar] [CrossRef]
- Strube, W.; Auer, S.; Block, W.; Heinen, E.; Kretzdorn, D.; Rodenbach, C.; Schmeer, N. A gE deleted infectious bovine rhinotracheitis marker vaccine for use in improved bovine herpesvirus 1 control programs. Vet. Microbiol. 1996, 53, 181–189. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; van Engelenburg, F.A.; Moerman, A.; Gielkens, A.L.; Rijsewijk, F.A.; van Oirschot, J.T. Virulence and immunogenicity in calves of thymidine kinase- and glycoprotein E-negative bovine herpesvirus 1 mutants. Vet. Microbiol. 1996, 48, 143–153. [Google Scholar] [CrossRef]
- Van Engelenburg, F.A.; Kaashoek, M.J.; Rijsewijk, F.A.; van den Burg, L.; Moerman, A.; Gielkens, A.L.; van Oirschot, J.T. A glycoprotein E deletion mutant of bovine herpesvirus 1 is avirulent in calves. J. Gen. Virol. 1994, 75 Pt 9, 2311–2318. [Google Scholar] [CrossRef]
- Belknap, E.B.; Walters, L.M.; Kelling, C.; Ayers, V.K.; Norris, J.; McMillen, J.; Hayhow, C.; Cochran, M.; Reddy, D.N.; Wright, J.; et al. Immunogenicity and protective efficacy of a gE, gG and US2 gene-deleted bovine herpesvirus-1 (BHV-1) vaccine. Vaccine 1999, 17, 2297–2305. [Google Scholar] [CrossRef]
- Chowdhury, S.I.; Wei, H.; Weiss, M.; Pannhorst, K.; Paulsen, D.B. A triple gene mutant of BoHV-1 administered intranasally is significantly more efficacious than a BoHV-1 glycoprotein E-deleted virus against a virulent BoHV-1 challenge. Vaccine 2014, 32, 4909–4915. [Google Scholar] [CrossRef] [PubMed]
- Nunez Villavicencio-Diaz, T.; Rodríguez-Ulloa, A.; Guirola-Cruz, O.; Perez-Riverol, Y. Bioinformatics tools for the functional interpretation of quantitative proteomics results. Curr. Top. Med. Chem. 2014, 14, 435–449. [Google Scholar] [CrossRef]
- Zhang, M.; Fu, S.; Deng, M.; Xie, Q.; Xu, H.; Liu, Z.; Hu, C.; Chen, H.; Guo, A. Attenuation of bovine herpesvirus type 1 by deletion of its glycoprotein G and tk genes and protection against virulent viral challenge. Vaccine 2011, 29, 8943–8950. [Google Scholar] [CrossRef] [PubMed]
- Barber, K.; Daugherty, H.; Ander, S.; Jefferson, V.; Shack, L.; Pechan, T.; Nanduri, B.; Meyer, F. Protein composition of the bovine herpesvirus 1.1 virion. Vet. Sci. 2017, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Al-Mubarak, A.; Simon, J.; Coats, C.; Okemba, J.D.; Burton, M.D.; Chowdhury, S.I. Glycoprotein E (gE) specified by bovine herpesvirus type 5 (BHV-5) enables trans-neuronal virus spread and neurovirulence without being a structural component of enveloped virions. J. Virol. 2007, 365, 398–409. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Cai, T.; Li, T.; Xue, P.; Zhou, B.; He, X.; Wei, P.; Liu, P.; Yang, F.; Wei, T. Lysine malonylation is elevated in type 2 diabetic mouse models and enriched in metabolic associated proteins. Mol. Cell. Proteom. 2015, 14, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Valera, A.R.; Pidone, C.L.; Massone, A.R.; Quiroga, M.A.; Riganti, J.G.; Corva, S.G.; Galosi, C.M. A simple method of infecting rabbits with Bovine herpesvirus 1 and 5. J. Virol. Methods 2008, 150, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; He, J.; Paulsen, D.B.; Chowdhury, S.I. Bovine herpesvirus type 1 (BHV-1) mutant lacking U(L)49.5 luminal domain residues 30–32 and cytoplasmic tail residues 80–96 induces more rapid onset of virus neutralizing antibody and cellular immune responses in calves than the wild-type strain Cooper. Vet. Immunol. Immunopathol. 2012, 147, 223–229. [Google Scholar] [CrossRef]
- Chowdhury, S.I. Construction and characterization of an attenuated bovine herpesvirus type 1 (BHV-1) recombinant virus. Vet. Microbiol. 1996, 52, 13–23. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; Villarreal-Ramos, B.; Cockle, P.J.; McAulay, M.; Rhodes, S.G.; Thacker, T.; Gilbert, S.C.; McShane, H.; Hill, A.V.; Xing, Z. Viral booster vaccines improve Mycobacterium bovis BCG-induced protection against bovine tuberculosis. Infect. Immun. 2009, 77, 3364–3373. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Han, X.; Chen, Y.; Mustafa, R.; Qi, J.; Chen, X.; Hu, C.; Chen, H.; Guo, A. Attenuated Mycoplasma bovis strains provide protection against virulent infection in calves. Vaccine 2014, 32, 3107–3114. [Google Scholar] [CrossRef]
- Ackermann, M.; Engels, M. Pro and contra IBR-eradication. Vet. Microbiol. 2006, 113, 293–302. [Google Scholar] [CrossRef]
- Uddowla, S.; Hollister, J.; Pacheco, J.M.; Rodriguez, L.L.; Rieder, E. A safe foot-and-mouth disease vaccine platform with two negative markers for differentiating infected from vaccinated animals. J. Virol. 2012, 86, 11675–11685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.; Chowdhury, S. A review of the biology of bovine herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex and development of improved vaccines. Anim. Health Res. Rev. 2007, 8, 187–205. [Google Scholar] [CrossRef]
- Rho, J.H.; Qin, S.; Wang, J.Y.; Roehrl, M.H. Proteomic expression analysis of surgical human colorectal cancer tissues: Up-regulation of PSB7, PRDX1, and SRP9 and hypoxic adaptation in cancer. J. Proteome Res. 2008, 7, 2959–2972. [Google Scholar] [CrossRef] [Green Version]
- Alfonso, P.; Núñez, A.; Madoz-Gurpide, J.; Lombardia, L.; Sánchez, L.; Casal, J.I. Proteomic expression analysis of colorectal cancer by two-dimensional differential gel electrophoresis. Proteomics 2005, 5, 2602–2611. [Google Scholar] [CrossRef]
- Chowdhury, S.I.; Ross, C.S.; Lee, B.J.; Hall, V.; Chu, H.J. Construction and characterization of a glycoprotein E gene-deleted bovine herpesvirus type 1 recombinant. Am. J. Vet. Res. 1999, 60, 227–232. [Google Scholar]
- Schwyzer, M.; Ackermann, M. Molecular virology of ruminant herpesviruses. Vet. Microbiol. 1996, 53, 17–29. [Google Scholar] [CrossRef]
- Muylkens, B.; Meurens, F.; Schynts, F.; Farnir, F.; Pourchet, A.; Bardiau, M.; Gogev, S.; Thiry, J.; Cuisenaire, A.; Vanderplasschen, A.; et al. Intraspecific bovine herpesvirus 1 recombinants carrying glycoprotein E deletion as a vaccine marker are virulent in cattle. J. Gen. Virol. 2006, 87, 2149–2154. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; Moerman, A.; Madic, J.; Rijsewijk, F.A.; Quak, J.; Gielkens, A.L.; van Oirschot, J.T. A conventionally attenuated glycoprotein E-negative strain of bovine herpesvirus type 1 is an efficacious and safe vaccine. Vaccine 1994, 12, 439–444. [Google Scholar] [CrossRef]
- Shaw, A.; Braun, L.; Frew, T.; Hurley, D.; Rowland, R.; Chase, C. A role for bovine herpesvirus 1 (BHV-1) glycoprotein E (gE) tyrosine phosphorylation in replication of BHV-1 wild-type virus but not BHV-1 gE deletion mutant virus. Virology 2000, 268, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrini, S.; Righi, C.; Iscaro, C.; Viola, G.; Gobbi, P.; Scoccia, E.; Rossi, E.; Pellegrini, C.; De Mia, G.M. Evaluation of Passive Immunity Induced by Immunisation Using Two Inactivated gE-deleted Marker Vaccines against Infectious Bovine Rhinotracheitis (IBR) in Calves. Vaccines 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Ampe, B.; Duchateau, L.; Speybroeck, N.; Berkvens, D.; Dupont, A.; Kerkhofs, P.; Thiry, E.; Dispas, M. Assessment of the long-term effect of vaccination on transmission of infectious bovine rhinotracheitis virus in cattle herds hyperimmunized with glycoprotein E-deleted marker vaccine. Am. J. Vet. Res. 2012, 73, 1787–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kholy, A.A.; Rady, D.I.; Abdou, E.R.; Elseafy, M.M.; Abdelrahman, K.A.; Soliman, H. Construction, characterization and immunogenicity of a glycoprotein E negative bovine herpesvirus-1.1 Egyptian strain “Abu-Hammad”. J. Virol. Methods 2013, 194, 74–81. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; Moerman, A.; Madic, J.; Weerdmeester, K.; Maris-Veldhuis, M.; Rijsewijk, F.A.; van Oirschot, J.T. An inactivated vaccine based on a glycoprotein E-negative strain of bovine herpesvirus 1 induces protective immunity and allows serological differentiation. Vaccine 1995, 13, 342–346. [Google Scholar] [CrossRef]
- Anziliero, D.; Santos, C.M.; Brum, M.C.; Weiblen, R.; Chowdhury, S.I.; Flores, E.F. A recombinant bovine herpesvirus 5 defective in thymidine kinase and glycoprotein E is immunogenic for calves and confers protection upon homologous challenge and BoHV-1 challenge. Vet. Microbiol. 2011, 154, 14–22. [Google Scholar] [CrossRef]
- Del Medico Zajac, M.P.; Ladelfa, M.F.; Kotsias, F.; Muylkens, B.; Thiry, J.; Thiry, E.; Romera, S.A. Biology of bovine herpesvirus 5. Vet. J. 2010, 184, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Del Medico Zajac, M.P.; Puntel, M.; Zamorano, P.I.; Sadir, A.M.; Romera, S.A. BHV-1 vaccine induces cross-protection against BHV-5 disease in cattle. Res. Vet. Sci. 2006, 81, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.D.; Spilki, F.R.; Franco, A.C.; Esteves, P.A.; Hubner, S.O.; Driemeier, D.; Oliveira, A.P.; Rijsewijk, F.; Roehe, P.M. Vaccination with a gE-negative bovine herpesvirus type 1 vaccine confers insufficient protection to a bovine herpesvirus type 5 challenge. Vaccine 2006, 24, 3313–3320. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
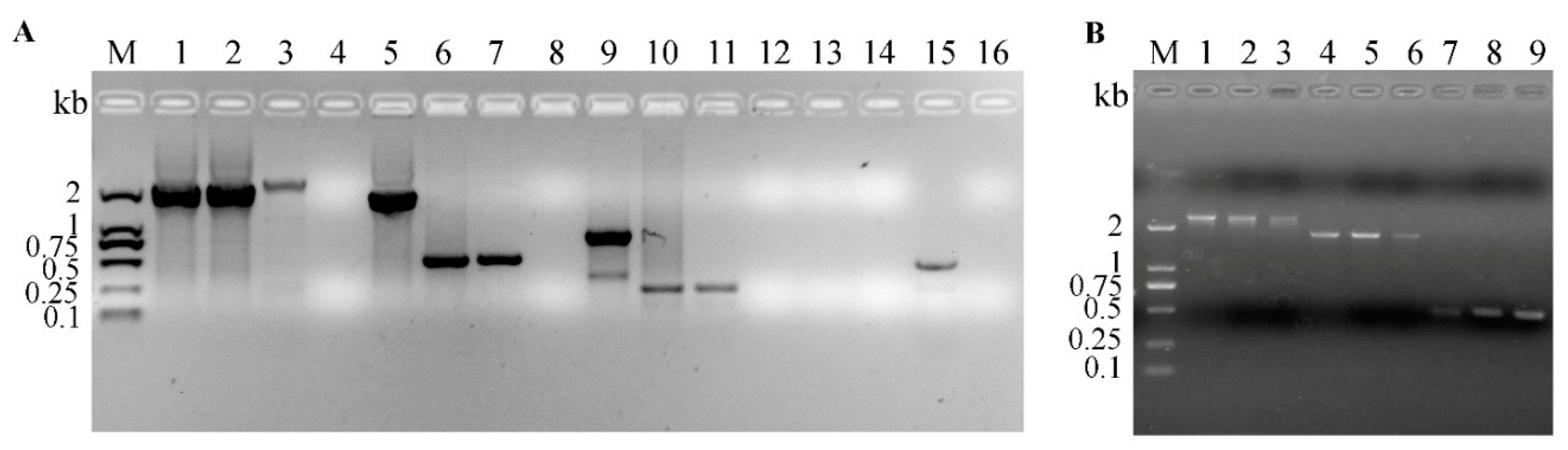{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal | Virus | No | Vaccinated Route and Dose | Dex Injection a | Challenge Virus | Challenge Date and Dose | Euthanasia and Necropsy |
|---|---|---|---|---|---|---|---|
| Calf | BoHV-1 gG-/tk-/gE- | 6 | IN, 4 × 107 PFU | 21–25 dpi | wt BoHV-1 (3) b wt BoHV-5 (3) b | 35 dpi, 4 × 107 PFU | 28 dpc |
| BoHV-1 gG-/tk- | 6 | IN, 4 × 107 PFU | 21–25 dpi | wt BoHV-1 (3) b wt BoHV-5 (3) b | 35 dpi, 4 × 107 PFU | 28 dpc | |
| wt BoHV-1 | 3 | IN, 4 × 107 PFU | 21–25 dpi | 35 dpi, 4 × 107 PFU | 28 dpc | ||
| Unvaccinated control | 6 | DMEM | 21–25 dpi | wt BoHV-1 (3) b wt BoHV-5 (3) b | 35 dpi, 4×107 PFU | 28 dpc | |
| Negative control | 3 | 28 dpc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marawan, M.A.; Deng, M.; Wang, C.; Chen, Y.; Hu, C.; Chen, J.; Chen, X.; Chen, H.; Guo, A. Characterization of BoHV-1 gG-/tk-/gE- Mutant in Differential Protein Expression, Virulence, and Immunity. Vet. Sci. 2021, 8, 253. https://doi.org/10.3390/vetsci8110253
Marawan MA, Deng M, Wang C, Chen Y, Hu C, Chen J, Chen X, Chen H, Guo A. Characterization of BoHV-1 gG-/tk-/gE- Mutant in Differential Protein Expression, Virulence, and Immunity. Veterinary Sciences. 2021; 8(11):253. https://doi.org/10.3390/vetsci8110253
Chicago/Turabian StyleMarawan, Marawan A., Mingliang Deng, Chen Wang, Yingyu Chen, Changmin Hu, Jianguo Chen, Xi Chen, Huanchun Chen, and Aizhen Guo. 2021. "Characterization of BoHV-1 gG-/tk-/gE- Mutant in Differential Protein Expression, Virulence, and Immunity" Veterinary Sciences 8, no. 11: 253. https://doi.org/10.3390/vetsci8110253
APA StyleMarawan, M. A., Deng, M., Wang, C., Chen, Y., Hu, C., Chen, J., Chen, X., Chen, H., & Guo, A. (2021). Characterization of BoHV-1 gG-/tk-/gE- Mutant in Differential Protein Expression, Virulence, and Immunity. Veterinary Sciences, 8(11), 253. https://doi.org/10.3390/vetsci8110253