
In Vitro Neuroprotective Effect of the Bovine Umbilical Vein Endothelial Cell Conditioned Medium Mediated by Downregulation of IL-1β, Caspase-3, and Caspase-9 Expression

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cells
2.3. Isolation, Cultivation, and Cryopreservation of Bovine Umbilical Vein Endothelial Cells (BUVECs)
2.4. Maintenance of PC12 and SH-SY5Y Cell Lines
2.5. Isolation and Analysis of BUVEC-CM with Liquid Chromatography–Mass Spectrometry (LC-MS) and High-Resolution Mass Spectrometry (HR-MS)
2.6. Morphological Assessment of the PC12 and SH-SY5Y Cell Lines
2.7. CCK-8 Cell Viability Assay of PC12 and SH-SY5Y Cell Lines with BUVEC-CM
2.8. Treatment and Preparation of Cell Lysate and Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of IL-1β, Caspase-3, and Caspase-9
2.9. Statistics
2.10. Ethical Clearance
3. Results
3.1. LC-MS and HR-MS Analysis of BUVEC-CM
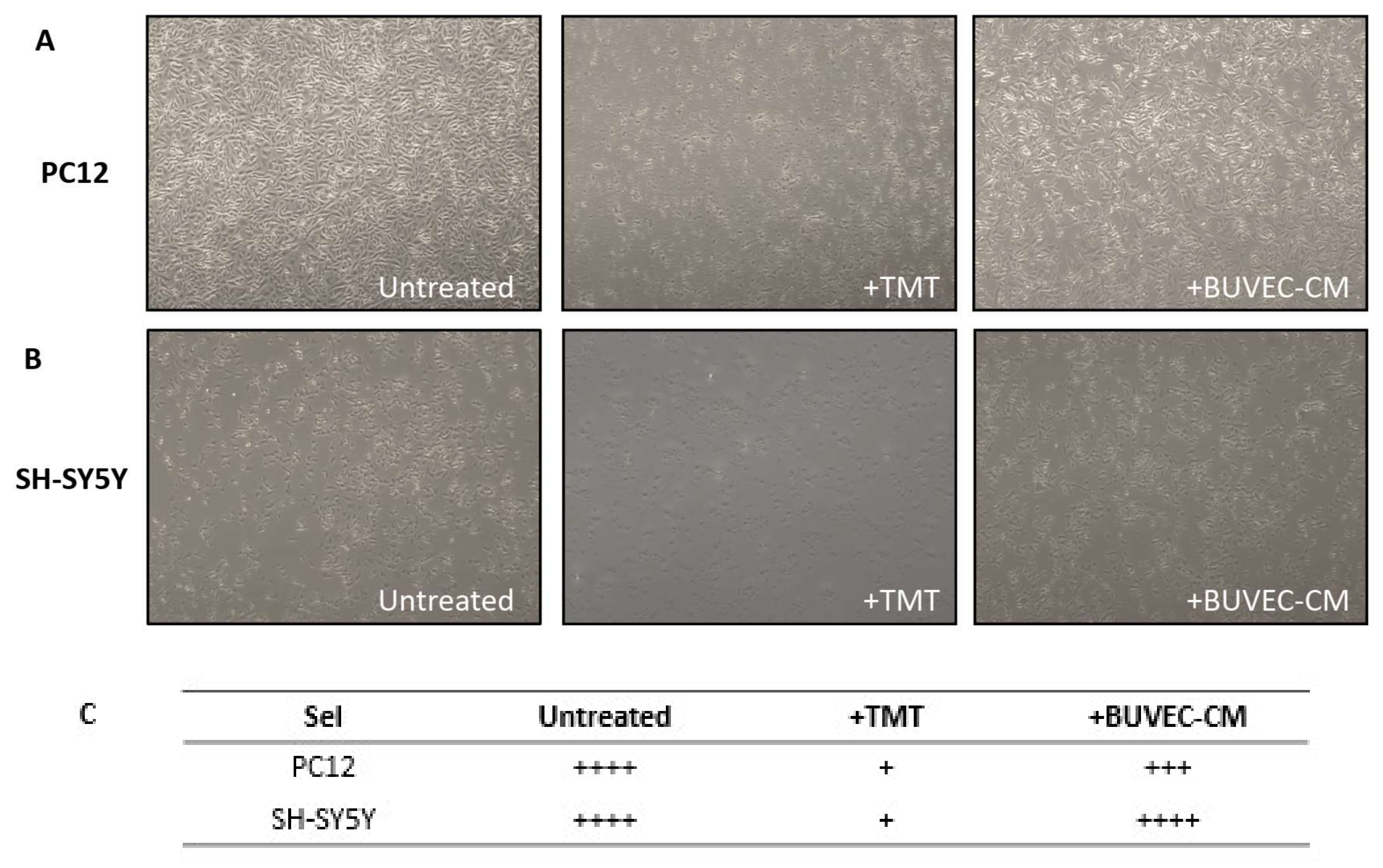
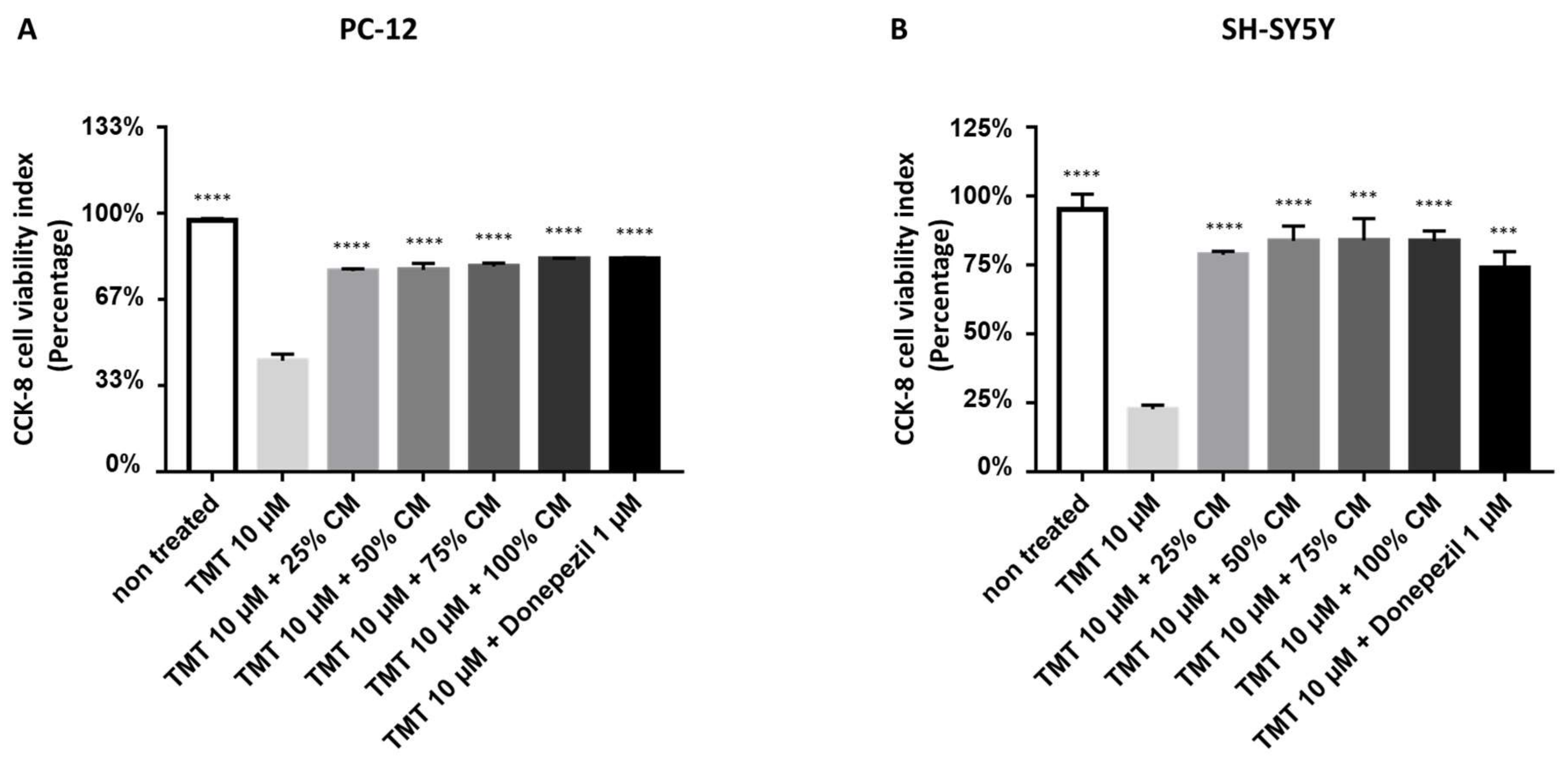
3.2. BUVEC-CM Maintains the Morphological Integrity and Cellular Viability of Rat Pheochromocytoma PC12 and Human Neuron-like SH-SY5Y Cell Lines after Treatment with the Neurotoxic Agent TMT
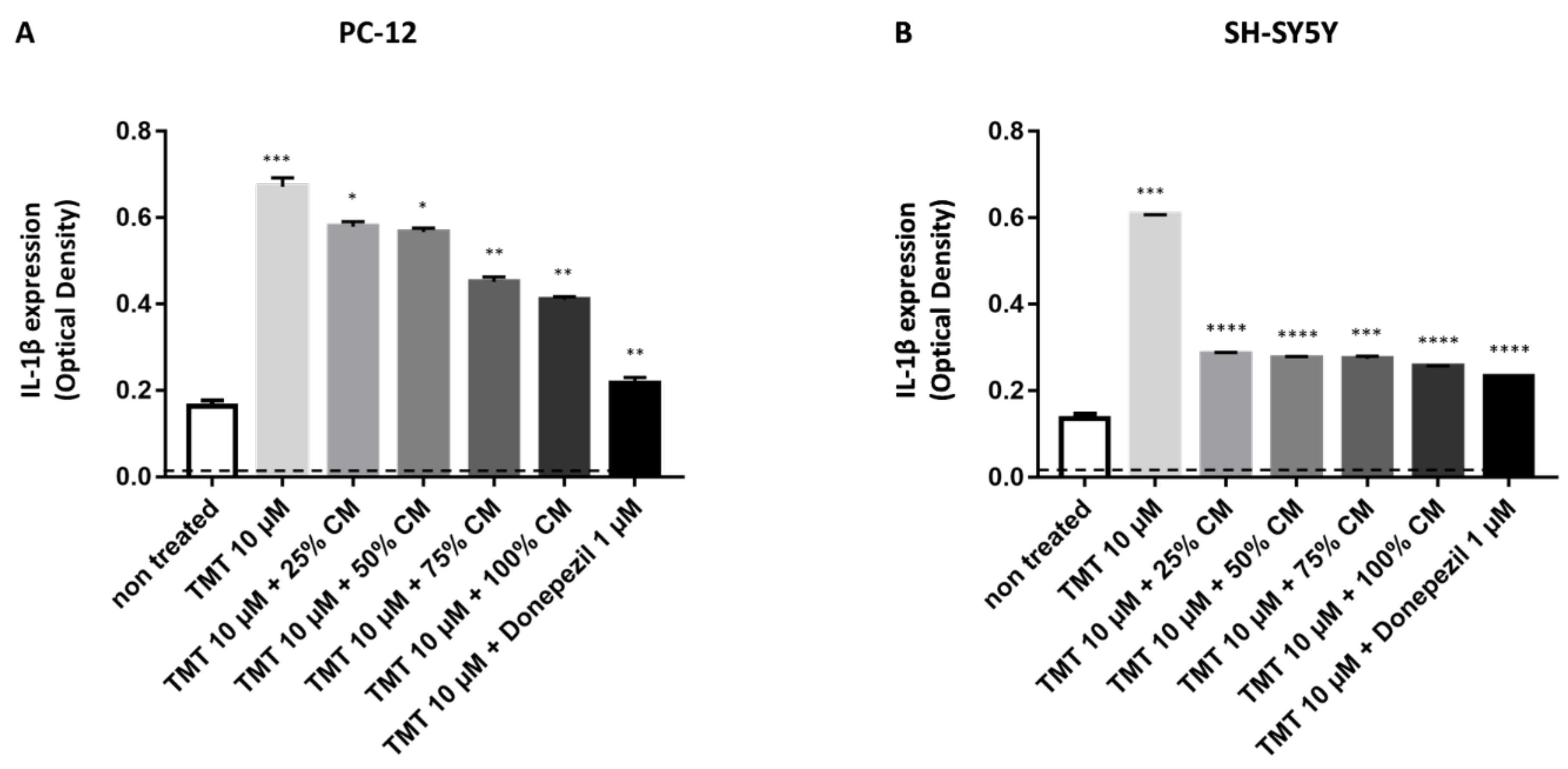
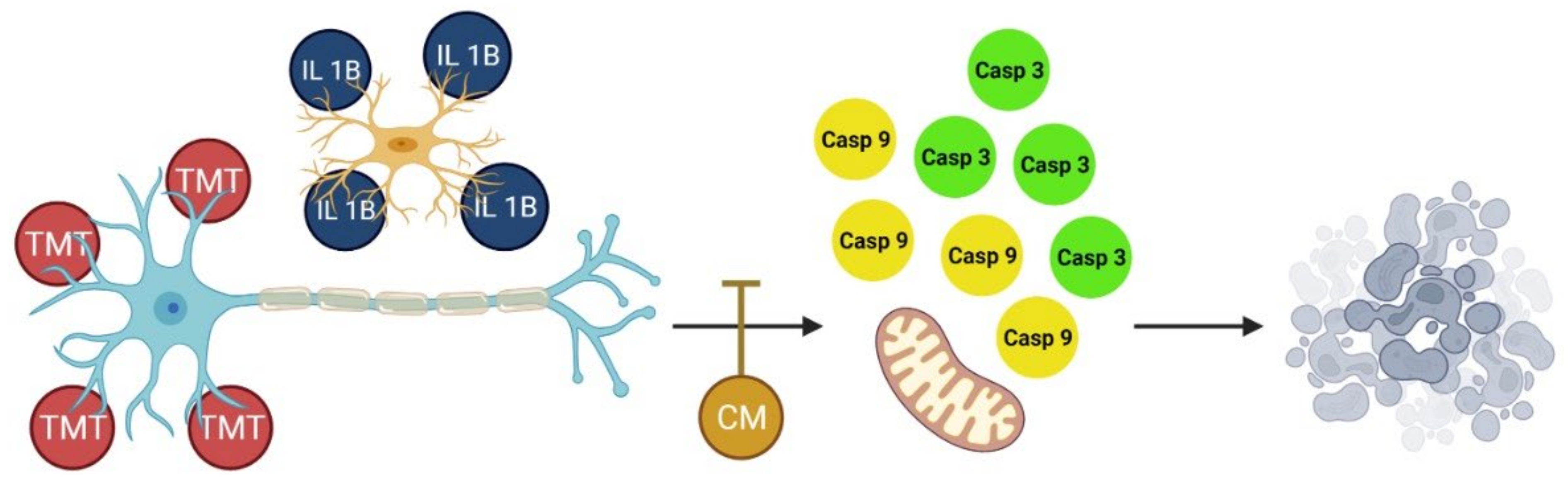
3.3. BUVEC-CM Inhibits Proinflammatory Cytokines IL-1β Production of Rat Pheochromocytoma PC12 and Human Neuron-like SH-SY5Y Cell Lines after Induction with the Proapoptotic Substance TMT
