
Establishment of a Newborn Lamb Gut-Loop Model to Evaluate New Methods of Enteric Disease Control and Reduce Experimental Animal Use

 , , and
, , and 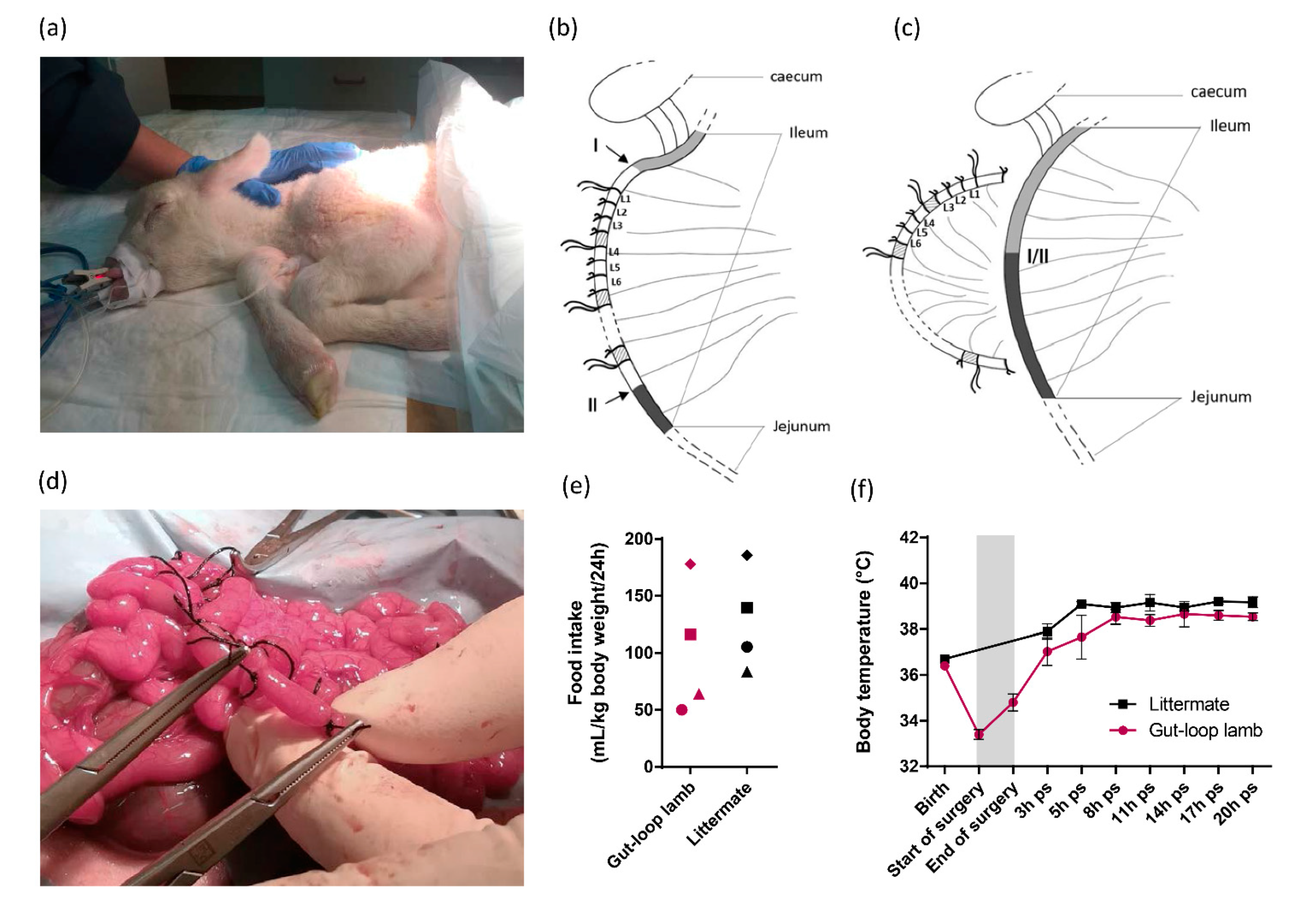{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Development of a New Animal Model
2.1.1. Ethic Statements
2.1.2. Neonatal Care of Newborn Lambs after Cesarean Surgery
2.1.3. Intestinal Loop Surgery
2.2. Use of the Gut-Loop Model for Investigating Immune Responses and Infection with C. parvum
2.2.1. Intestinal Stimulations
2.2.2. RNA Extraction and RT-qPCR
2.2.3. C. parvum Infection
2.2.4. Parasite Burden
2.2.5. Histology and Immunofluorescence
2.2.6. Statistics
3. Results
3.1. Establishment of Gut-Loop Model on a Cesarean-Born Neonatal Lamb
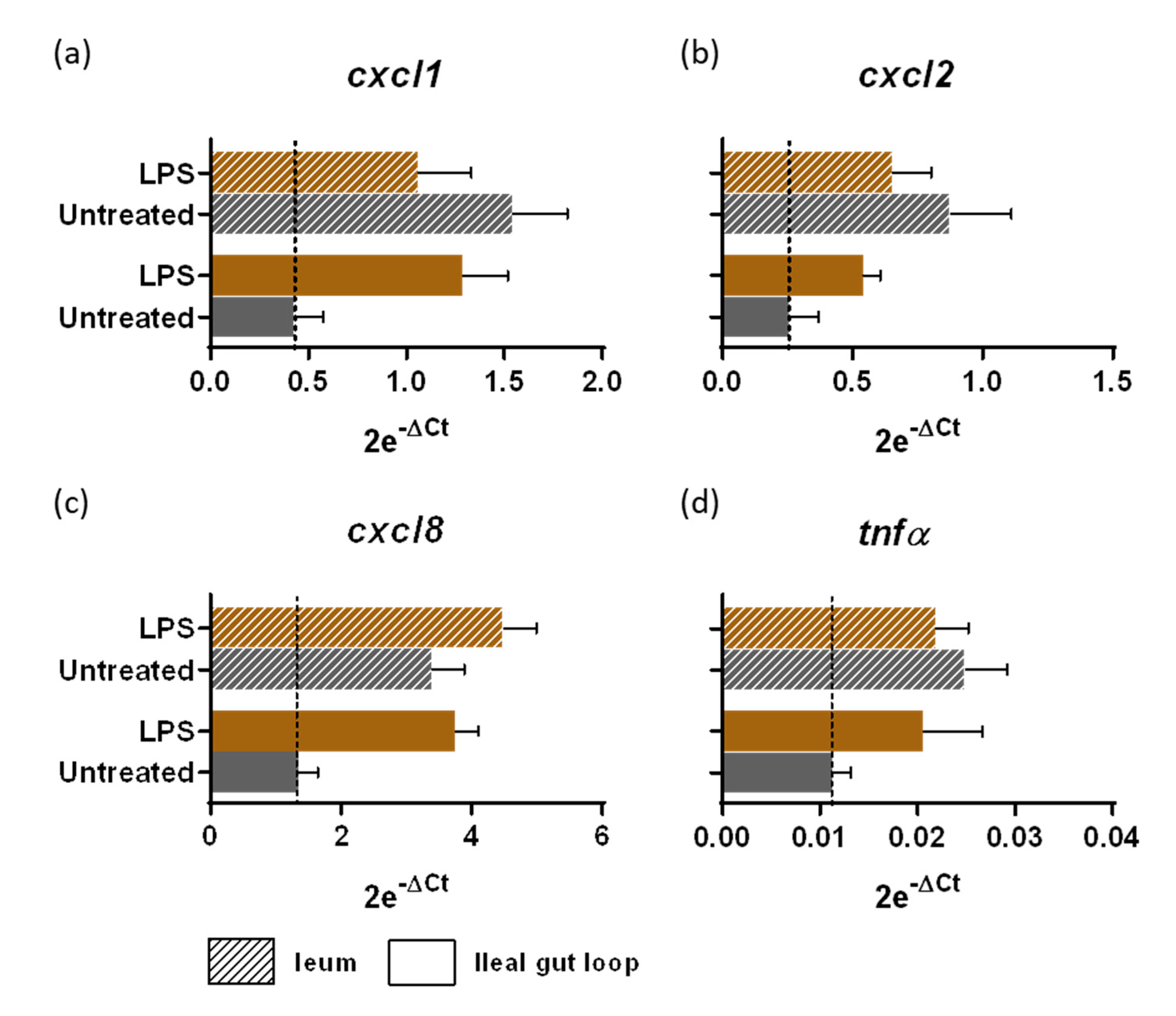
3.2. Innate Immune Response to LPS Stimulation of Sterile Gut Loop
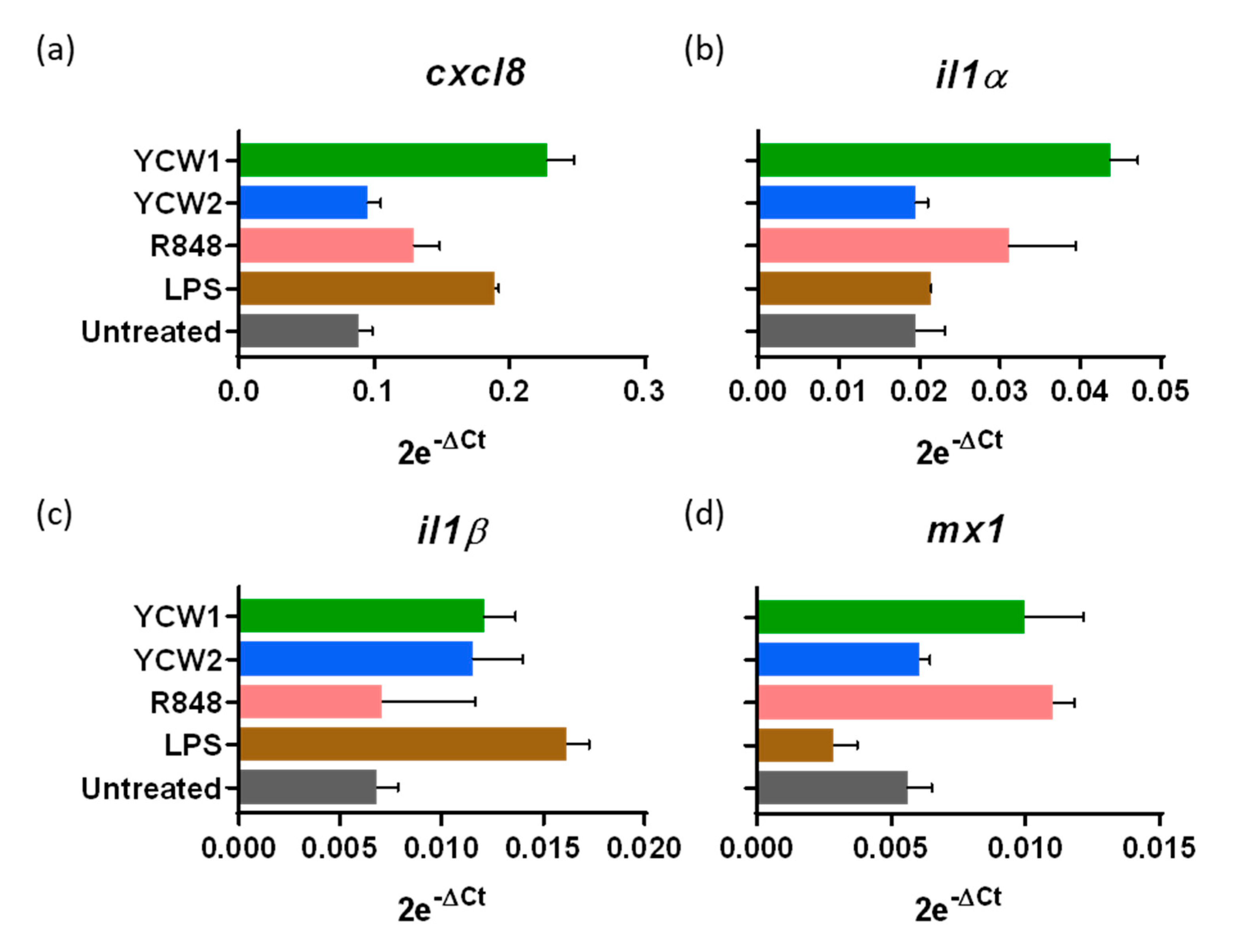
3.3. Innate Ileal Immune Response to TLR-Ligands and YCW Fractions in Isolated Loops
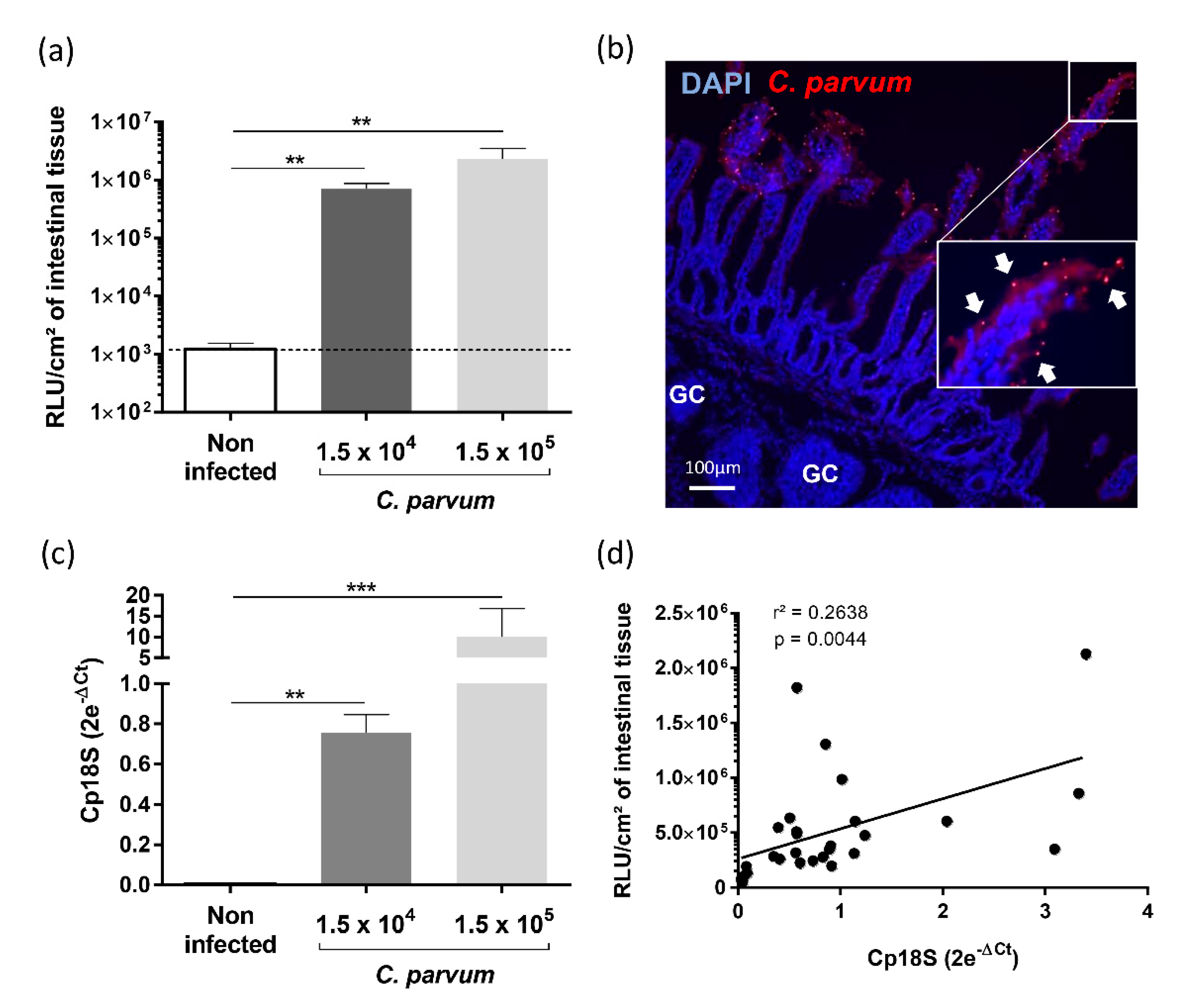
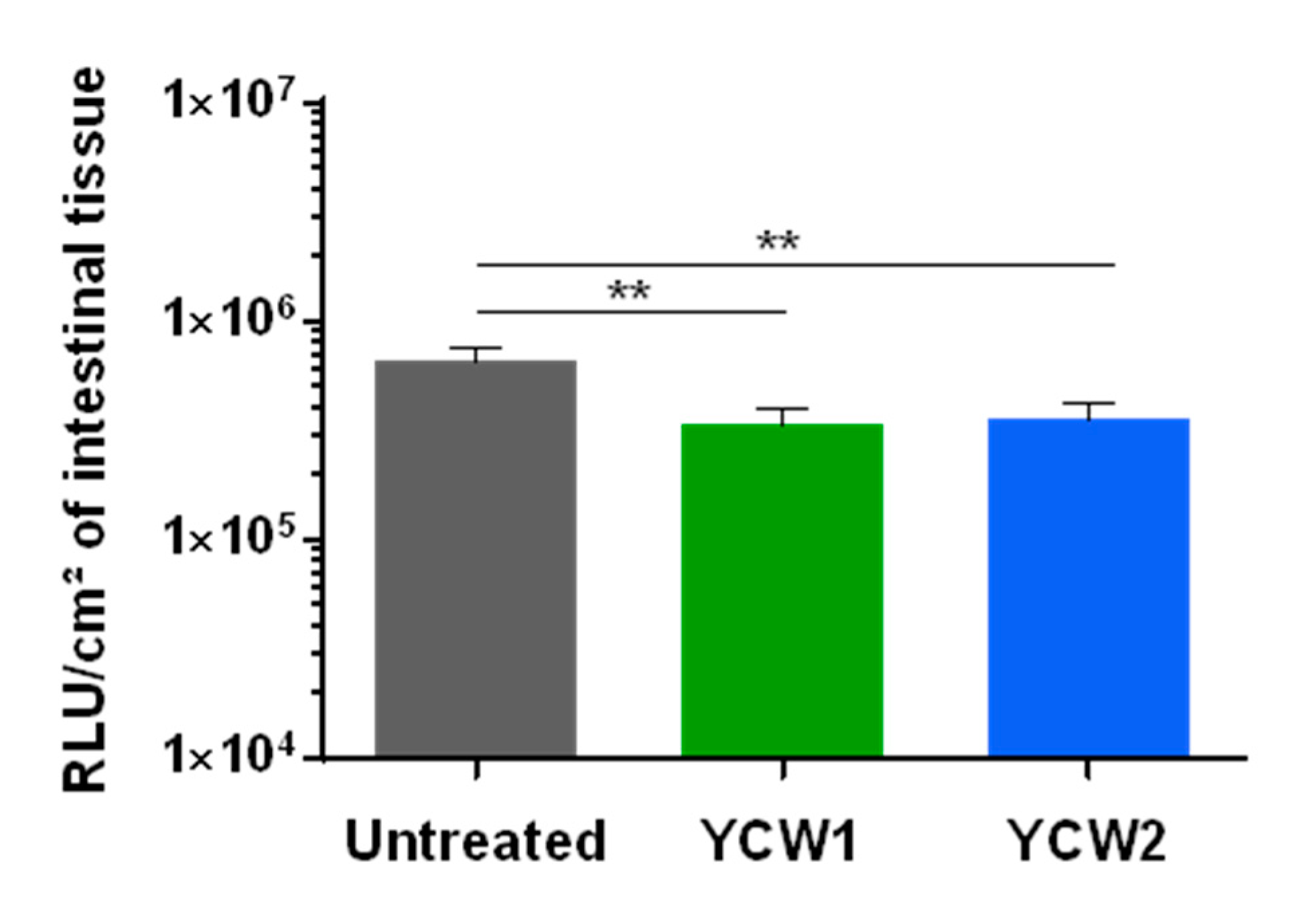
3.4. Evaluation of YCW Fractions on Early Cryptosporidium parvum Invasion and Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayr, L.M.; Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 2009, 9, 580–588. [Google Scholar] [CrossRef]
