
The First Report of Elaphostrongylus cervi Infection in Two Imported Wapitis (Cervus canadensis) in Slovenia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Case History
3. Results
3.1. Case 1
3.1.1. Blood and Biochemical Results
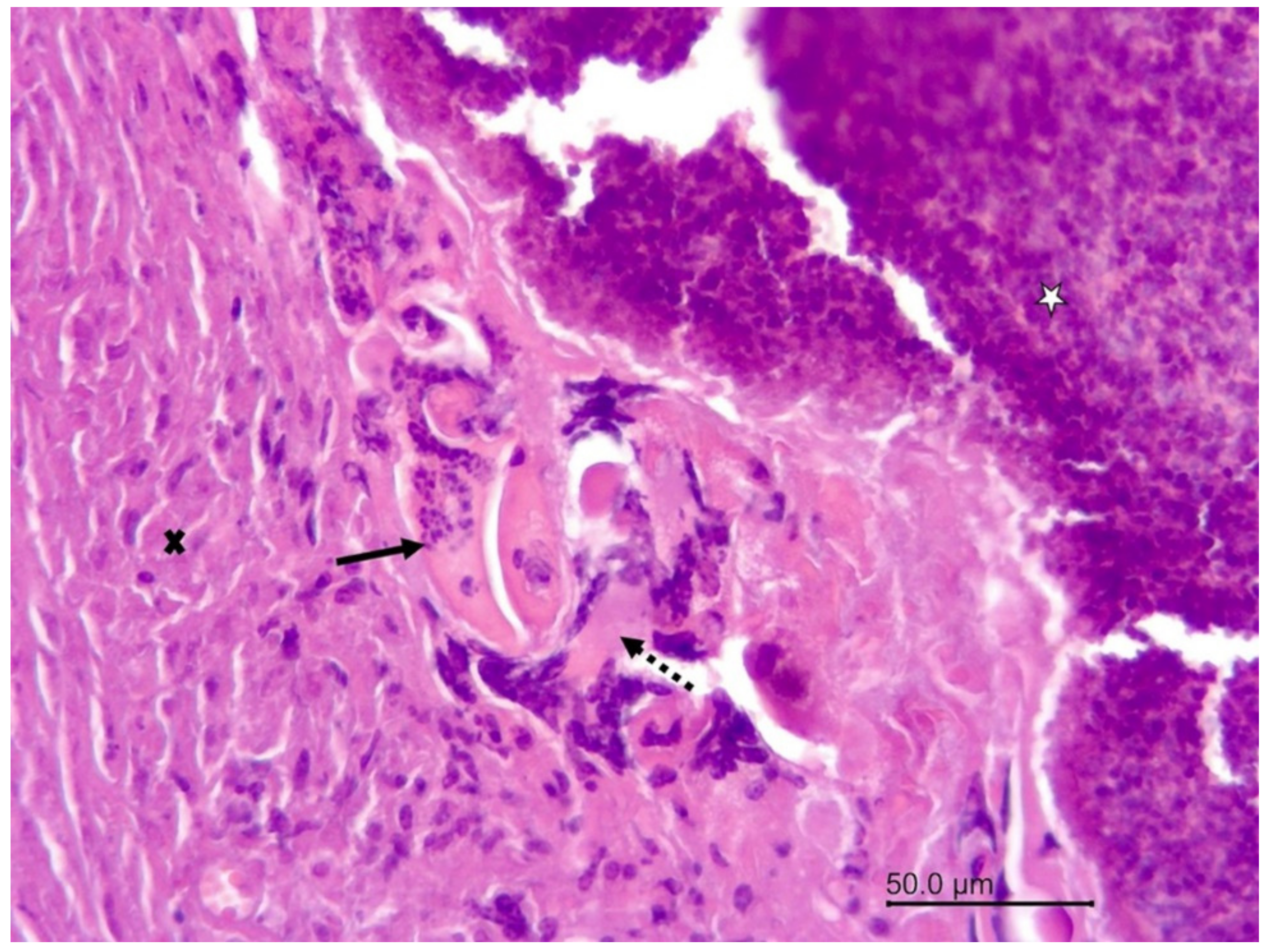
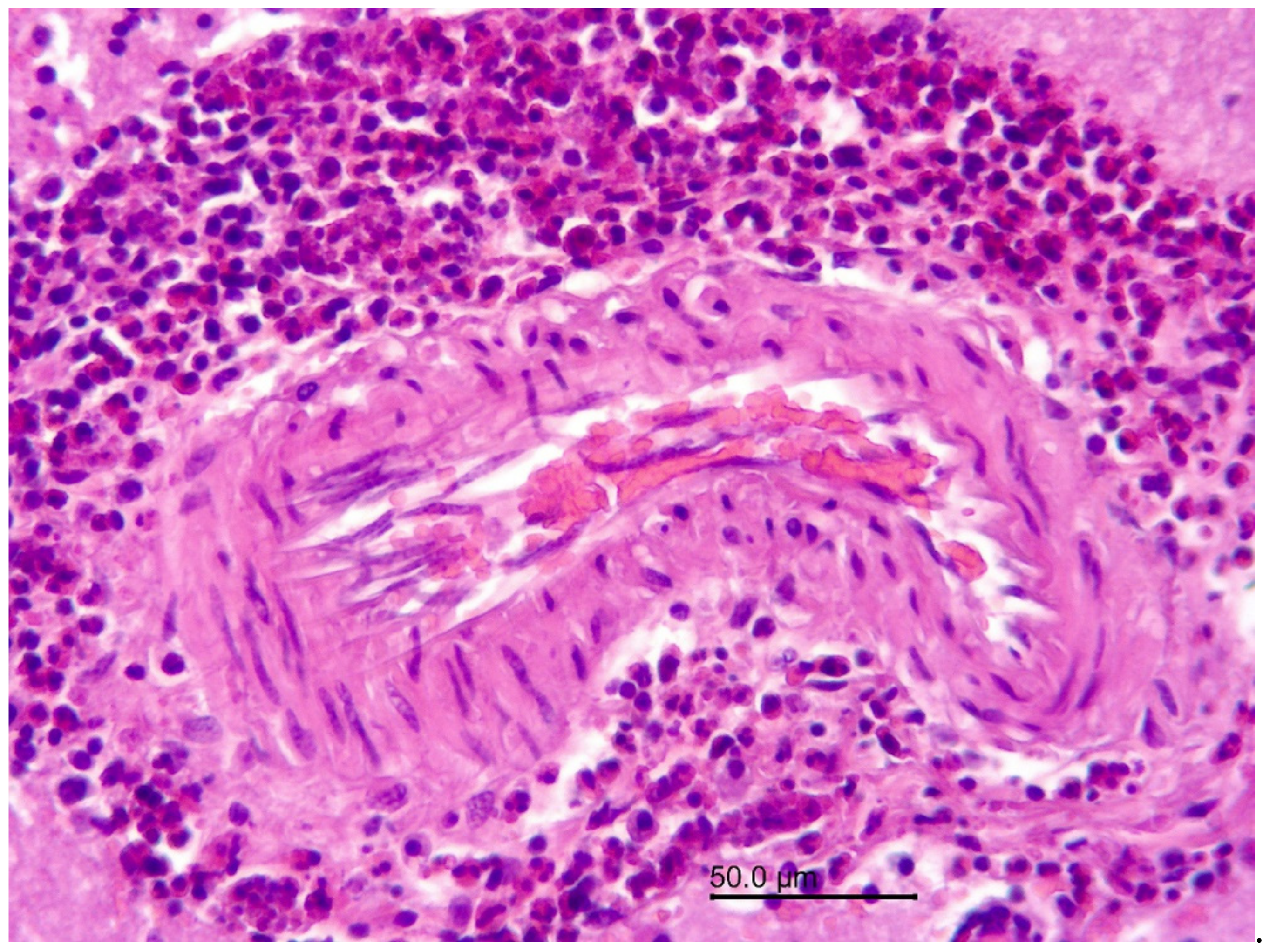
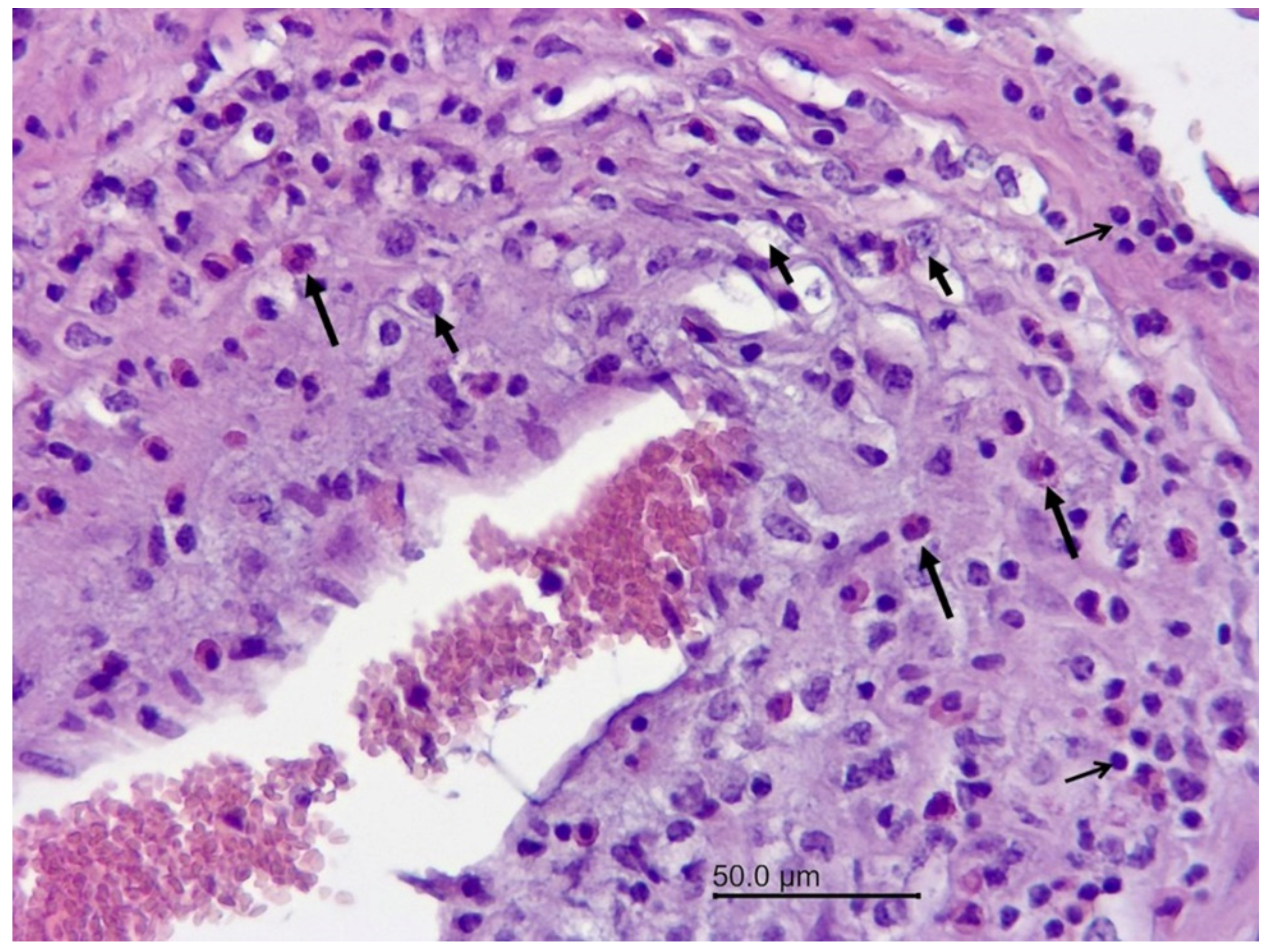
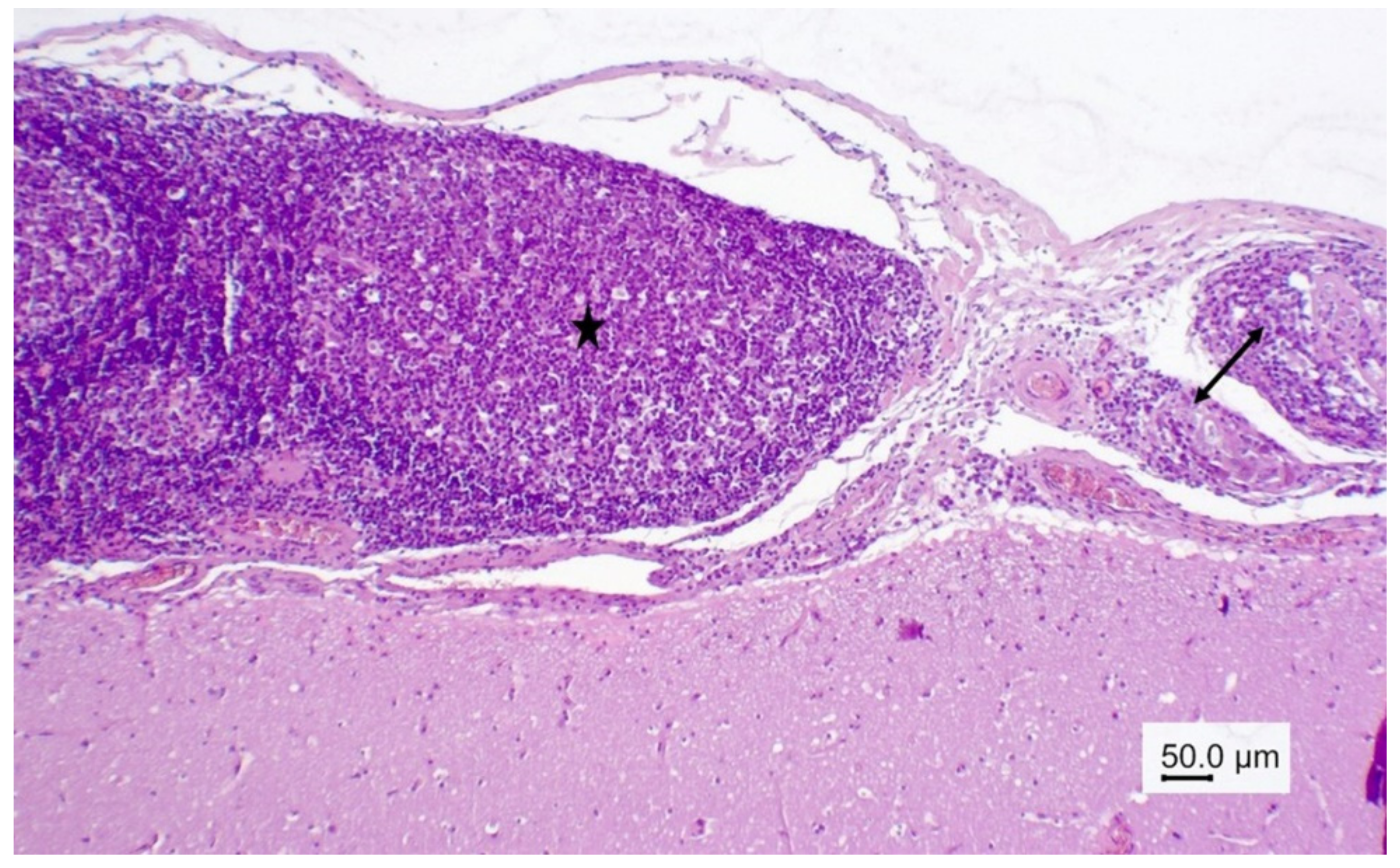
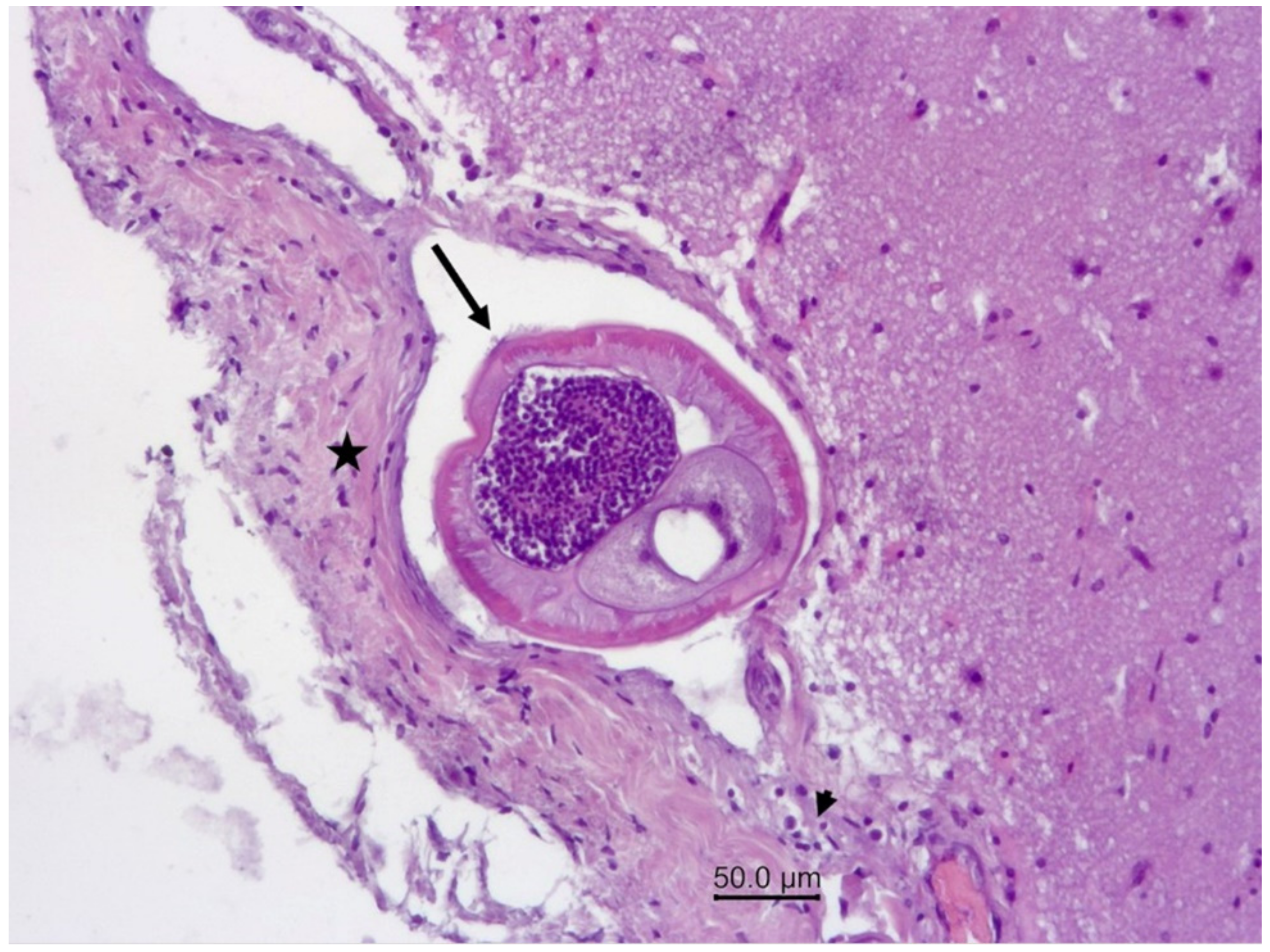
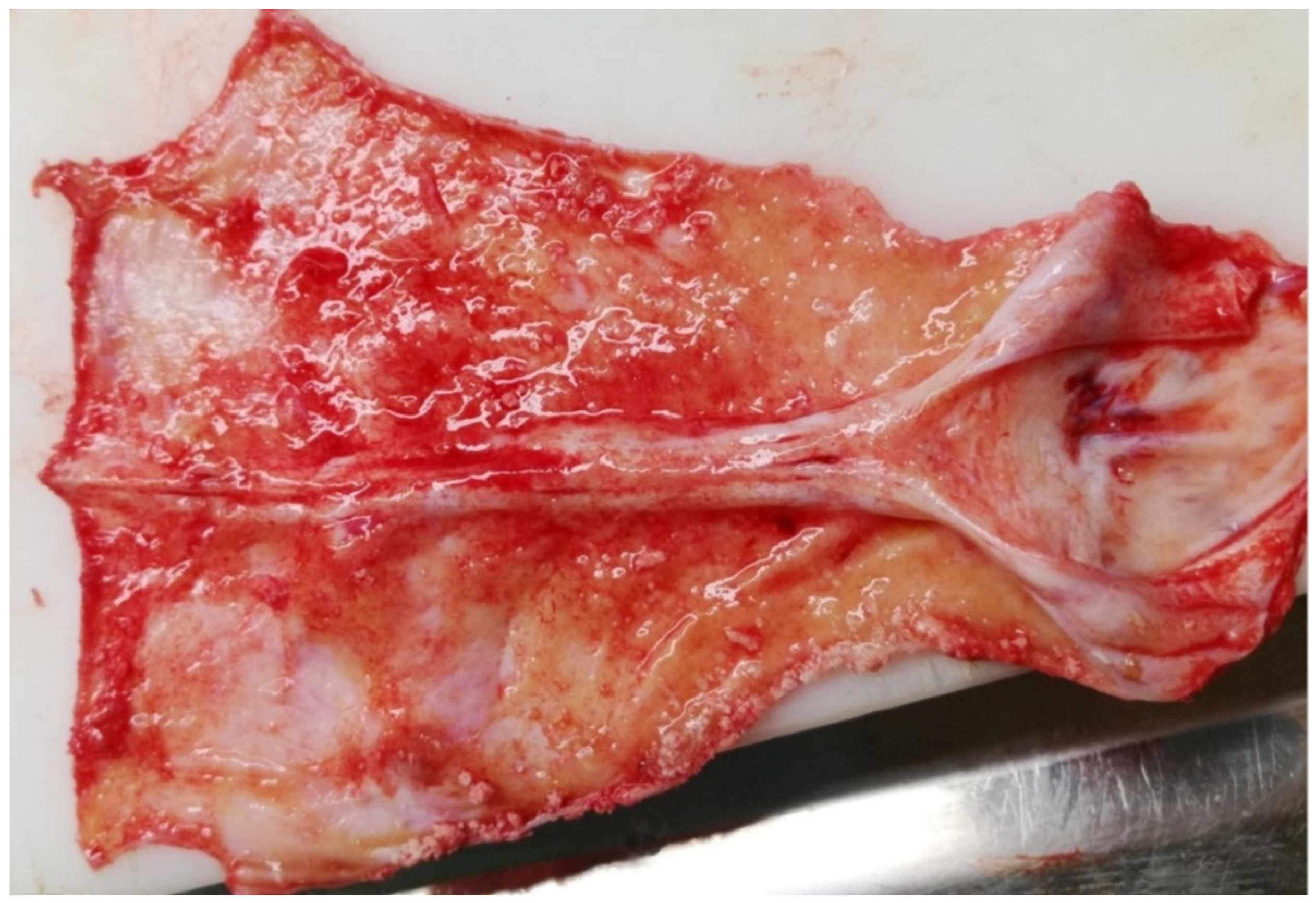
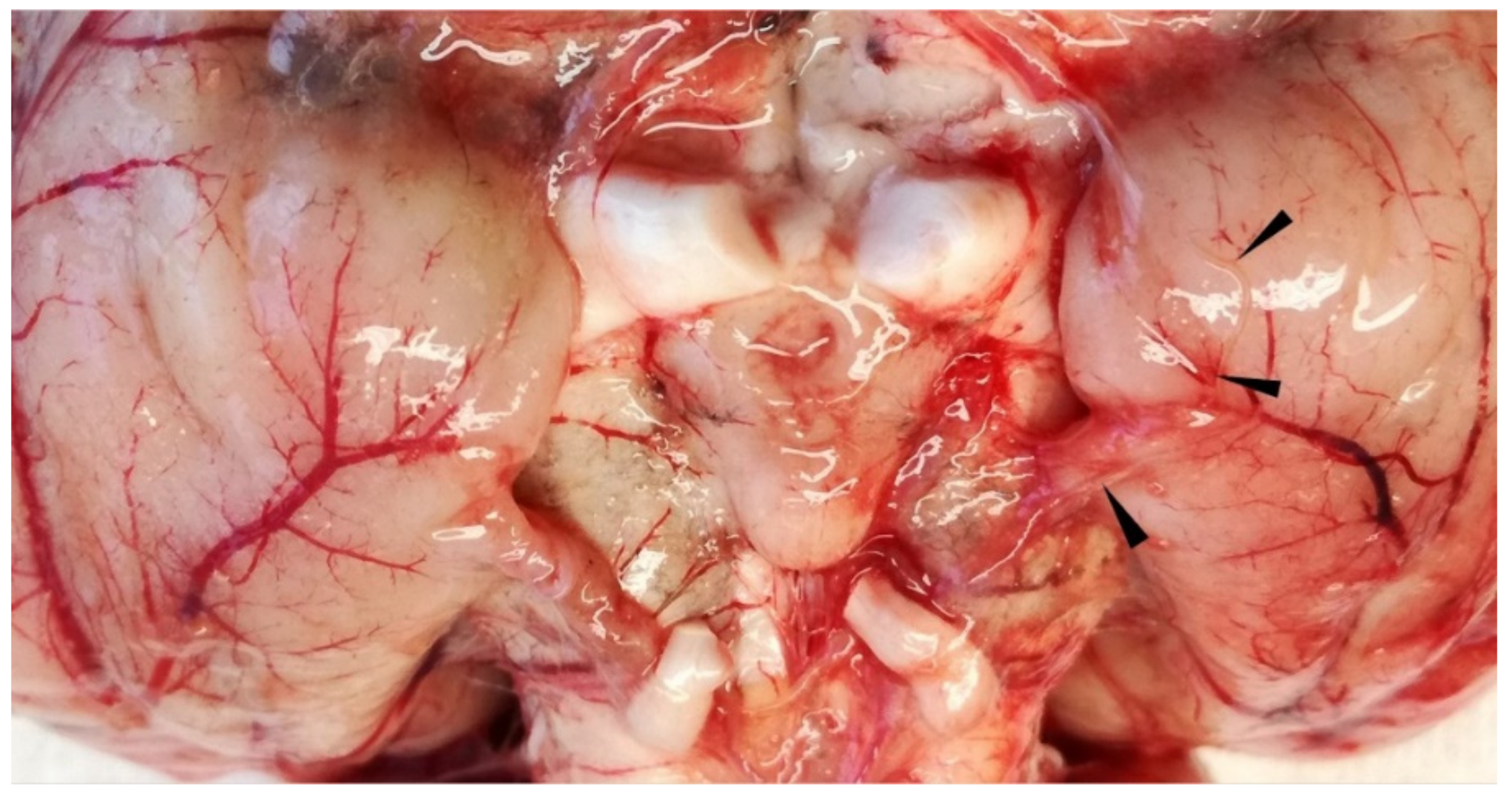
3.1.2. Necropsy and Histopathology
3.2. Case 2
3.2.1. Necropsy and Histopathology
3.2.2. Parasitological Examination
3.2.3. Molecular Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gajadhar, A.A.; Tessaro, S.V.; Yates, W.D. Diagnosis of Elaphostrongylus cervi infection in New Zealand red deer (Cervus elaphus) quarantined in Canada, and experimental determination of a new extended prepatent period. Can. Vet. J. 1994, 35, 433–437. [Google Scholar]
- Mason, P.C. Elaphostrongylus cervi—A review. Surveillance 1989, 16, 3–10. [Google Scholar]
- Lankester, M.W. Extrapulmonary lungworms of cervids. In Parasitic Diseases of Wild Mammals, 2nd ed.; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Manson Publishing/The Veterinary Press: London, UK, 2001. [Google Scholar]