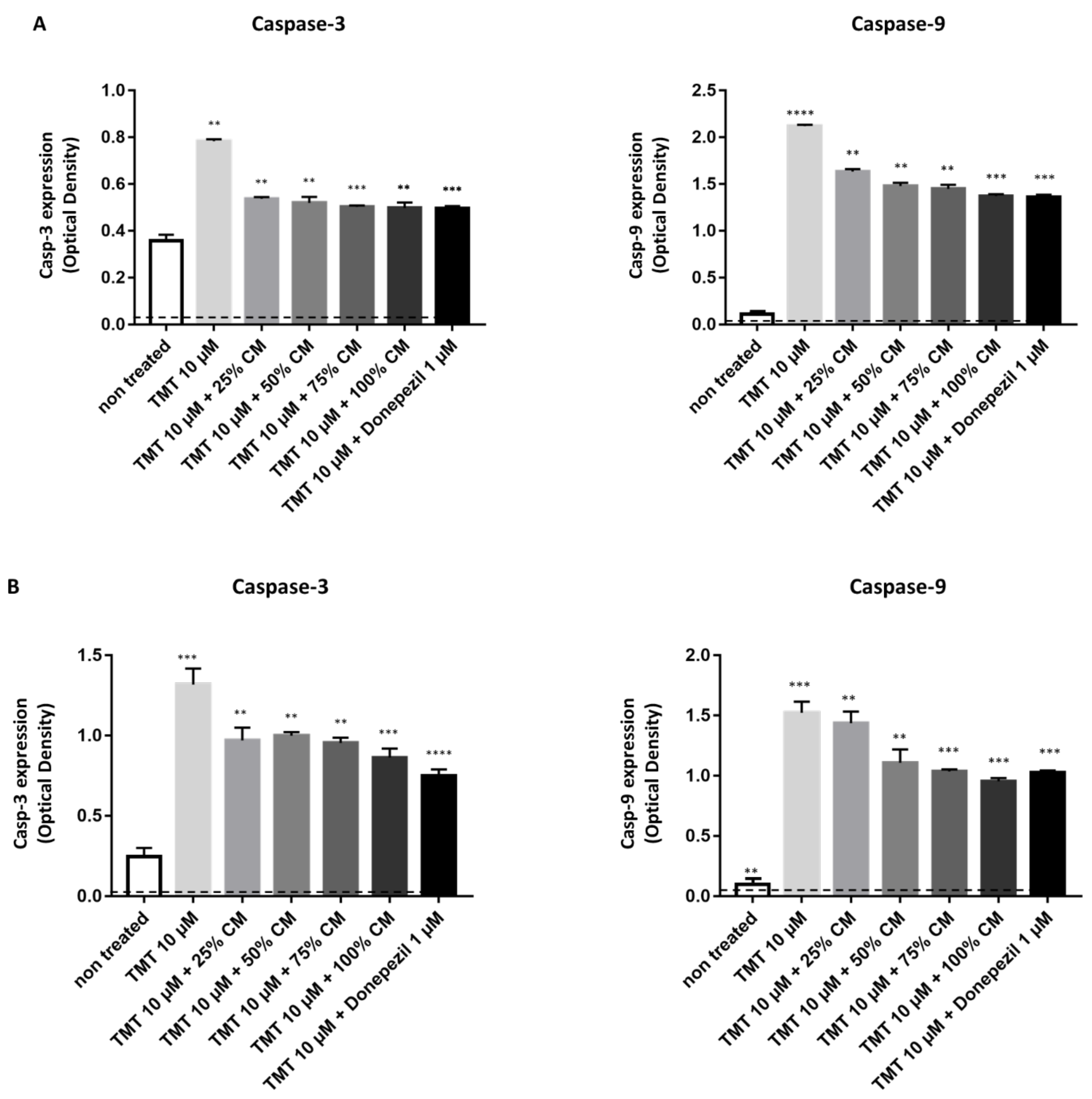
3.4. BUVEC-CM Inhibits Proapoptotic Caspase-3 and Caspase-9 Production of Rat Pheochromocytoma PC12 and Human Neuron-like SH-SY5Y Cell Lines Treated with TMT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed]
- Ruffini, N.; Klingenberg, S.; Schweiger, S.; Gerber, S. Common Factors in Neurodegeneration: A Meta-Study Revealing Shared Patterns on a Multi-Omics Scale. Cells 2020, 9, 2642. [Google Scholar] [CrossRef] [PubMed]
- Deuschl, G.; Beghi, E.; Fazekas, F.; Varga, T.; Christoforidi, K.A.; Sipido, E.; Feigin, V.L. The burden of neurological diseases in Europe: An analysis for the Global Burden of Disease Study 2017. Lancet Public Health 2020, 5, e551–e567. [Google Scholar] [CrossRef]
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Ellenbogen, R.G. Global, regional, and national burden of neurological disorders, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Kiyota, T.; Okuyama, S.; Swan, R.J.; Jacobsen, M.T.; Gendelman, H.E.; Ikezu, T. CNS expression of anti-inflammatory cytokine interleukin-4 attenuates Alzheimer’s disease-like pathogenesis in APP+PS1 bigenic mice. FASEB J. 2010, 24, 3093–3102. [Google Scholar] [CrossRef] [Green Version]