- Brodin, P.; Christophe, T. High-content screening in infectious diseases. Curr. Opin. Chem. Biol. 2011, 15, 534–539. [Google Scholar] [CrossRef]
- Li, V.S.W. Modelling intestinal inflammation and infection using ‘mini-gut’ organoids. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Blanc, F.; Cherbuy, C.; Egidy, G.; Giuffra, E.; Lacroix-Lamande, S.; Wiedemann, A. Intestinal organoids in farm animals. Vet. Res. 2021, 52, 33. [Google Scholar] [CrossRef]
- Richmond, J. The 3Rs—Past, present and future. Scand. J. Lab. Anim. Sci. 2000, 27, 84–92. [Google Scholar]
- Pinto, D.J.; Vinayak, S. Cryptosporidium: Host-Parasite Interactions and Pathogenesis. Curr. Clin. Microbiol. Rep. 2021, 8, 62–67. [Google Scholar] [CrossRef]
- Gururajan, A.; Rajkumari, N.; Devi, U.; Borah, P. Cryptosporidium and waterborne outbreaks—A mini review. Trop. Parasitol. 2021, 11, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef] [Green Version]
- Striepen, B. Parasitic infections: Time to tackle cryptosporidiosis. Nature 2013, 503, 189–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefay, D.; Naciri, M.; Poirier, P.; Chermette, R. Prevalence of Cryptosporidium infection in calves in France. Vet. Parasitol. 2000, 89, 1–9. [Google Scholar] [CrossRef]
- Van Voorhis, W.C.; Hulverson, M.A.; Choi, R.; Huang, W.; Arnold, S.L.M.; Schaefer, D.A.; Betzer, D.P.; Vidadala, R.S.R.; Lee, S.; Whitman, G.R.; et al. One health therapeutics: Target-Based drug development for cryptosporidiosis and other apicomplexa diseases. Vet. Parasitol. 2021, 289, 109336. [Google Scholar] [CrossRef]
- Lotz, M.; Gutle, D.; Walther, S.; Menard, S.; Bogdan, C.; Hornef, M.W. Postnatal acquisition of endotoxin tolerance in intestinal epithelial cells. J. Exp. Med. 2006, 203, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Chassin, C.; Kocur, M.; Pott, J.; Duerr, C.U.; Gutle, D.; Lotz, M.; Hornef, M.W. miR-146a mediates protective innate immune tolerance in the neonate intestine. Cell Host Microbe 2010, 8, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutwiri, G.; Watts, T.; Lew, L.; Beskorwayne, T.; Papp, Z.; Baca-Estrada, M.E.; Griebel, P. Ileal and jejunal Peyer’s patches play distinct roles in mucosal immunity of sheep. Immunology 1999, 97, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Gerdts, V.; Uwiera, R.R.; Mutwiri, G.K.; Wilson, D.J.; Bowersock, T.; Kidane, A.; Babiuk, L.A.; Griebel, P.J. Multiple intestinal ‘loops’ provide an in vivo model to analyse multiple mucosal immune responses. J. Immunol. Methods 2001, 256, 19–33. [Google Scholar] [CrossRef]
- Swale, C.; Bougdour, A.; Gnahoui-David, A.; Tottey, J.; Georgeault, S.; Laurent, F.; Palencia, A.; Hakimi, M.A. Metal-captured inhibition of pre-mRNA processing activity by CPSF3 controls Cryptosporidium infection. Sci. Transl. Med. 2019, 11, eaax7161. [Google Scholar] [CrossRef] [PubMed]
- Lantier, L.; Lacroix-Lamande, S.; Potiron, L.; Metton, C.; Drouet, F.; Guesdon, W.; Gnahoui-David, A.; Le Vern, Y.; Deriaud, E.; Fenis, A.; et al. Intestinal CD103+ dendritic cells are key players in the innate immune control of Cryptosporidium parvum infection in neonatal mice. PLoS Pathog. 2013, 9, e1003801. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.; van de Veerdonk, F.L.; Brown, A.J.; Netea, M.G. Candida albicans morphogenesis and host defence: Discriminating invasion from colonization. Nat. Rev. Microbiol. 2011, 10, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Urie, N.J.; Lombard, J.E.; Shivley, C.B.; Kopral, C.A.; Adams, A.E.; Earleywine, T.J.; Olson, J.D.; Garry, F.B. Preweaned heifer management on US dairy operations: Part V. Factors associated with morbidity and mortality in preweaned dairy heifer calves. J. Dairy Sci. 2018, 101, 9229–9244. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.; Kelton, D.F.; Duffield, T.F.; Renaud, D.L. Risk factors identified on arrival associated with morbidity and mortality at a grain-fed veal facility: A prospective, single-cohort study. J. Dairy Sci. 2019, 102, 9224–9235. [Google Scholar] [CrossRef]
- Chen, W.; Harp, J.A.; Harmsen, A.G. Cryptosporidium parvum infection in gene-targeted B cell-deficient mice. J. Parasitol. 2003, 89, 391–393. [Google Scholar] [CrossRef]
- Bond, C. Evaluation of lamb colostrum supplements. Vet. Rec. 2020, 187, e100. [Google Scholar] [CrossRef]
- Chang, M.N.; Wei, J.Y.; Hao, L.Y.; Ma, F.T.; Li, H.Y.; Zhao, S.G.; Sun, P. Effects of different types of zinc supplement on the growth, incidence of diarrhea, immune function, and rectal microbiota of newborn dairy calves. J. Dairy Sci. 2020, 103, 6100–6113. [Google Scholar] [CrossRef]
- Renaud, D.L.; Kelton, D.F.; Weese, J.S.; Noble, C.; Duffield, T.F. Evaluation of a multispecies probiotic as a supportive treatment for diarrhea in dairy calves: A randomized clinical trial. J. Dairy Sci. 2019, 102, 4498–4505. [Google Scholar] [CrossRef]
- Cangiano, L.R.; Yohe, T.T.; Steele, M.A.; Renaud, D.L. INVITED REVIEW: Strategic use of microbial-based probiotics and prebiotics in dairy calf rearing. Appl. Anim. Sci. 2020, 36, 630–651. [Google Scholar] [CrossRef]
- Davis, M.E.; Maxwell, C.V.; Erf, G.F.; Brown, D.C.; Wistuba, T.J. Dietary supplementation with phosphorylated mannans improves growth response and modulates immune function of weanling pigs. J. Anim. Sci. 2004, 82, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Galvao, K.N.; Santos, J.E.; Coscioni, A.; Villasenor, M.; Sischo, W.M.; Berge, A.C. Effect of feeding live yeast products to calves with failure of passive transfer on performance and patterns of antibiotic resistance in fecal Escherichia coli. Reprod. Nutr. Dev. 2005, 45, 427–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammeri, M.; Chevillot, A.; Thomas, M.; Polack, B.; Julien, C.; Marden, J.P.; Auclair, E.; Vallee, I.; Adjou, K.T. Efficacy of chitosan, a natural polysaccharide, against Cryptosporidium parvum in vitro and in vivo in neonatal mice. Exp. Parasitol. 2018, 194, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lucey, P.M.; Lean, I.J.; Aly, S.S.; Golder, H.M.; Block, E.; Thompson, J.S.; Rossow, H.A. Effects of mannan-oligosaccharide and Bacillus subtilis supplementation to preweaning Holstein dairy heifers on body weight gain, diarrhea, and shedding of fecal pathogens. J. Dairy Sci. 2021, 104, 4290–4302. [Google Scholar] [CrossRef]
- Ganner, A.; Schatzmayr, G. Capability of yeast derivatives to adhere enteropathogenic bacteria and to modulate cells of the innate immune system. Appl. Microbiol. Biotechnol. 2012, 95, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Posadas, G.A.; Broadway, P.R.; Thornton, J.A.; Carroll, J.A.; Lawrence, A.; Corley, J.R.; Thompson, A.; Donaldson, J.R. Yeast Pro- and Paraprobiotics Have the Capability to Bind Pathogenic Bacteria Associated with Animal Disease1. Transl. Anim. Sci. 2017, 1, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Joe, A.; Hamer, D.H.; Kelley, M.A.; Pereira, M.E.; Keusch, G.T.; Tzipori, S.; Ward, H.D. Role of a Gal/GalNAc-specific sporozoite surface lectin in Cryptosporidium parvum-host cell interaction. J. Eukaryot. Microbiol. 1994, 41, 44S. [Google Scholar] [PubMed]
- Ludington, J.G.; Ward, H.D. The Cryptosporidium parvum C-Type Lectin CpClec Mediates Infection of Intestinal Epithelial Cells via Interactions with Sulfated Proteoglycans. Infect. Immun. 2016, 84, 1593–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, M.; Jenne, C.N.; Kennedy, L.J.; Reynolds, J.D. The sheep and cattle Peyer’s patch as a site of B-cell development. Vet. Res. 2006, 37, 401–415. [Google Scholar] [CrossRef] [Green Version]
- Walachowski, S.; Tabouret, G.; Foucras, G. Triggering Dectin-1-Pathway Alone Is Not Sufficient to Induce Cytokine Production by Murine Macrophages. PLoS ONE 2016, 11, e0148464. [Google Scholar] [CrossRef] [Green Version]
- Meurens, F.; Berri, M.; Auray, G.; Melo, S.; Levast, B.; Virlogeux-Payant, I.; Chevaleyre, C.; Gerdts, V.; Salmon, H. Early immune response following Salmonella enterica subspecies enterica serovar Typhimurium infection in porcine jejunal gut loops. Vet. Res. 2009, 40, 5. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baillou, A.; Kasal-Hoc, N.; Barc, C.; Cognié, J.; Pinard, A.; Pezant, J.; Schulthess, J.; Peltier-Pain, P.; Lacroix-Lamandé, S.; Laurent, F. Establishment of a Newborn Lamb Gut-Loop Model to Evaluate New Methods of Enteric Disease Control and Reduce Experimental Animal Use. Vet. Sci. 2021, 8, 170. https://doi.org/10.3390/vetsci8090170
Baillou A, Kasal-Hoc N, Barc C, Cognié J, Pinard A, Pezant J, Schulthess J, Peltier-Pain P, Lacroix-Lamandé S, Laurent F. Establishment of a Newborn Lamb Gut-Loop Model to Evaluate New Methods of Enteric Disease Control and Reduce Experimental Animal Use. Veterinary Sciences. 2021; 8(9):170. https://doi.org/10.3390/vetsci8090170
Chicago/Turabian StyleBaillou, Ambre, Nathalie Kasal-Hoc, Céline Barc, Juliette Cognié, Anne Pinard, Jérémy Pezant, Julie Schulthess, Pauline Peltier-Pain, Sonia Lacroix-Lamandé, and Fabrice Laurent. 2021. "Establishment of a Newborn Lamb Gut-Loop Model to Evaluate New Methods of Enteric Disease Control and Reduce Experimental Animal Use" Veterinary Sciences 8, no. 9: 170. https://doi.org/10.3390/vetsci8090170
APA StyleBaillou, A., Kasal-Hoc, N., Barc, C., Cognié, J., Pinard, A., Pezant, J., Schulthess, J., Peltier-Pain, P., Lacroix-Lamandé, S., & Laurent, F. (2021). Establishment of a Newborn Lamb Gut-Loop Model to Evaluate New Methods of Enteric Disease Control and Reduce Experimental Animal Use. Veterinary Sciences, 8(9), 170. https://doi.org/10.3390/vetsci8090170