- Davidson, R.K.; Mørk, T.; Holmgren, K.E.; Oksanen, A. Infection with brainworm (Elaphostrongylus rangiferi) in reindeer (Rangifer tarandus ssp.) in Fennoscandia. Acta Vet. Scand. 2020, 62, 24. [Google Scholar] [CrossRef] [PubMed]
- Stéen, M.; Chabaud, A.G.; Rehbinder, C. Species of the genus Elaphostrongylus parasite of Swedish Cervidae. A description of E. alces n.sp. Ann. Parasitol. Hum. Comp. 1989, 64, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankester, M.W.; Crichton, V.J.; Timmermann, H.R. A protostrongylid nematode (Strongylida: Protostrongylidae) in woodland caribou (Rangifer tarandus caribou). Can. J. Zool. 1976, 54, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.C.; Kiddey, N.R.; Sutherland, R.J.; Rutherford, D.M.; Green, A.G. Letter: Elaphostrongylus cervi in red deer. N. Z. Vet. J. 1976, 24, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Said, Y.; Cabaret, J.; Harmache, A.; Lahmar, S. Identification of first-stage dorsal-spined lungworm larvae of Tunisian barbary red deer: First report of Varestrongylus sagittatus and Elaphostrongylus cervi in Africa. Parasitol. Int. 2018, 67, 386–388. [Google Scholar] [CrossRef] [PubMed]
- Cameron, T.W.M. On two new species of nematode from the Scottish Red Deer. J. Helminthol. 1931, 9, 213–216. [Google Scholar] [CrossRef]