- Berebichez-Fridman, R.; Montero-Olvera, P.R.M. Sources and Clinical Applications of Mesenchymal Stem Cells. Sultan Qaboos Univ. Med. J. 2018, 18, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Reyhani, S.; Abbaspanah, B.; Mousavi, S.H. Umbilical Cord-derived Mesenchymal Stem Cells in Neurodegenerative Disorders: From Literature to Clinical Practice. Regen. Med. 2020, 15, 1561–1578. [Google Scholar] [CrossRef]
- Song, W.-J.; Yun, J.-H.; Jeong, M.-S.; Kim, K.-N.; Shin, T.; Kim, H.-C.; Wie, M.-B. Inhibitors of Lipoxygenase and Cyclooxygenase-2 Attenuate Trimethyltin-Induced Neurotoxicity through Regulating Oxidative Stress and Pro-Inflammatory Cytokines in Human Neuroblastoma SH-SY5Y Cells. Brain Sci. 2021, 11, 1116. [Google Scholar] [CrossRef]
- Shen, J.; Xu, S.; Zhou, H.; Liu, H.; Jiang, W.; Hao, J.; Hu, Z. IL-1β Induces Apoptosis and Autophagy via Mitochondria Pathway in Human Degenerative Pulposus Cells. Sci. Rep. 2017, 7, 41067. [Google Scholar] [CrossRef]
- Pawitan, J.A. Review Article: Prospect of Stem Cell Conditioned Medium in Regenerative Medicine. Biomed Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Chen, Y.C.; Chang, Y.C.; Wang, L.W.; Lin, Y.C.; Chiang, Y.L.; Ho, C.J.; Huang, C.C. Human Umbilical Vein Endothelial Cells Protect Against Hypoxic-Ischemic Damage in Neonatal Brain via Stromal Cell-derived Factor 1/C-X-C Chemokine Receptor Type 4. Stroke 2013, 44, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Debbarma, P.; Mondal, T.; Manna, C.; Kumar, K.; Mukerjee, J.; Das, B.C.; Bag, S.; Das, K. Post-calving Umbilical Cord Tissue Offcut: A Potential Source for the Isolation of Bovine Mesenchymal Stem Cells. Vet. World 2020, 13, 2772–2779. [Google Scholar] [CrossRef] [PubMed]
- Medina-Leyte, D.J.; Domínguez-Pérez, M.; Mercado, I.; Villarreal-Molina, M.T.; Jacobo-Albavera, L. Use of Human Umbilical Vein Endothelial Cells (HUVEC) as a Model to Study Cardiovascular Disease: A Review. Appl. Sci. 2020, 10, 938. [Google Scholar] [CrossRef] [Green Version]
- Mataram, M.B.A.; Hening, P.; Harjanti, N.F.; Karnati, S.; Wasityastuti, W.; Nugrahaningsih, D.A.A.; Kusindarta, D.L.; Wihadmadyatami, H. The neuroprotective effect of ethanolic extract Ocimum sanctum Linn. in the regulation of neuronal density in hippocampus areas as a central autobiography memory on the rat model of Alzheimer’s disease. J. Chem. Neuroanat. 2021, 111, 101885. [Google Scholar] [CrossRef]
- Kiaei, M. New hopes and challenges for treatment of neurodegenerative disorders: Great opportunities for young neuroscientists. Basic Clin. Neurosci. 2013, 4, 3–4. [Google Scholar]
- Honig, L.S.; Vellas, B.; Woodward, M.; Boada, M.; Bullock, R.; Borrie, M.; Hager, K.; Andreasen, N.; Scarpini, E.; Liu-Seifert, H.; et al. Trial of Solanezumab for Mild Dementia Due to Alzheimer’s Disease. N. Engl. J. Med. 2018, 378, 321–330. [Google Scholar] [CrossRef]
- Guadagno, J.; Xu, X.; Karajgikar, M.; Brown, A.; Cregan, S.P. Microglia-derived TNFα Induces Apoptosis in Neural Precursor Cells via Transcriptional Activation of the Bcl-2 Family Member Puma. Cell Death Dis. 2013, 4, e538. [Google Scholar] [CrossRef]
- Wang, W.Y.; Tan, M.S.; Yu, J.T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar]
- Amelio, M.D.; Cavallucci, V.; Cecconi, F. Neuronal Caspase-3 Signaling: Not only Cell Death. Cell Death Differ. 2010, 17, 1104–1114. [Google Scholar] [CrossRef]
- Yang, J.P.; Liu, H.J.; Yang, H.; Feng, P.Y. Therapeutic time window for the neuroprotective effects of NGF when administered after focal cerebral ischemia. Neurol. Sci. 2011, 32, 433–441. [Google Scholar] [CrossRef]
- Del Re, D.P.; Amgalan, D.; Linkermann, A.; Liu, Q.; Kitsis, R.N. Fundamental Mechanisms of Regulated Cell Death and Implications for Heart Disease. Physiol. Rev. 2019, 99, 1765–1817. [Google Scholar] [CrossRef]
- Tjalsma, H.; Bolhuis, A.; Jongbloed, J.D.; Bron, S.; van Dijl, J.M. Signal peptide-dependent protein transport in Bacillus subtilis: A genome-based survey of the secretome. Microbiol. Mol. Biol. Rev. 2000, 64, 515–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagiannis, G.S.; Pavlou, M.P.; Diamandis, E.P. Cancer secretomics reveal pathophysiological pathways in cancer molecular oncology. Mol. Oncol. 2010, 4, 496–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, C.A.; Fraga, J.S.; Grãos, M.; Neves, N.M.; Reis, R.L.; Gimble, J.M.; Sousa, N.; Salgado, A.J. The secretome of stem cells isolated from the adipose tissue and Wharton jelly acts differently on central nervous system derived cell populations. Stem Cell Res. Ther. 2012, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Salgado, A.J.; Sousa, J.C.; Costa, B.M.; Pires, A.O.; Mateus-Pinheiro, A.; Teixeira, F.G.; Pinto, L.; Sousa, N. Mesenchymal stem cells secretome as a modulator of the neurogenic niche: Basic insights and therapeutic opportunities. Front. Cell. Neurosci. 2015, 9, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, F.G.; Carvalho, M.M.; Sousa, N.; Salgado, A.J. Mesenchymal stem cells secretome: A new paradigm for central nervous system regeneration? Cell. Mol. Life Sci. 2013, 70, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Tang, J.C.; Pan, M.X.; Chen, S.F.; Zhao, D.; Zhang, Y.; Liao, H.B.; Zhuang, Y.; Lei, R.X.; Wang, S.; et al. l-lysine confers neuroprotection by suppressing inflammatory response via microRNA-575/PTEN signaling after mouse intracerebral hemorrhage injury. Exp. Neurol. 2020, 327, 113214. [Google Scholar] [CrossRef]
- Ullah, R.; Jo, M.H.; Riaz, M.; Alam, S.I.; Saeed, K.; Ali, W.; Rehman, I.U.; Ikram, M.; Kim, M.O. Glycine, the smallest amino acid, confers neuroprotection against d-galactose-induced neurodegeneration and memory impairment by regulating c-Jun N-terminal kinase in the mouse brain. J. Neuroinflamm. 2020, 17, 303. [Google Scholar] [CrossRef]
- Li, S.Y.; Qi, Y.; Hu, S.H.; Piao, F.Y.; Guan, H.; Wang, Z.M.; Chen, R.L.; Liu, S. Mesenchymal Stem Cells-Conditioned Medium Protects PC12 Cells Against 2,5-hexanedione-induced Apoptosis Via Inhibiting Mitochondria-dependent Caspase 3 Pathway. Toxicol. Ind. Health 2015, 33, 107–118. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chang, Y.W.; Tan, K.P.; Shen, Y.S.; Wang, Y.H.; Chang, C.H. Can Mesenchymal Stem Cells and Their Conditioned Medium Assist Inflammatory Chondrocytes Recovery? PLoS ONE 2018, 13, e0205563. [Google Scholar]
- Cai, L.; Qin, X.; Xu, Z.; Song, Y.; Jiang, H.; Wu, Y.; Ruan, H.; Chen, J. Comparison of Cytotoxicity Evaluation of Anticancer Drugs between Real-Time Cell Analysis and CCK-8 Method. ACS Omega 2019, 4, 12036–12042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Icöz, S.K.; Zhou, P.; Guo, Y.; Loganathan, S.; Brlecic, P.; Radovits, T.; Sayour, A.A.; Ruppert, M.; Veres, G.; Karck, M.; et al. Mesenchymal Stem Cell-derived Conditioned Medium Protects Vascular Grafts of Brain-dead Rats Against in vitro Ischemia/reperfusion Injury. Stem Cell Res. Ther. 2021, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Kocherova, I.; Bryja, A.; Mozdziak, P.; Volponi, A.A.; Konwińska, M.D.; Kempisty, H.P.; Antosik, P.; Bukowska, D.; Bruska, M.; