
Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
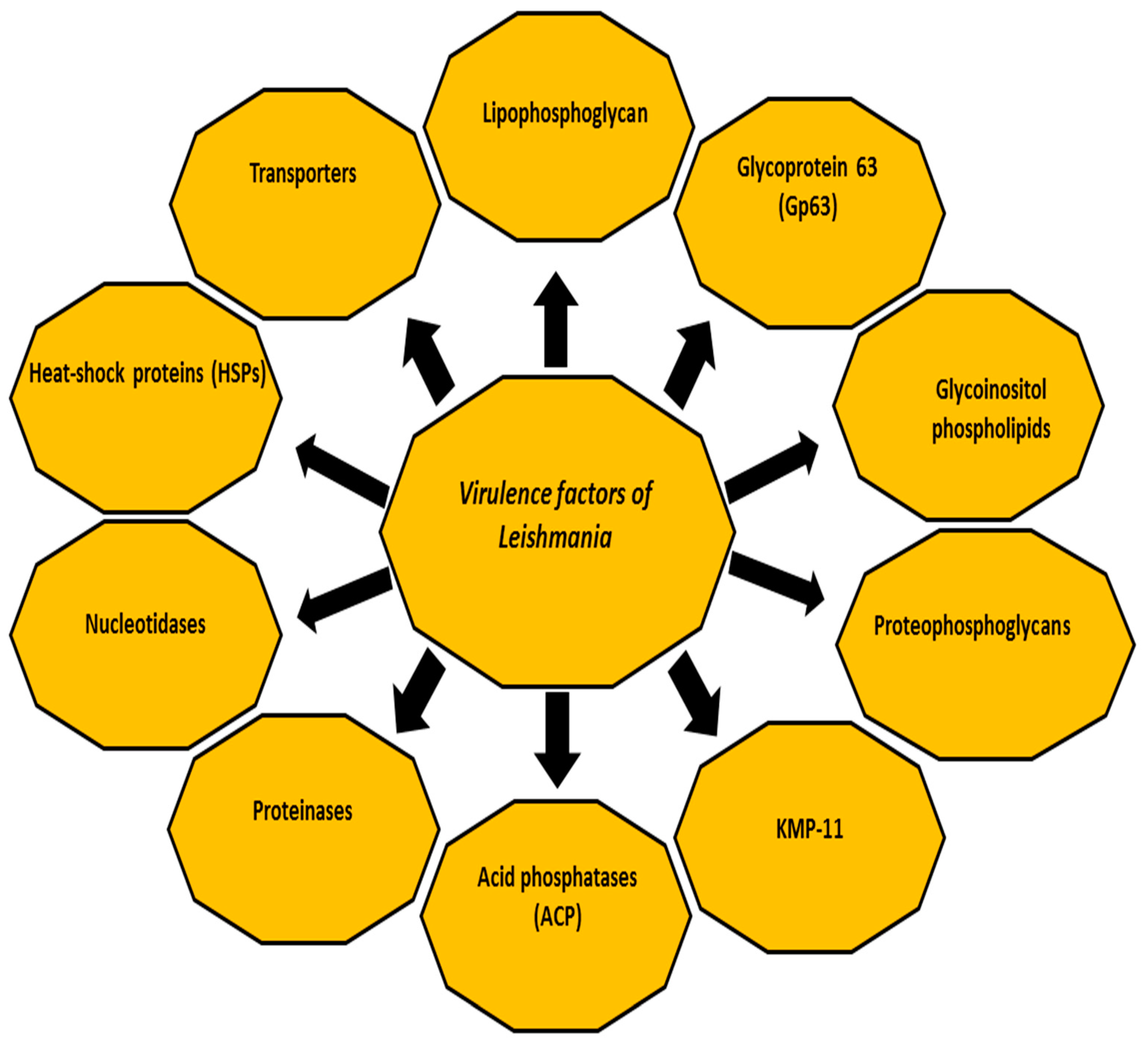
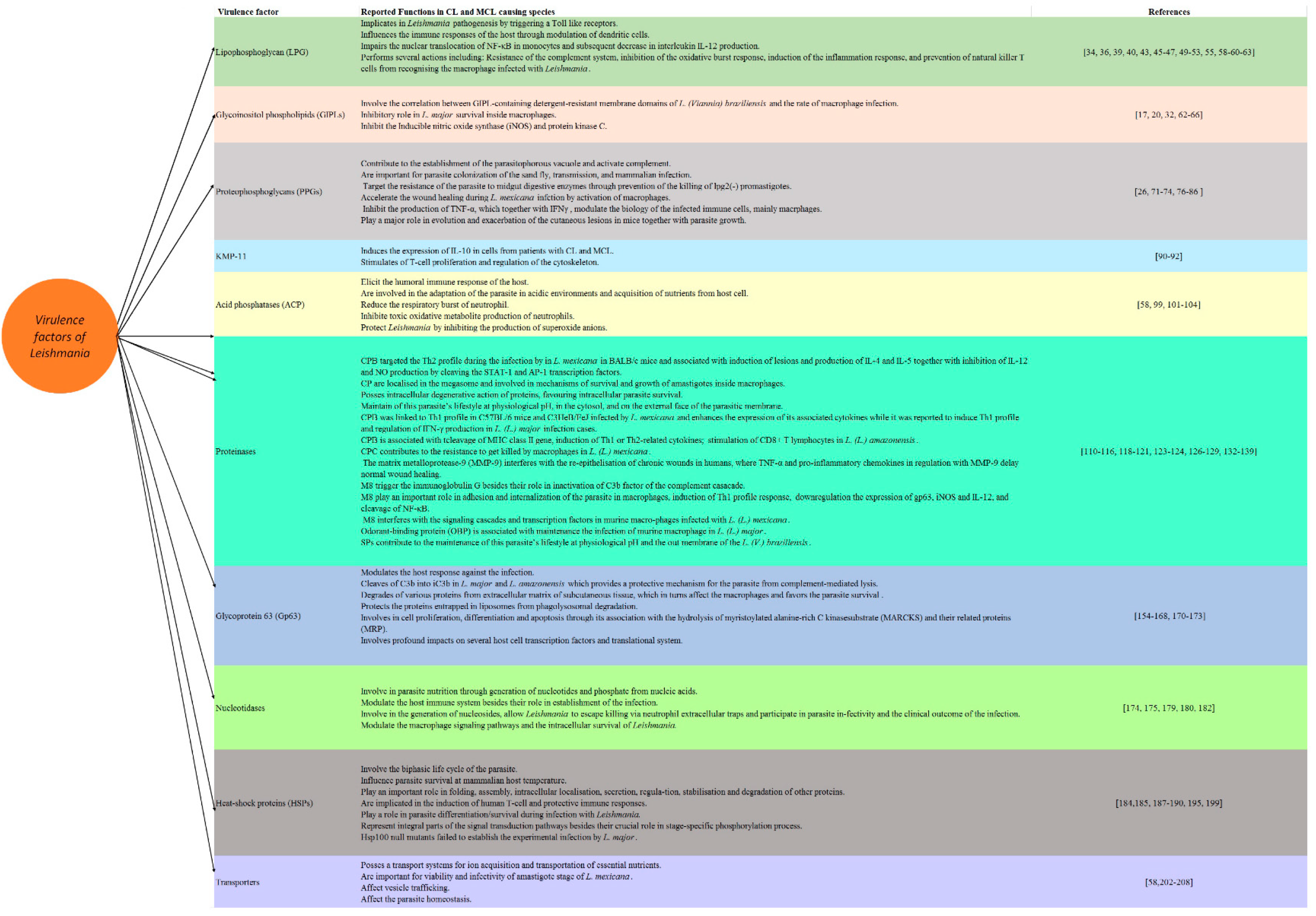
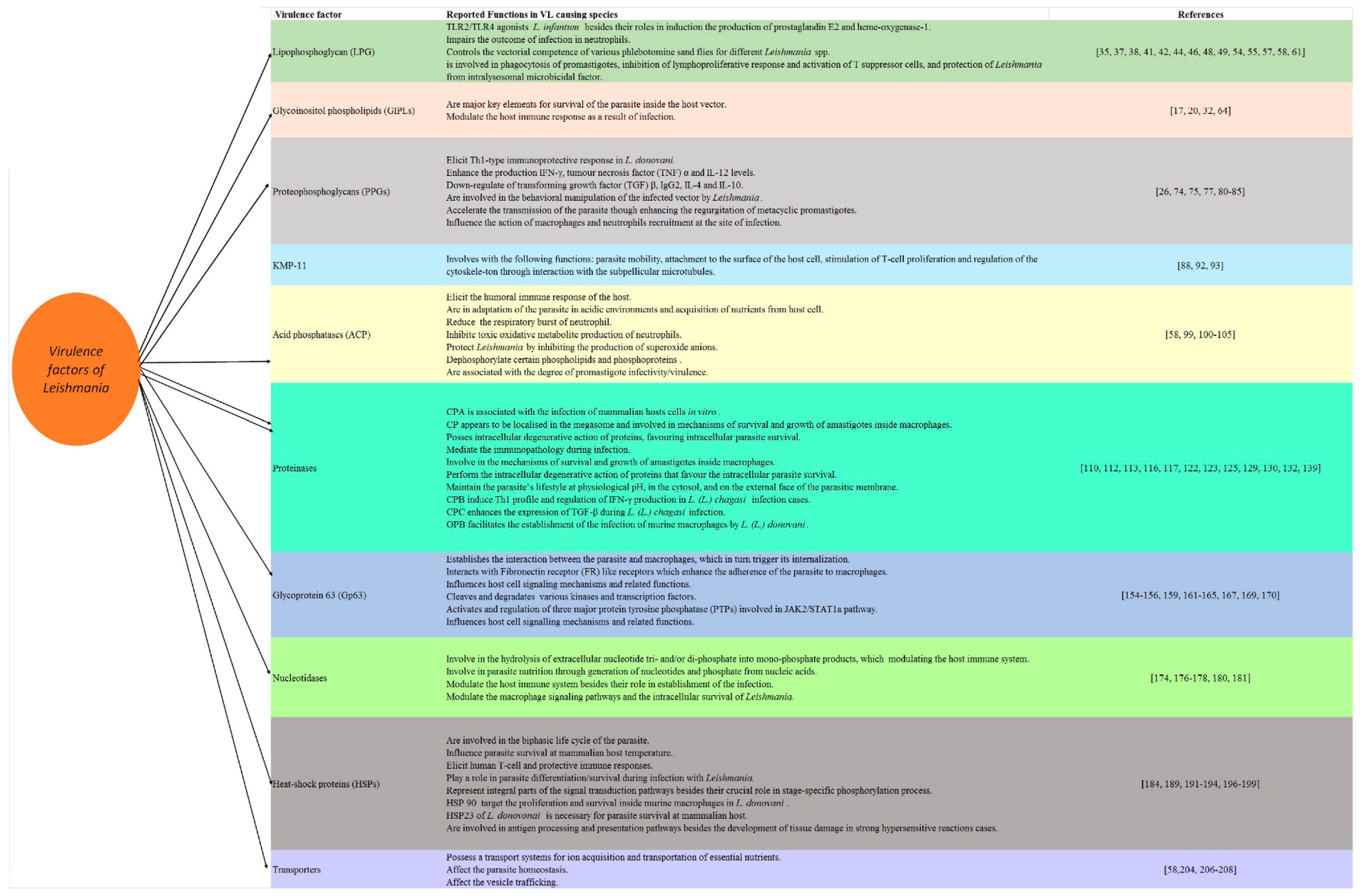
2. Virulence Factors of Leishmania
2.1. Lipophosphoglycan
2.2. Glycoinositolphospholipids (GIPLs)
2.3. Proteophosphoglycans (PPGs)
2.4. 11 kDa Kinetoplastid Membrane Protein (KMP-11)
2.5. Acid Phosphatases (ACPs)
2.6. Proteinases
2.7. Nucleotidases
2.8. Heat-Shock Proteins (HSPs)
2.9. Transporters
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clem, A. A current perspective on leishmaniasis. J. Glob. Infect. Dis. 2010, 2, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Romero, H.D.; Lde, A.S.; Silva-Vergara, M.L.; Rodrigues, V.; Costa, R.T.; Guimaraes, S.F.; Alecrim, W.; Moraes-Souza, H.; Prata, A. Comparative study of serologic tests for the diagnosis of asymptomatic visceral leishmaniasis in an endemic area. Am. J. Trop. Med. Hyg. 2009, 81, 27–33. [Google Scholar] [CrossRef] [PubMed]
- WHO. Control of the leishmaniases. World Health Organ. Tech. Rep. Ser. 2010, xii–xiii, 1–186. [Google Scholar]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Barrett, M.P.; Croft, S.L. Management of trypanosomiasis and leishmaniasis. Br. Med. Bull. 2012, 104, 175–196. [Google Scholar] [CrossRef] [Green Version]
- Elmahallawy, E.K.; Agil, A. Treatment of leishmaniasis: A review and assessment of recent research. Curr. Pharm. Dis. 2015, 21, 2259–2275. [Google Scholar] [CrossRef] [PubMed]
- Elmahallawy, E.K.; Martinez, A.S.; Rodriguez-Granger, J.; Hoyos-Mallecot, Y.; Agil, A.; Mari, J.M.N.; Fernandez, J.G. Diagnosis of leishmaniasis. J. Infect. Dev. Ctries. 2014, 8, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L. Leishmaniasis. Lancet 1999, 354, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Desjeux, P. Human leishmaniases: Epidemiology and public health aspects. World Health Stat. Q. Rapp. Trimest. Stat. Sanit. Mond. 1992, 45, 267–275. [Google Scholar]
- Desjeux, P. Leishmaniasis: Current situation and new perspectives. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Bray, R.C. Human Parasitic Diseases: Leishmaniasis; Elsevier Science Publishers: Oxford, UK, 1985. [Google Scholar]
- Chappuis, F.; Sundar, S.; Hailu, A.; Ghalib, H.; Rijal, S.; Peeling, R.W.; Alvar, J.; Boelaert, M. Visceral leishmaniasis: What are the needs for diagnosis, treatment and control? Nat. Rev. Microbiol 2007, 5, 873–882. [Google Scholar] [CrossRef]
- Okwor, I.; Uzonna, J. Persistent parasites and immunologic memory in cutaneous leishmaniasis: Implications for vaccine designs and vaccination strategies. Immunol. Res. 2008, 41, 123–136. [Google Scholar] [CrossRef]
- Vanloubbeeck, Y.; Jones, D.E. The immunology of Leishmania infection and the implications for vaccine development. Ann. N. Y. Acad. Sci. 2004, 1026, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Contreras, I.; Gomez, M.A.; Nguyen, O.; Shio, M.T.; McMaster, R.W.; Olivier, M. Leishmania-induced inactivation of the macrophage transcription factor AP-1 is mediated by the parasite metalloprotease GP63. PLoS Pathog. 2010, 6, e1001148. [Google Scholar] [CrossRef] [Green Version]
- Shio, M.T.; Hassani, K.; Isnard, A.; Ralph, B.; Contreras, I.; Gomez, M.A.; Abu-Dayyeh, I.; Olivier, M. Host cell signalling and leishmania mechanisms of evasion. J. Trop. Med. 2012, 2012, 819512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandonisio, O.; Panaro, M.A.; Sisto, M.; Acquafredda, A.; Fumarola, L.; Leogrande, D. Interactions between Leishmania parasites and host cells. Parassitologia 2000, 42, 183–190. [Google Scholar] [PubMed]
- Olivier, M.; Atayde, V.D.; Isnard, A.; Hassani, K.; Shio, M.T. Leishmania virulence factors: Focus on the metalloprotease GP63. Microbes Infect. 2012, 14, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Naderer, T.; Vince, J.E.; McConville, M.J. Surface determinants of Leishmania parasites and their role in infectivity in the mammalian host. Curr. Mol. Med. 2004, 4, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Silva-Almeida, M.; Pereira, B.A.; Ribeiro-Guimaraes, M.L.; Alves, C.R. Proteinases as virulence factors in Leishmania spp. infection in mammals. Parasit Vectors 2012, 5, 160. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, L.U. Leishmania mexicana infection induces IgG to parasite surface glycoinositol phospholipids that can induce IL-10 in mice and humans. PLoS Negl. Trop. Dis. 2013, 7, e2224. [Google Scholar] [CrossRef] [Green Version]
- Zufferey, R.; Allen, S.; Barron, T.; Sullivan, D.R.; Denny, P.W.; Almeida, I.C.; Smith, D.F.; Turco, S.J.; Ferguson, M.A.; Beverley, S.M. Ether phospholipids and glycosylinositolphospholipids are not required for amastigote virulence or for inhibition of macrophage activation by Leishmania major. J. Biol. Chem. 2003, 278, 44708–44718. [Google Scholar] [CrossRef] [Green Version]
- Matlashewski, G. Leishmania infection and virulence. Med. Microbiol. Immunol. 2001, 190, 37–42. [Google Scholar] [CrossRef]
- Dostalova, A.; Volf, P. Leishmania development in sand flies: Parasite-vector interactions overview. Parasites Vectors 2012, 5, 276. [Google Scholar] [CrossRef] [Green Version]
- Spath, G.F.; Epstein, L.; Leader, B.; Singer, S.M.; Avila, H.A.; Turco, S.J.; Beverley, S.M. Lipophosphoglycan is a virulence factor distinct from related glycoconjugates in the protozoan parasite Leishmania major. Proc. Natl. Acad. Sci. USA 2000, 97, 9258–9263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M.E. The role of leishmania proteophosphoglycans in sand fly transmission and infection of the Mammalian host. Front. Microbiol. 2012, 3, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.P.; Figueiredo, F.B.; Morgado, F.N.; Renzetti, A.R.d.S.; de Souza, S.M.M.; Pereira, S.A.; Rodrigues-Da-Silva, R.N.; Lima-Junior, J.D.C.; De Luca, P.M. Leishmania infantum Virulence Factor A2 Protein: Linear B-Cell Epitope Mapping and Identification of Three Main Linear B-Cell Epitopes in Vaccinated and Naturally Infected Dogs. Front. Immunol. 2018, 9, 1690. [Google Scholar] [CrossRef] [PubMed]
- Guha, R.; Das, S.; Ghosh, J.; Naskar, K.; Mandala, A.; Sundar, S.; Dujardin, J.C.; Roy, S. Heterologous priming-boosting with DNA and vaccinia virus expressing kinetoplastid membrane protein-11 induces potent cellular immune response and confers protection against infection with antimony resistant and sensitive strains of Leishmania (Leishmania) donovani. Vaccine 2013, 31, 1905–1915. [Google Scholar] [CrossRef]
- Proudfoot, L.; O’Donnell, C.A.; Liew, F.Y. Glycoinositolphospholipids of Leishmania major inhibit nitric oxide synthesis and reduce leishmanicidal activity in murine macrophages. Eur. J. Immunol. 1995, 25, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Reed, S.G.; McGwire, B.S.; Soong, L. Leishmania model for microbial virulence: The relevance of parasite multiplication and pathoantigenicity. Acta Trop. 2003, 85, 375–390. [Google Scholar] [CrossRef]
- Chang, K.P.; McGwire, B.S. Molecular determinants and regulation of Leishmania virulence. Kinetoplastid Biol. Dis. 2002, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- da Pires, S.F.; Fialho, L.C., Jr.; Silva, S.O.; Melo, M.N.; de Souza, C.C.; Tafuri, W.L.; Romero, O.B.; de Andrade, H.M. Identification of virulence factors in Leishmania infantum strains by a proteomic approach. J. Proteome Res. 2014, 13, 1860–1872. [Google Scholar] [CrossRef]
- Forestier, C.L.; Gao, Q.; Boons, G.J. Leishmania lipophosphoglycan: How to establish structure-activity relationships for this highly complex and multifunctional glycoconjugate? Front. Cell. Infect. Microbiol. 2014, 4, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodge, R.; Descoteaux, A. Leishmania donovani promastigotes induce periphagosomal F-actin accumulation through retention of the GTPase Cdc42. Cell Microbiol. 2005, 7, 1647–1658. [Google Scholar] [CrossRef]
- de Assis, R.R.; Ibraim, I.C.; Nogueira, P.M.; Soares, R.P.; Turco, S.J. Glycoconjugates in New World species of Leishmania: Polymorphisms in lipophosphoglycan and glycoinositolphospholipids and interaction with hosts. Biochim. Biophys. Acta 2012, 1820, 1354–1365. [Google Scholar] [CrossRef]
- Sacks, D.L.; Pimenta, P.; McConville, M.J.; Schneider, P.; Turco, S.J. Stage-specific binding of Leishmania donovani to the sand fly vector midgut is regulated by conformational changes in the abundant surface lipophosphoglycan. J. Exp. Med. 1995, 181, 685–697. [Google Scholar] [CrossRef] [Green Version]
- Sacks, D.L.; Brodin, T.N.; Turco, S.J. Developmental modification of the lipophosphoglycan from Leishmania major promastigotes during metacyclogenesis. Mol. Biochem. Parasitol. 1990, 42, 225–233. [Google Scholar] [CrossRef]
- Soares, R.P.; Macedo, M.E.; Ropert, C.; Gontijo, N.F.; Almeida, I.C.; Gazzinelli, R.T.; Pimenta, P.F.; Turco, S.J. Leishmania chagasi: Lipophosphoglycan characterization and binding to the midgut of the sand fly vector Lutzomyia longipalpis. Mol. Biochem. Parasitol. 2002, 121, 213–224. [Google Scholar] [CrossRef]
- Coelho-Finamore, J.; Freitas, V.; Assis, R.; Melo, M.; Novozhilova, N.; Secundino, N.; Pimenta, P.; Turco, S.; Soares, R. Leishmania infantum: Lipophosphoglycan intraspecific variation and interaction with vertebrate and invertebrate hosts. Int. J. Parasitol. 2011, 41, 333–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volf, P.; Nogueira, P.M.; Myskova, J.; Turco, S.J.; Soares, R.P. Structural comparison of lipophosphoglycan from Leishmania turanica and L. major, two species transmitted by Phlebotomus papatasi. Parasitol. Int. 2014, 63, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Dobson, D.E.; Scholtes, L.D.; Myler, P.J.; Turco, S.J.; Beverley, S.M. Genomic organization and expression of the expanded SCG/L/R gene family of Leishmania major: Internal clusters and telomeric localization of SCGs mediating species-specific LPG modifications. Mol. Biochem. Parasitol. 2006, 146, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.A.; Araujo, G.V.; Sandoval, C.M.; Nogueira, P.M.; Zúniga, C.; Sosa-Ochoa, W.H.; Laurenti, M.D.; Soares, R.P. Lipophosphoglycans from dermotropic Leishmania infantum are more pro-inflammatory than those from viscerotropic strains. Memórias Instituto Oswaldo Cruz 2020, 115, 115. [Google Scholar] [CrossRef] [PubMed]
- Ibraim, I.C.; de Assis, R.R.; Pessoa, N.L.; Campos, M.A.; Melo, M.N.; Turco, S.J.; Soares, R.P. Two biochemically distinct lipophosphoglycans from Leishmania braziliensis and Leishmania infantum trigger different innate immune responses in murine macrophages. Parasites Vectors 2013, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, P.M.; Assis, R.R.; Torrecilhas, A.C.; Saraiva, E.M.; Pessoa, N.L.; Campos, M.A.; Marialva, E.F.; Ríos-Velasquez, C.M.; Pessoa, F.A.; Secundino, N.F. Lipophosphoglycans from Leishmania amazonensis strains display immunomodulatory properties via TLR4 and do not affect sand fly infection. PLoS Negl. Trop. Dis. 2016, 10, e0004848. [Google Scholar] [CrossRef]
- McConville, M.J.; Blackwell, J.M. Developmental changes in the glycosylated phosphatidylinositols of Leishmania donovani. Characterization of the promastigote and amastigote glycolipids. J. Biol. Chem. 1991, 266, 15170–15179. [Google Scholar] [CrossRef]
- Bahr, V.; Stierhof, Y.D.; Ilg, T.; Demar, M.; Quinten, M.; Overath, P. Expression of lipophosphoglycan, high-molecular weight phosphoglycan and glycoprotein 63 in promastigotes and amastigotes of Leishmania mexicana. Mol. Biochem. Parasitol. 1993, 58, 107–121. [Google Scholar] [CrossRef]
- Amer, A.O.; Swanson, M.S. A phagosome of one’s own: A microbial guide to life in the macrophage. Curr. Opin. Microbiol. 2002, 5, 56–61. [Google Scholar] [CrossRef]
- Spath, G.F.; Garraway, L.A.; Turco, S.J.; Beverley, S.M. The role(s) of lipophosphoglycan (LPG) in the establishment of Leishmania major infections in mammalian hosts. Proc. Natl. Acad. Sci. USA 2003, 100, 9536–9541. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.B.; Araujo-Santos, T.; Lazaro-Souza, M.; Carneiro, A.B.; Ibraim, I.C.; Jesus-Santos, F.H.; Luz, N.F.; Pontes, S.M.; Entringer, P.F.; Descoteaux, A.; et al. Leishmania infantum lipophosphoglycan induced-Prostaglandin E2 production in association with PPAR-gamma expression via activation of Toll like receptors-1 and 2. Sci. Rep. 2017, 7, 14321. [Google Scholar] [CrossRef] [PubMed]
- Handman, E.; McConville, M.J.; Goding, J.W. Carbohydrate antigens as possible parasite vaccines A case for the Leishmania glycolipid. Immunol. Today 1987, 8, 181–185. [Google Scholar] [CrossRef]
- Svarovska, A.; Ant, T.H.; Seblova, V.; Jecna, L.; Beverley, S.M.; Volf, P. Leishmania major glycosylation mutants require phosphoglycans (lpg2-) but not lipophosphoglycan (lpg1-) for survival in permissive sand fly vectors. PLoS Negl. Trop. Dis. 2010, 4, e580. [Google Scholar] [CrossRef] [PubMed]
- Argueta-Donohue, J.; Carrillo, N.; Valdes-Reyes, L.; Zentella, A.; Aguirre-Garcia, M.; Becker, I.; Gutierrez-Kobeh, L. Leishmania mexicana: Participation of NF-kappaB in the differential production of IL-12 in dendritic cells and monocytes induced by lipophosphoglycan (LPG). Exp. Parasitol. 2008, 120, 1–9. [Google Scholar] [CrossRef]
- Von Stebut, E. Immunology of cutaneous leishmaniasis: The role of mast cells, phagocytes and dendritic cells for protective immunity. Eur. J. Dermatol. 2007, 17, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Carrada, G.; Caneda, C.; Salaiza, N.; Delgado, J.; Ruiz, A.; Sanchez, B.; Gutierrez-Kobeh, L.; Aguirre, M.; Becker, I. Monocyte cytokine and costimulatory molecule expression in patients infected with Leishmania mexicana. Parasite Immunol. 2007, 29, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Luz, N.F.; Andrade, B.B.; Feijó, D.F.; Araújo-Santos, T.; Carvalho, G.Q.; Andrade, D.; Abánades, D.R.; Melo, E.V.; Silva, A.M.; Brodskyn, C.I. Heme oxygenase-1 promotes the persistence of Leishmania chagasi infection. J. Immunol. 2012, 188, 4460–4467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Kebaier, C.; Pakpour, N.; Capul, A.A.; Beverley, S.M.; Scott, P.; Uzonna, J.E. Leishmania major phosphoglycans influence the host early immune response by modulating dendritic cell functions. Infect. Immun. 2009, 77, 3272–3283. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, P.F.; Saraiva, E.M.; Rowton, E.; Modi, G.B.; Garraway, L.A.; Beverley, S.M.; Turco, S.J.; Sacks, D.L. Evidence that the vectorial competence of phlebotomine sand flies for different species of Leishmania is controlled by structural polymorphisms in the surface lipophosphoglycan. Proc. Natl. Acad. Sci. USA 1994, 91, 9155–9159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesus-Santos, F.H.; Lobo-Silva, J.; Ramos, P.I.P.; Descoteaux, A.; Lima, J.B.; Borges, V.M.; Farias, L.P. LPG2 gene duplication in Leishmania infantum: A case for CRISPR-Cas9 gene editing. Front. Cell. Infect. Microbiol. 2020, 10, 408. [Google Scholar] [CrossRef]
- Chang, K.P.; Chaudhuri, G.; Fong, D. Molecular determinants of Leishmania virulence. Annu. Rev. Microbiol. 1990, 44, 499–529. [Google Scholar] [CrossRef]
- Da Silva, R.P.; Hall, B.F.; Joiner, K.A.; Sacks, D.L. CR1, the C3b receptor, mediates binding of infective Leishmania major metacyclic promastigotes to human macrophages. J. Immunol. 1989, 143, 617–622. [Google Scholar]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef] [Green Version]
- McConville, M.J.; Bacic, A. A family of glycoinositol phospholipids from Leishmania major. Isolation, characterization, and antigenicity. J. Biol. Chem. 1989, 264, 757–766. [Google Scholar] [CrossRef]
- Yoneyama, K.A.; Tanaka, A.K.; Silveira, T.G.; Takahashi, H.K.; Straus, A.H. Characterization of Leishmania (Viannia) braziliensis membrane microdomains, and their role in macrophage infectivity. J. Lipid Res. 2006, 47, 2171–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broermann, L.; Heidenreich, W. Malaria tropica and pregnancy. Geburtshilfe Und Frauenheilkd 1992, 52, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Passero, L.F.; Assis, R.R.; da Silva, T.N.; Nogueira, P.M.; Macedo, D.H.; Pessoa, N.L.; Campos, M.A.; Laurenti, M.D.; Soares, R.P. Differential modulation of macrophage response elicited by glycoinositolphospholipids and lipophosphoglycan from Leishmania (Viannia) shawi. Parasitol. Int. 2015, 64, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Paranaíba, L.F.; de Assis, R.R.; Nogueira, P.M.; Torrecilhas, A.C.; Campos, J.H.; de Silveira, A.C.O.; Martins-Filho, O.A.; Pessoa, N.L.; Campos, M.A.; Parreiras, P.M. Leishmania enriettii: Biochemical characterisation of lipophosphoglycans (LPGs) and glycoinositolphospholipids (GIPLs) and infectivity to Cavia porcellus. Parasites Vectors 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilg, T.; Montgomery, J.; Stierhof, Y.D.; Handman, E. Molecular cloning and characterization of a novel repeat-containing Leishmania major gene, ppg1, that encodes a membrane-associated form of proteophosphoglycan with a putative glycosylphosphatidylinositol anchor. J. Biol. Chem. 1999, 274, 31410–31420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilg, T. Proteophosphoglycans of Leishmania. Parasitol. Today 2000, 16, 489–497. [Google Scholar] [CrossRef]
- Klein, C.; Gopfert, U.; Goehring, N.; Stierhof, Y.D.; Ilg, T. Proteophosphoglycans of Leishmania mexicana. Identification, purification, structural and ultrastructural characterization of the secreted promastigote proteophosphoglycan pPPG2, a stage-specific glycoisoform of amastigote aPPG. Biochem. J. 1999, 344 Pt 3, 775–786. [Google Scholar] [CrossRef]
- Piani, A.; Ilg, T.; Elefanty, A.G.; Curtis, J.; Handman, E. Leishmania major proteophosphoglycan is expressed by amastigotes and has an immunomodulatory effect on macrophage function. Microbes Infect. 1999, 1, 589–599. [Google Scholar] [CrossRef]
- Aebischer, T.; Harbecke, D.; Ilg, T. Proteophosphoglycan, a major secreted product of intracellular Leishmania mexicana amastigotes, is a poor B-cell antigen and does not elicit a specific conventional CD4+ T-cell response. Infect. Immun. 1999, 67, 5379–5385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, C.; Kawakami, M.; Kaul, M.; Ilg, T.; Overath, P.; Aebischer, T. Secreted proteophosphoglycan of Leishmania mexicana amastigotes activates complement by triggering the mannan binding lectin pathway. Eur. J. Immunol. 1997, 27, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.; Stierhof, Y.D.; Ilg, T. Proteophosphoglycan secreted by Leishmania mexicana amastigotes causes vacuole formation in macrophages. Infect. Immun. 1997, 65, 783–786. [Google Scholar] [CrossRef] [Green Version]
- Duque, G.A.; Jardim, A.; Gagnon, E.; Fukuda, M.; Descoteaux, A. The host cell secretory pathway mediates the export of Leishmania virulence factors out of the parasitophorous vacuole. PLoS Pathog. 2019, 15, e1007982. [Google Scholar] [CrossRef] [Green Version]
- Samant, M.; Gupta, R.; Kumari, S.; Misra, P.; Khare, P.; Kushawaha, P.K.; Sahasrabuddhe, A.A.; Dube, A. Immunization with the DNA-encoding N-terminal domain of proteophosphoglycan of Leishmania donovani generates Th1-type immunoprotective response against experimental visceral leishmaniasis. J. Immunol 2009, 183, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M.E.; Ilg, T.; Nikolaev, A.V.; Ferguson, M.A.; Bates, P.A. Transmission of cutaneous leishmaniasis by sand flies is enhanced by regurgitation of fPPG. Nature 2004, 430, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M.E.; Bates, P.A. Leishmania manipulation of sand fly feeding behavior results in enhanced transmission. PLoS Pathog. 2007, 3, e91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secundino, N.; Kimblin, N.; Peters, N.C.; Lawyer, P.; Capul, A.A.; Beverley, S.M.; Turco, S.J.; Sacks, D. Proteophosphoglycan confers resistance of Leishmania major to midgut digestive enzymes induced by blood feeding in vector sand flies. Cell. Microbiol. 2010, 12, 906–918. [Google Scholar] [CrossRef] [Green Version]
- Giraud, E.; Lestinova, T.; Derrick, T.; Martin, O.; Dillon, R.J.; Volf, P.; Műller, I.; Bates, P.A.; Rogers, M.E. Leishmania proteophosphoglycans regurgitated from infected sand flies accelerate dermal wound repair and exacerbate leishmaniasis via insulin-like growth factor 1-dependent signalling. PLoS Pathog. 2018, 14, e1006794. [Google Scholar] [CrossRef] [Green Version]
- Stierhof, Y.-D.; Bates, P.A.; Jacobson, R.L.; Rogers, M.E.; Schlein, Y.; Handman, E.; Ilg, T. Filamentous proteophosphoglycan secreted by Leishmania promastigotes forms gel-like three-dimensional networks that obstruct the digestive tract of infected sandfly vectors. Eur. J. Cell Biol. 1999, 78, 675–689. [Google Scholar] [CrossRef]
- Rogers, M.; Kropf, P.; Choi, B.-S.; Dillon, R.; Podinovskaia, M.; Bates, P.; Müller, I. Proteophosophoglycans regurgitated by Leishmania-infected sand flies target the L-arginine metabolism of host macrophages to promote parasite survival. PLoS Pathog. 2009, 5, e1000555. [Google Scholar] [CrossRef] [Green Version]
- Sacks, D.; Kamhawi, S. Molecular aspects of parasite-vector and vector-host interactions in leishmaniasis. Annu. Rev. Microbiol. 2001, 55, 453–483. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.C.; Egen, J.G.; Secundino, N.; Debrabant, A.; Kimblin, N.; Kamhawi, S.; Lawyer, P.; Fay, M.P.; Germain, R.N.; Sacks, D. In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies. Science 2008, 321, 970–974. [Google Scholar] [CrossRef] [Green Version]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Kropf, P.; Fuentes, J.M.; Fähnrich, E.; Arpa, L.; Herath, S.; Weber, V.; Soler, G.; Celada, A.; Modolell, M.; Müller, I. Arginase and polyamine synthesis are key factors in the regulation of experimental leishmaniasis in vivo. FASEB J. 2005, 19, 1000–1002. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Svobodová, M.; Müller, I.; Volf, P.; Rogers, M. Promastigote secretory gel from natural and unnatural sand fly vectors exacerbate Leishmania major and Leishmania tropica cutaneous leishmaniasis in mice. Parasitology 2019, 146, 1796–1802. [Google Scholar] [CrossRef] [Green Version]
- Jardim, A.; Funk, V.; Caprioli, R.M.; Olafson, R.W. Isolation and structural characterization of the Leishmania donovani kinetoplastid membrane protein-11, a major immunoreactive membrane glycoprotein. Biochem. J. 1995, 305 Pt 1, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Tolson, D.L.; Jardim, A.; Schnur, L.F.; Stebeck, C.; Tuckey, C.; Beecroft, R.P.; Teh, H.S.; Olafson, R.W.; Pearson, T.W. The kinetoplastid membrane protein 11 of Leishmania donovani and African trypanosomes is a potent stimulator of T-lymphocyte proliferation. Infect. Immun. 1994, 62, 4893–4899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, S.F. Molecular variation in Leishmania. Acta Trop. 1993, 53, 185–204. [Google Scholar] [CrossRef]
- Matos, D.C.; Faccioli, L.A.; Cysne-Finkelstein, L.; Luca, P.M.; Corte-Real, S.; Armoa, G.R.; Lemes, E.M.; Decote-Ricardo, D.; Mendonca, S.C. Kinetoplastid membrane protein-11 is present in promastigotes and amastigotes of Leishmania amazonensis and its surface expression increases during metacyclogenesis. Memórias Instituto Oswaldo Cruz 2010, 105, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.P.; Passos, S.; Dutra, W.O.; Soto, M.; Alonso, C.; Gollob, K.J.; Carvalho, E.M.; de Jesus, A.R. Effect of LACK and KMP11 on IFN-gamma production by peripheral blood mononuclear cells from cutaneous and mucosal leishmaniasis patients. Scand. J. Immunol. 2005, 61, 337–342. [Google Scholar] [CrossRef]
- Skeiky, Y.A.; Guderian, J.A.; Benson, D.R.; Bacelar, O.; Carvalho, E.M.; Kubin, M.; Badaro, R.; Trinchieri, G.; Reed, S.G. A recombinant Leishmania antigen that stimulates human peripheral blood mononuclear cells to express a Th1-type cytokine profile and to produce interleukin 12. J. Exp. Med. 1995, 181, 1527–1537. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Sen, P.; Majumder, H.K.; Roy, S. Reduced expression of lipophosphoglycan (LPG) and kinetoplastid membrane protein (KMP)-11 in Leishmania donovani promastigotes in axenic culture. J. Parasitol. 1998, 84, 644–647. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, Y.; Tal, E.; Sulman, F.G. Heat-induced migraine and its treatment. Harefuah 1978, 95, 158–161. [Google Scholar]
- Bates, P.A.; Dwyer, D.M. Biosynthesis and secretion of acid phosphatase by Leishmania donovani promastigotes. Mol. Biochem. Parasitol. 1987, 26, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Freitas-Mesquita, A.L.; Fonseca-de-Souza, A.L.; Meyer-Fernandes, J.R. Leishmania amazonensis: Characterization of an ecto-pyrophosphatase activity. Exp. Parasitol. 2014, 137, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Ilg, T.; Stierhof, Y.D.; Etges, R.; Adrian, M.; Harbecke, D.; Overath, P. Secreted acid phosphatase of Leishmania mexicana: A filamentous phosphoglycoprotein polymer. Proc. Natl. Acad. Sci. USA 1991, 88, 8774–8778. [Google Scholar] [CrossRef] [Green Version]
- Stierhof, Y.D.; Wiese, M.; Ilg, T.; Overath, P.; Haner, M.; Aebi, U. Structure of a filamentous phosphoglycoprotein polymer: The secreted acid phosphatase of Leishmania mexicana. J. Mol. Biol. 1998, 282, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Glew, R.H.; Saha, A.K.; Das, S.; Remaley, A.T. Biochemistry of the Leishmania species. Microbiol. Rev. 1988, 52, 412–432. [Google Scholar] [CrossRef]
- Remaley, A.T.; Glew, R.H.; Kuhns, D.B.; Basford, R.E.; Waggoner, A.S.; Ernst, L.A.; Pope, M. Leishmania donovani: Surface membrane acid phosphatase blocks neutrophil oxidative metabolite production. Exp. Parasitol. 1985, 60, 331–341. [Google Scholar] [CrossRef]
- Kane, M.M.; Mosser, D.M. Leishmania parasites and their ploys to disrupt macrophage activation. Curr. Opin. Hematol. 2000, 7, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Stafford, J.L.; Neumann, N.F.; Belosevic, M. Macrophage-mediated innate host defense against protozoan parasites. Crit. Rev. Microbiol. 2002, 28, 187–248. [Google Scholar] [CrossRef]
- Saha, A.K.; Das, S.; Glew, R.H.; Gottlieb, M. Resistance of leishmanial phosphatases to inactivation by oxygen metabolites. J. Clin. Microbiol. 1985, 22, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, S.L.; Shakarian, A.M.; Dwyer, D.M. Leishmania: Amastigotes synthesize conserved secretory acid phosphatases during human infection. Exp. Parasitol. 1998, 89, 161–168. [Google Scholar] [CrossRef]
- Singla, N.; Khuller, G.K.; Vinayak, V.K. Acid phosphatase activity of promastigotes of Leishmania donovani: A marker of virulence. FEMS Microbiol. Lett. 1992, 73, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.J. Classification of peptidases. Methods Enzymol. 1994, 244, 1–15. [Google Scholar] [PubMed]
- McKerrow, J.H.; Caffrey, C.; Kelly, B.; Loke, P.N.; Sajid, M. Proteases in parasitic diseases. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 497–536. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Bateman, A. MEROPS: The peptidase database. Nucleic Acids Res. 2010, 38, D227–D233. [Google Scholar] [CrossRef]
- Rawlings, N.D. Peptidase inhibitors in the MEROPS database. Biochimie 2010, 92, 1463–1483. [Google Scholar] [CrossRef]
- North, M.J.; Mottram, J.C.; Coombs, G.H. Cysteine proteinases of parasitic protozoa. Parasitol. Today 1990, 6, 270–275. [Google Scholar] [CrossRef]
- Pral, E.M.; Bijovsky, A.T.; Balanco, J.M.; Alfieri, S.C. Leishmania mexicana: Proteinase activities and megasomes in axenically cultivated amastigote-like forms. Exp. Parasitol. 1993, 77, 62–73. [Google Scholar] [CrossRef]
- Das, P.; Alam, M.N.; Paik, D.; Karmakar, K.; De, T.; Chakraborti, T. Protease inhibitors in potential drug development for Leishmaniasis. Indian J. Biochem. Biophys. 2013, 50, 363–376. [Google Scholar] [PubMed]
- Sajid, M.; McKerrow, J.H. Cysteine proteases of parasitic organisms. Mol. Biochem. Parasitol. 2002, 120, 1–21. [Google Scholar] [CrossRef]
- Pupkis, M.F.; Tetley, L.; Coombs, G.H. Leishmania mexicana: Amastigote hydrolases in unusual lysosomes. Exp. Parasitol. 1986, 62, 29–39. [Google Scholar] [CrossRef]
- Rafati, S.; Couty-Jouve, S.; Alimohammadian, M.; Louis, J. Biochemical analysis and immunogenicity of Leishmania major amastigote fractions in cutaneous leishmaniasis. Clin. Exp. Immunol. 1997, 110, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Mottram, J.C.; Coombs, G.H.; Alexander, J. Cysteine peptidases as virulence factors of Leishmania. Curr. Opin. Microbiol. 2004, 7, 375–381. [Google Scholar] [CrossRef]
- Denise, H.; Poot, J.; Jiménez, M.; Ambit, A.; Herrmann, D.C.; Vermeulen, A.N.; Coombs, G.H.; Mottram, J.C. Studies on the CPA cysteine peptidase in the Leishmania infantum genome strain JPCM5. BMC Mol. Biol. 2006, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pollock, K.G.; McNeil, K.S.; Mottram, J.C.; Lyons, R.E.; Brewer, J.M.; Scott, P.; Coombs, G.H.; Alexander, J. The Leishmania mexicana cysteine protease, CPB2. 8, induces potent Th2 responses. J. Immunol. 2003, 170, 1746–1753. [Google Scholar] [CrossRef] [Green Version]
- Cameron, P.; McGachy, A.; Anderson, M.; Paul, A.; Coombs, G.H.; Mottram, J.C.; Alexander, J.; Plevin, R. Inhibition of lipopolysaccharide-induced macrophage IL-12 production by Leishmania mexicana amastigotes: The role of cysteine peptidases and the NF-κB signaling pathway. J. Immunol. 2004, 173, 3297–3304. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, L.U.; Denise, H.; Coombs, G.H.; Alexander, J.; Mottram, J.C.; Scott, P. Cysteine protease B of Leishmania mexicana inhibits host Th1 responses and protective immunity. J. Immunol. 2003, 171, 3711–3717. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.; Coombs, G.H.; Mottram, J.C. Leishmania mexicana cysteine proteinase-deficient mutants have attenuated virulence for mice and potentiate a Th1 response. J. Immunol. 1998, 161, 6794–6801. [Google Scholar] [PubMed]
- da Pinheiro, P.H.C.; de Dias, S.S.; Eulálio, K.D.; Mendonça, I.L.; Katz, S.; Barbiéri, C.L. Recombinant cysteine proteinase from Leishmania (Leishmania) chagasi implicated in human and dog T-cell responses. Infect. Immun. 2005, 73, 3787–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, C.; Benévolo-de-Andrade, T.; Alves, J.; Pirmez, C. Th1 and Th2 immunological profile induced by cysteine proteinase in murine leishmaniasis. Parasite Immunol. 2004, 26, 127–135. [Google Scholar] [CrossRef]
- Leao, S.D.S.; Lang, T.; Prina, E.; Hellio, R.; Antoine, J.-C. Intracellular Leishmania amazonensis amastigotes internalize and degrade MHC class II molecules of their host cells. J. Cell Sci. 1995, 108, 3219–3231. [Google Scholar]
- Rafati, S.; Fasel, N.; Masina, S. Leishmania cysteine proteinases: From gene to subunit vaccine. Curr. Genom. 2003, 4, 253–261. [Google Scholar] [CrossRef]
- Traub-Cseko, Y.M.; Duboise, M.; Boukai, L.K.; McMahon-Pratt, D. Identification of two distinct cysteine proteinase genes of Leishmania pifanoi axenic amastigotes using the polymerase chain reaction. Mol. Biochem. Parasitol. 1993, 57, 101–115. [Google Scholar] [CrossRef]
- Alves, C.R.; Côrte-Real, S.; De-Freitas, M.R.; Giovanni-De-Simone, S. Detection of cysteine-proteinases in Leishmania amazonensis promastigotes using a cross-reactive antiserum. FEMS Microbiol. Lett. 2000, 186, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Sakanari, J.A.; Nadler, S.A.; Chan, V.J.; Engel, J.C.; Leptak, C.; Bouvier, J. Leishmania major: Comparison of the cathepsin L-and B-like cysteine protease genes with those of other trypanosomatids. Exp. Parasitol. 1997, 85, 63–76. [Google Scholar] [CrossRef]
- Caffrey, C.R.; Lima, A.P.; Steverding, D. Cysteine peptidases of kinetoplastid parasites. Adv. Exp. Med. Biol. 2011, 712, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Somanna, A.; Mundodi, V.; Gedamu, L. Functional Analysis of Cathepsin B-like Cysteine Proteases fromLeishmania donovani Complex: Evidence for the activation of latent transforming growth factor β. J. Biol. Chem. 2002, 277, 25305–25312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frame, M.; Mottram, J.; Coombs, G. Analysis of the roles of cysteine proteinases of Leishmania mexicana in the host–parasite interaction. Parasitology 2000, 121, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Mottram, J.C.; Brooks, D.R.; Coombs, G.H. Roles of cysteine proteinases of trypanosomes and Leishmania in host-parasite interactions. Curr. Opin. Microbiol. 1998, 1, 455–460. [Google Scholar] [CrossRef]
- Alves, C.R.; Corte-Real, S.; Bourguignon, S.C.; Chaves, C.S.; Saraiva, E.M. Leishmania amazonensis: Early proteinase activities during promastigote-amastigote differentiation in vitro. Exp. Parasitol. 2005, 109, 38–48. [Google Scholar] [CrossRef]
- Santos-de-Souza, R.; de Castro Cortes, L.M.; Dos Charret, K.S.; Cysne-Finkelstein, L.; Alves, C.R.; Souza-Silva, F. Serine Proteinases in Leishmania (Viannia) braziliensis Promastigotes Have Distinct Subcellular Distributions and Expression. Int. J. Mol. Sci. 2019, 20, 1315. [Google Scholar] [CrossRef] [Green Version]
- Maretti-Mira, A.C.; de Oliveira-Neto, M.P.; Da-Cruz, A.M.; de Oliveira, M.P.; Craft, N.; Pirmez, C. Therapeutic failure in American cutaneous leishmaniasis is associated with gelatinase activity and cytokine expression. Clin. Exp. Immunol. 2011, 163, 207–214. [Google Scholar] [CrossRef]
- Reiss, M.J.; Han, Y.P.; Garcia, E.; Goldberg, M.; Yu, H.; Garner, W.L. Matrix metalloproteinase-9 delays wound healing in a murine wound model. Surgery 2010, 147, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Campos, T.M.; Passos, S.T.; Novais, F.O.; Beiting, D.P.; Costa, R.S.; Queiroz, A.; Mosser, D.; Scott, P.; Carvalho, E.M.; Carvalho, L.P. Matrix metalloproteinase 9 production by monocytes is enhanced by TNF and participates in the pathology of human cutaneous Leishmaniasis. PLoS Negl. Trop. Dis. 2014, 8, e3282. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.P.; Martins, V.T.; Santos, T.T.; Lage, D.P.; Ramos, F.F.; Salles, B.; Costa, L.E.; Dias, D.S.; Ribeiro, P.A.; Schneider, M.S. Small myristoylated protein-3, identified as a potential virulence factor in Leishmania amazonensis, proves to be a protective antigen against visceral leishmaniasis. Int. J. Mol. Sci. 2018, 19, 129. [Google Scholar] [CrossRef] [Green Version]
- Yao, C. Major surface protease of trypanosomatids: One size fits all? Infect. Immun. 2010, 78, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Thiakaki, M.; Kolli, B.; Chang, K.-P.; Soteriadou, K. Down-regulation of gp63 level in Leishmania amazonensis promastigotes reduces their infectivity in BALB/c mice. Microbes Infect. 2006, 8, 1455–1463. [Google Scholar] [CrossRef]
- Gregory, D.J.; Godbout, M.; Contreras, I.; Forget, G.; Olivier, M. A novel form of NF-κB is induced by Leishmania infection: Involvement in macrophage gene expression. Eur. J. Immunol. 2008, 38, 1071–1081. [Google Scholar] [CrossRef]
- Lieke, T.; Nylen, S.; Eidsmo, L.; McMaster, W.; Mohammadi, A.M.; Khamesipour, A.; Berg, L.; Akuffo, H. Leishmania surface protein gp63 binds directly to human natural killer cells and inhibits proliferation. Clin. Exp. Immunol. 2008, 153, 221–230. [Google Scholar] [CrossRef]
- Alves, C.R.; de Souza, R.S.; dos Charret, K.S.; de Cortes, L.M.C.; de Sa-Silva, M.P.; Barral-Veloso, L.; Oliveira, L.F.G.; da Silva, F.S. Understanding serine proteases implications on Leishmania spp lifecycle. Exp. Parasitol. 2018, 184, 67–81. [Google Scholar] [CrossRef]
- Swenerton, R.K.; Zhang, S.; Sajid, M.; Medzihradszky, K.F.; Craik, C.S.; Kelly, B.L.; McKerrow, J.H. The oligopeptidase B of Leishmania regulates parasite enolase and immune evasion. J. Biol. Chem. 2011, 286, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, R.O.; Pinto, E.F.; Lopes, J.R.C.; Guedes, H.L.M.; Fentanes, R.F.; Rossi-Bergmann, B. TGF-β-associated enhanced susceptibility to leishmaniasis following intramuscular vaccination of mice with Leishmania amazonensis antigens. Microbes Infect. 2005, 7, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.G.; Wilhelm, H. The involvement of the major surface glycoprotein (gp63) of Leishmania promastigotes in attachment to macrophages. J. Immunol. 1986, 136, 2613–2620. [Google Scholar] [PubMed]
- Bouvier, J.; Etges, R.; Bordier, C. Identification of the promastigote surface protease in seven species of Leishmania. Mol. Biochem. Parasitol. 1987, 24, 73–79. [Google Scholar] [CrossRef]
- Etges, R.; Bouvier, J.; Bordier, C. The major surface protein of Leishmania promastigotes is a protease. J. Biol. Chem. 1986, 261, 9098–9101. [Google Scholar] [CrossRef]
- Pandey, S.; Chakraborti, P.; Sharma, R.; Bandyopadhyay, S.; Sarkar, D.; Adhya, S. Involvement of Leishmania donovani major surface glycoprotein gp63 in promastigote multiplication. J. Biosci. 2004, 29, 15–22. [Google Scholar] [CrossRef]
- Hey, A.S.; Theander, T.G.; Hviid, L.; Hazrati, S.M.; Kemp, M.; Kharazmi, A. The major surface glycoprotein (gp63) from Leishmania major and Leishmania donovani cleaves CD4 molecules on human T cells. J. Immunol. 1994, 152, 4542–4548. [Google Scholar]
- Colomer-Gould, V.; Quintao, L.G.; Keithly, J.; Nogueira, N. A common major surface antigen on amastigotes and promastigotes of Leishmania species. J. Exp. Med. 1985, 162, 902–916. [Google Scholar] [CrossRef]
- Ellis, M.; Sharma, D.K.; Hilley, J.D.; Coombs, G.H.; Mottram, J.C. Processing and trafficking of Leishmania mexicana GP63. Analysis using GP18 mutants deficient in glycosylphosphatidylinositol protein anchoring. J. Biol. Chem. 2002, 277, 27968–27974. [Google Scholar] [CrossRef] [Green Version]
- Pimenta, P.F.; Saraiva, E.M.; Sacks, D.L. The comparative fine structure and surface glycoconjugate expression of three life stages of Leishmania major. Exp. Parasitol. 1991, 72, 191–204. [Google Scholar] [CrossRef]
- Brittingham, A.; Morrison, C.J.; McMaster, W.R.; McGwire, B.S.; Chang, K.-P.; Mosser, D.M. Role of the Leishmania surface protease gp63 in complement fixation, cell adhesion, and resistance to complement-mediated lysis. J. Immunol. 1995, 155, 3102–3111. [Google Scholar] [CrossRef]
- Brittingham, A.; Chen, G.; McGwire, B.S.; Chang, K.-P.; Mosser, D.M. Interaction of Leishmania gp63 with cellular receptors for fibronectin. Infect. Immun. 1999, 67, 4477–4484. [Google Scholar] [CrossRef] [Green Version]
- McGwire, B.S.; Chang, K.-P.; Engman, D.M. Migration through the extracellular matrix by the parasitic protozoan Leishmania is enhanced by surface metalloprotease gp63. Infect. Immun. 2003, 71, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, G.; Chaudhuri, M.; Pan, A.; Chang, K.P. Surface acid proteinase (gp63) of Leishmania mexicana: A metalloenzyme capable of protecting liposome-encapsulated proteins from phagolysosomal degradation by macrophages. J. Biol. Chem. 1989, 264, 7483–7489. [Google Scholar] [CrossRef]
- Chen, D.-Q.; Kolli, B.K.; Yadava, N.; Lu, H.G.; Gilman-Sachs, A.; Peterson, D.A.; Chang, K.-P. Episomal expression of specific sense and antisense mRNAs in Leishmania amazonensis: Modulation of gp63 level in promastigotes and their infection of macrophages in vitro. Infect. Immun. 2000, 68, 80–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, M.A.; Contreras, I.; Halle, M.; Tremblay, M.L.; McMaster, R.W.; Olivier, M. Leishmania GP63 alters host signaling through cleavage-activated protein tyrosine phosphatases. Sci. Signal. 2009, 2, ra58. [Google Scholar] [CrossRef]
- Corradin, S.; Ransijn, A.; Corradin, G.; Roggero, M.A.; Schmitz, A.A.; Schneider, P.; Mauël, J.; Vergères, G. MARCKS-related protein (MRP) is a substrate for the Leishmania major surface protease leishmanolysin (gp63). J. Biol. Chem. 1999, 274, 25411–25418. [Google Scholar] [CrossRef] [Green Version]
- Blanchette, J.; Racette, N.; Faure, R.; Siminovitch, K.A.; Olivier, M. Leishmania-induced increases in activation of macrophage SHP-1 tyrosine phosphatase are associated with impaired IFN-γ-triggered JAK2 activation. Eur. J. Immunol. 1999, 29, 3737–3744. [Google Scholar] [CrossRef]
- Jaramillo, M.; Gomez, M.A.; Larsson, O.; Shio, M.T.; Topisirovic, I.; Contreras, I.; Luxenburg, R.; Rosenfeld, A.; Colina, R.; McMaster, R.W. Leishmania repression of host translation through mTOR cleavage is required for parasite survival and infection. Cell Host Microbe 2011, 9, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, C.; Donhauser, N.; Döring, R.; Röllinghoff, M.; Diefenbach, A.; Rittig, M.G. Fibroblasts as host cells in latent leishmaniosis. J. Exp. Med. 2000, 191, 2121–2130. [Google Scholar] [CrossRef]
- Isnard, A.; Shio, M.T.; Olivier, M. Impact of Leishmania metalloprotease GP63 on macrophage signaling. Front. Cell Infect. Microbiol. 2012, 2, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.L.; Branquinha, M.H.; D’Avila-Levy, C.M. The ubiquitous gp63-like metalloprotease from lower trypanosomatids: In the search for a function. Anais Acad. Bras. Ciências 2006, 78, 687–714. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, G.; Chang, K.P. Acid protease activity of a major surface membrane glycoprotein (gp63) from Leishmania mexicana promastigotes. Mol. Biochem. Parasitol. 1988, 27, 43–52. [Google Scholar] [CrossRef]
- Moradin, N.; Descoteaux, A. Leishmania promastigotes: Building a safe niche within macrophages. Front. Cell. Infect. Microbiol. 2012, 2, 121. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; McSorley, S.J.; Chatfield, S.N.; Dougan, G.; Liew, F.Y. Protection against Leishmania major infection in genetically susceptible BALB/c mice by gp63 delivered orally in attenuated Salmonella typhimurium (AroA- AroD-). Immunology 1995, 85, 1–7. [Google Scholar]
- Reed, S.G.; Badaro, R.; Lloyd, R.M. Identification of specific and cross-reactive antigens of Leishmania donovani chagasi by human infection sera. J. Immunol. 1987, 138, 1596–1601. [Google Scholar]
- Vale, A.M.; Fujiwara, R.T.; da Neto, A.F.S.; Miret, J.A.; Alvarez, D.C.; da Silva, J.C.; Campos-Neto, A.; Reed, S.; Mayrink, W.; Nascimento, E. Identification of highly specific and cross-reactive antigens of Leishmania species by antibodies from Leishmania (Leishmania) chagasi naturally infected dogs. Zoonoses Public Health 2009, 56, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Medina, L.S.; Souza, B.A.; Queiroz, A.; Guimarães, L.H.; Machado, P.R.L.; Carvalho, E.M.; Wilson, M.E.; Schriefer, A. The gp63 Gene Cluster Is Highly Polymorphic in Natural Leishmania (Viannia) braziliensis Populations, but Functional Sites Are Conserved. PLoS ONE 2016, 11, e0163284. [Google Scholar] [CrossRef]
- Joshi, P.B.; Sacks, D.L.; Modi, G.; McMaster, W.R. Targeted gene deletion of Leishmania major genes encoding developmental stage-specific leishmanolysin (GP63). Mol. Microbiol. 1998, 27, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.B.; Kelly, B.L.; Kamhawi, S.; Sacks, D.L.; McMaster, W.R. Targeted gene deletion in Leishmania major identifies leishmanolysin (GP63) as a virulence factor. Mol. Biochem. Parasitol. 2002, 120, 33–40. [Google Scholar] [CrossRef]
- Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R. Ecto-nucleotidases and Ecto-phosphatases from Leishmania and Trypanosoma parasites. Sub-Cell. Biochem. 2014, 74, 217–252. [Google Scholar] [CrossRef]
- Leite, P.M.; Gomes, R.S.; Figueiredo, A.B.; Serafim, T.D.; Tafuri, W.L.; de Souza, C.C.; Moura, S.A.; Fietto, J.L.; Melo, M.N.; Ribeiro-Dias, F.; et al. Ecto-nucleotidase activities of promastigotes from Leishmania (Viannia) braziliensis relates to parasite infectivity and disease clinical outcome. PLoS Negl. Trop. Dis. 2012, 6, e1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, M.B.; Dwyer, D.M. Molecular and functional analyses of a novel class I secretory nuclease from the human pathogen, Leishmania donovani. J. Biol. Chem. 2007, 282, 10079–10095. [Google Scholar] [CrossRef] [Green Version]
- Boitz, J.M.; Ullman, B. A conditional mutant deficient in hypoxanthine-guanine phosphoribosyltransferase and xanthine phosphoribosyltransferase validates the purine salvage pathway of Leishmania donovani. J. Biol. Chem. 2006, 281, 16084–16089. [Google Scholar] [CrossRef] [Green Version]
- Peres, N.T.A.; Cunha, L.C.S.; Barbosa, M.L.A.; Santos, M.B.; de Oliveira, F.A.; de Jesus, A.M.R.; de Almeida, R.P. Infection of Human Macrophages by Leishmania infantum Is Influenced by Ecto-Nucleotidases. Front. Immunol 2017, 8, 1954. [Google Scholar] [CrossRef]
- de Souza, M.C.; de Assis, E.A.; Gomes, R.S.; da Ede, A.M.S.; Melo, M.N.; Fietto, J.L.; Afonso, L.C. The influence of ecto-nucleotidases on Leishmania amazonensis infection and immune response in C57B/6 mice. Acta Trop. 2010, 115, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes-Costa, A.B.; DeSouza-Vieira, T.S.; Paletta-Silva, R.; Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R.; Saraiva, E.M. 3’-nucleotidase/nuclease activity allows Leishmania parasites to escape killing by neutrophil extracellular traps. Infect. Immun. 2014, 82, 1732–1740. [Google Scholar] [CrossRef] [Green Version]
- Vasconcellos Rde, S.; Mariotini-Moura, C.; Gomes, R.S.; Serafim, T.D.; Firmino Rde, C.; Silva, E.B.M.; Castro, F.F.; Oliveira, C.M.; Borges-Pereira, L.; de Souza, A.C.; et al. Leishmania infantum ecto-nucleoside triphosphate diphosphohydrolase-2 is an apyrase involved in macrophage infection and expressed in infected dogs. PLoS Negl. Trop. Dis. 2014, 8, e3309. [Google Scholar] [CrossRef]
- Paletta-Silva, R.; Vieira, D.P.; Vieira-Bernardo, R.; Majerowicz, D.; Gondim, K.C.; Vannier-Santos, M.A.; Lopes, A.H.; Meyer-Fernandes, J.R. Leishmania amazonensis: Characterization of an ecto-3’-nucleotidase activity and its possible role in virulence. Exp. Parasitol. 2011, 129, 277–283. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Hombach, A.; Clos, J.; Tatu, U. No stress—Hsp90 and signal transduction in Leishmania. Parasitology 2014, 141, 1156–1166. [Google Scholar] [CrossRef]
- Kröber-Boncardo, C.; Grünebast, J.; Clos, J. Heat Shock Proteins in Leishmania Parasites; Springer: Dordrecht, The Netherlands, 2020; pp. 1–20. [Google Scholar]
- Krobitsch, S.; Clos, J. A novel role for 100 kD heat shock proteins in the parasite Leishmania donovani. Cell Stress Chaperones 1999, 4, 191. [Google Scholar] [CrossRef]
- Hübel, A.; Krobitsch, S.; Hörauf, A.; Clos, J. Leishmania major Hsp100 is required chiefly in the mammalian stage of the parasite. Mol. Cell. Biol. 1997, 17, 5987–5995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K. Leishmania exosomes modulate innate and adaptive immune responses through effects on monocytes and dendritic cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, M.; Gabdank, I.; Ben-David, M.; Zilka, A.; Orr, I.; Barash, D.; Shapira, M. Preferential translation of Hsp83 in Leishmania requires a thermosensitive polypyrimidine-rich element in the 3’ UTR and involves scanning of the 5’ UTR. RNA 2010, 16, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Young, J.C.; Agashe, V.R.; Siegers, K.; Hartl, F.U. Pathways of chaperone-mediated protein folding in the cytosol. Nat. Rev. Mol. Cell Biol. 2004, 5, 781–791. [Google Scholar] [CrossRef]
- Clos, J.; Krobitsch, S. Heat Shock as a Regular Feature of the Life Cycle of Leishmania Parasites. Am. Zool. 1999, 39, 848–856. [Google Scholar] [CrossRef] [Green Version]
- Hombach, A.; Ommen, G.; MacDonald, A.; Clos, J. A small heat shock protein is essential for thermotolerance and intracellular survival of Leishmania donovani. J. Cell Sci. 2014, 127, 4762–4773. [Google Scholar] [CrossRef] [Green Version]
- Wiesgigl, M.; Clos, J. Heat shock protein 90 homeostasis controls stage differentiation in Leishmania donovani. Mol. Biol. Cell 2001, 12, 3307–3316. [Google Scholar] [CrossRef] [Green Version]
- Bifeld, E.; Lorenzen, S.; Bartsch, K.; Vasquez, J.-J.; Siegel, T.N.; Clos, J. Ribosome profiling reveals HSP90 inhibitor effects on stage-specific protein synthesis in Leishmania donovani. Msystems 2018, 3, e00214. [Google Scholar] [CrossRef] [Green Version]
- Hombach, A.; Ommen, G.; Chrobak, M.; Clos, J. The Hsp 90–Sti 1 interaction is critical for L eishmania donovani proliferation in both life cycle stages. Cell. Microbiol. 2013, 15, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.A.; Watanabe, R.; Dacher, M.; Chafey, P.; Fortéa, J.O.; Scott, D.A.; Beverley, S.M.; Ommen, G.; Clos, J.; Hem, S. Phosphoproteome dynamics reveal heat-shock protein complexes specific to the Leishmania donovani infectious stage. Proc. Natl. Acad. Sci. USA 2010, 107, 8381–8386. [Google Scholar] [CrossRef] [Green Version]
- Adhuna, P.S.; Salotra, P.; Mukhopadhyay, B.; Bhatnagar, R. Modulation of macrophage heat shock proteins (HSPs) expression in response to intracellular infection by virulent and avirulent strains of Leishmania donovani. Biochem. Mol. Biol. Int. 1997, 43, 1265–1275. [Google Scholar] [CrossRef]
- Rico, A.I.; Del Real, G.; Soto, M.; Quijada, L.; Martinez, A.C.; Alonso, C.; Requena, J.M. Characterization of the immunostimulatory properties of Leishmania infantum HSP70 by fusion to the Escherichia coli maltose-binding protein in normal and nu/nu BALB/c mice. Infect. Immun. 1998, 66, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Skeiky, Y.A.; Benson, D.R.; Guderian, J.A.; Whittle, J.A.; Bacelar, O.; Carvalho, E.M.; Reed, S.G. Immune responses of leishmaniasis patients to heat shock proteins of Leishmania species and humans. Infect. Immun. 1995, 63, 4105–4114. [Google Scholar] [CrossRef] [Green Version]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef]
- Doige, C.A.; Ames, G.F. ATP-dependent transport systems in bacteria and humans: Relevance to cystic fibrosis and multidrug resistance. Annu. Rev. Microbiol. 1993, 47, 291–319. [Google Scholar] [CrossRef]
- Piper, R.C.; Xu, X.; Russell, D.G.; Little, B.M.; Landfear, S.M. Differential targeting of two glucose transporters from Leishmania enriettii is mediated by an NH2-terminal domain. J. Cell Biol. 1995, 128, 499–508. [Google Scholar] [CrossRef]
- Perez-Victoria, J.M.; Di Pietro, A.; Barron, D.; Ravelo, A.G.; Castanys, S.; Gamarro, F. Multidrug resistance phenotype mediated by the P-glycoprotein-like transporter in Leishmania: A search for reversal agents. Curr. Drug Targets 2002, 3, 311–333. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.; Leprohon, P.; Drummelsmith, J.; Ouellette, M. Growth phase regulation of the main folate transporter of Leishmania infantum and its role in methotrexate resistance. J. Biol. Chem. 2004, 279, 54494–54501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, J.I.; Garcia-Hernandez, R.; Castanys, S.; Gamarro, F. A new ABC half-transporter in Leishmania major is involved in resistance to antimony. Antimicrob. Agents Chemother. 2013, 57, 3719–3730. [Google Scholar] [CrossRef] [Green Version]
- Prati, F.; Goldman-Pinkovich, A.; Lizzi, F.; Belluti, F.; Koren, R.; Zilberstein, D.; Bolognesi, M.L. Quinone-amino acid conjugates targeting Leishmania amino acid transporters. PLoS ONE 2014, 9, e107994. [Google Scholar] [CrossRef]
- Parodi-Talice, A.; Araujo, J.M.; Torres, C.; Perez-Victoria, J.M.; Gamarro, F.; Castanys, S. The overexpression of a new ABC transporter in Leishmania is related to phospholipid trafficking and reduced infectivity. Biochim. Biophys. Acta 2003, 1612, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Ouellette, M.; Legare, D.; Haimeur, A.; Grondin, K.; Roy, G.; Brochu, C.; Papadopoulou, B. ABC transporters in Leishmania and their role in drug resistance. Drug Resist. Updates 1998, 1, 43–48. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmahallawy, E.K.; Alkhaldi, A.A.M. Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite. Vet. Sci. 2021, 8, 33. https://doi.org/10.3390/vetsci8020033
Elmahallawy EK, Alkhaldi AAM. Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite. Veterinary Sciences. 2021; 8(2):33. https://doi.org/10.3390/vetsci8020033
Chicago/Turabian StyleElmahallawy, Ehab Kotb, and Abdulsalam A. M. Alkhaldi. 2021. "Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite" Veterinary Sciences 8, no. 2: 33. https://doi.org/10.3390/vetsci8020033
APA StyleElmahallawy, E. K., & Alkhaldi, A. A. M. (2021). Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite. Veterinary Sciences, 8(2), 33. https://doi.org/10.3390/vetsci8020033