- Alberti, E.G.; Gioia, G.; Sironi, G.; Zanzani, S.; Riccaboni, P.; Magrini, M.; Manfredi, M.T. Elaphostrongylus cervi in a population of red deer (Cervus elaphus) and evidence of cerebrospinal nematodiasis in small ruminants in the province of Varese, Italy. J. Helminthol. 2011, 85, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Sugár, L.; Kával, A. Observations of Elaphostrongylus cervi (Cameron 1931) in native deer herd (In Hungarian). Parasit. Hung. 1977, 10, 95–96. [Google Scholar]
- Vicente, J.; Gortázar, C. High prevalence of large spiny-tailed protostrongylid larvae in Iberian red deer. Vet. Parasitol. 2001, 96, 165–170. [Google Scholar] [CrossRef]
- Kowal, J.; Kornaś, S.; Nosal, P.; Basiaga, M.; Wajdzik, M.; Skalska, M.; Wyrobisz, A. Lungworm (Nematoda: Protostrongylidae) infection in wild and domestic ruminants from Małopolska region of Poland. Ann. Parasitol. 2016, 62, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Handeland, K.; Gibbons, L.M.; Skorping, A. Experimental Elaphostrongylus cervi infection in sheep and goats. J. Comp. Pathol. 2000, 123, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Řezáč, P.; Palkovič, L.; Holasová, E.; Bušta, J. Modes of entry of the first-stage larvae of Elaphostrongylus cervi (Nematoda: Protostrongylidae) into pulmonated snails Arianta arbustorum and Helix pomatia. Folia Parasitol. 1994, 41, 209–214. [Google Scholar]
- Pusterla, N.; Caplazi, P.; Hertzberg, H.; Ehrensperger, F.; Braun, U. Elaphostrongylus cervi infection in a Swiss goat. Vet. Rec. 2001, 148, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.C.; McAllum, H.T.F. Letter: Dictyocaulus viviparus and Elaphostrongylus cervi in wapiti. N. Z. Vet. J. 1976, 24, 23. [Google Scholar] [CrossRef]
- Lankester, M.W.; Northcott, T.H. Elaphostrongylus cervi Cameron 1931 (Nematoda: Metastrongyloides) in caribou (Rangifer tarandus caribou) of Newfoundland. Can. J. Zool. 1979, 57, 1384–1392. [Google Scholar] [CrossRef]
- Bandelj, P.; Logar, K.; Usenik, A.M.; Vengust, M.; Ocepek, M. An improved qPCR protocol for rapid detection and quantification of Clostridium difficile in cattle feces. FEMS Microbiol. Lett. 2013, 341, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajadhar, A.; Steeves-Gurnsey, T.; Kendall, J.; Lankester, M.; Stéen, M. Differentiation of dorsal-spined elaphostrongyline larvae by polymerase chain reaction amplification of ITS-2 of rDNA. J. Wildl. Dis. 2000, 36, 713–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, A.; Woolf, A. The development of clinical signs and the population significance of neurologic disease in a captive wapiti herd. J. Wildl. Dis. 1978, 14, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Oevermann, A.; Zurbriggen, A.; Vandevelde, M. Rhombencephalitis Caused by Listeria monocytogenes in Humans and Ruminants: A Zoonosis on the Rise? Interdiscip. Perspect. Infect. Dis. 2010, 2010, 632513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handeland, K.; Flåøyen, A. Enzootic ataxia in a Norwegian red deer herd. Acta Vet. Scand. 2000, 41, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Blakley, B.R.; Haigh, J.C.; McCarthy, W.D. Concentrations of copper in tissues of wapiti raised in Saskatchewan. Can. Vet. J. 1992, 33, 549–550. [Google Scholar]
- Pryadko, E.I.; Visokov, S.N.; Frolov, V.S. Epizootiology of elaphostrongylosis in deer. Parasites Farm Anim. Kazakhstan 1963, 2, 74–85. [Google Scholar]
- Valcárcel, F.; Corchero, J.; Olmeda, A.S.; García Romero, C. Epidemiology of cerebrospinal Elaphostrongylus cervi infection in red deer in central Spain. J. Helminthol. 2004, 78, 265–270. [Google Scholar] [CrossRef]
- Wasel, S.M.; Samuel, W.M.; Crichton, V. Distribution and ecology of meningeal worm, Parelaphostrongylus tenuis (Nematoda), in northcentral North America. J. Wildl. Dis. 2003, 39, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Jacques, C.N.; Jenks, J.A.; Klaver, R.W.; Dubay, S.A. Associations among Habitat Characteristics and Meningeal Worm Prevalence in Eastern South Dakota, USA. J. Wildl. Dis. 2017, 53, 131–135. [Google Scholar] [CrossRef]
- Valcárcel, F.; García Romero, C. First report of Elaphostrongylus cervi in Spanish red deer Cervus elaphus hispanicus. J. Helminthol. 2002, 76, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Lankester, M.W.; Strelive, U.R. Further experimental studies of Pneumostrongylus tenuis in cervids. Can. J. Zool. 1966, 44, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Pybus, M.J.; Samuel, W.M.; Welch, D.A.; Smits, J.; Haigh, J.C. Mortality of fallow deer (Dama dama) experimentally-infected with meningeal worm, Parelaphostrongylus tenuis. J. Wildl. Dis. 1992, 28, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Handeland, K.; Gibbons, L.M.; Skorping, A. Aspects of the life cycle and pathogenesis of Elaphostrongylus cervi in red deer (Cervus elaphus). J. Parasitol. 2000, 86, 1061–1066. [Google Scholar] [CrossRef]
- Samuel, W.M.; Pybus, M.J.; Welch, D.A.; Wilke, C.J. Elk as a Potential Host for Meningeal Worm: Implications for Translocation. J. Wildl. Manag. 1992, 56, 629–639. [Google Scholar] [CrossRef]
- Mason, P.C.; Gladden, N.R. Survey of internal parasitism and anthelmintic use in farmed deer. N. Z. Vet. J. 1983, 31, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Valcárcel, F.; Meana, A.; Sacristán, E.; Uriarte, J.; Calvete, C.; Calavia, R.; Martínez Valladares, M.; Rojo-Vázquez, F.A. Small ruminants farm management practices in Spain and its influence on the development of anthelmintic resistance. Rev. Ibero-Latinoam. Parasitol. 2013, 72, 151–163. [Google Scholar]
- Isaza, R.; Courtney, C.H.; Kollias, G.V. Survey of parasite control programs used in captive wild ruminants. Zoo Biol. 1990, 9, 385–392. [Google Scholar] [CrossRef]
- Rodriguez, O.; Fernandez de Mera, J.G.; Vicente, J.; Peña, A.; Gortazar, C. Efficacy of in-feed-administered ivermectin on Elaphostrongylus cervi first-stage excretion in red deer (Cervus elaphus). Parasitol. Res. 2006, 98, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Mitskevich, V.Y. Life cycle of Elaphostrongylus rangiferi MIZ. In Parasites of Farm Animals in Kazakhstan; Izdatelstvo Akademii Nauk Kazakhstan: Alma-Ata, Kazakhstan, 1964; pp. 49–60. [Google Scholar]
- Panin, V.Y. Life cycle of Elaphostrongylus panticola Lubimov. In Parasites of Farm Animals in Kazakhstan; Izdatelstvo Akademii Nauk Kazakhstan: Alma-Ata, Kazakhstan, 1964; pp. 34–48. [Google Scholar]
- Sugár, L. On the nematode infestation of wild ruminants in Hungary (In Hungarian). Parasitol. Hung. 1978, 11, 146–148. [Google Scholar]
- Morandi, F.; Galuppi, R.; Nicoloso, S.; Benazzi, C.; Tampieri, M.P.; Simoni, P. Larvae of Elaphostrongylus cervi in a population of free-living red deer in Italy. J. Wildl. Dis. 2006, 42, 870–872. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandelj, P.; Juntes, P.; Vengušt, G.; Vengušt, D.Ž. The First Report of Elaphostrongylus cervi Infection in Two Imported Wapitis (Cervus canadensis) in Slovenia. Vet. Sci. 2022, 9, 19. https://doi.org/10.3390/vetsci9010019
Bandelj P, Juntes P, Vengušt G, Vengušt DŽ. The First Report of Elaphostrongylus cervi Infection in Two Imported Wapitis (Cervus canadensis) in Slovenia. Veterinary Sciences. 2022; 9(1):19. https://doi.org/10.3390/vetsci9010019
Chicago/Turabian StyleBandelj, Petra, Polona Juntes, Gorazd Vengušt, and Diana Žele Vengušt. 2022. "The First Report of Elaphostrongylus cervi Infection in Two Imported Wapitis (Cervus canadensis) in Slovenia" Veterinary Sciences 9, no. 1: 19. https://doi.org/10.3390/vetsci9010019
APA StyleBandelj, P., Juntes, P., Vengušt, G., & Vengušt, D. Ž. (2022). The First Report of Elaphostrongylus cervi Infection in Two Imported Wapitis (Cervus canadensis) in Slovenia. Veterinary Sciences, 9(1), 19. https://doi.org/10.3390/vetsci9010019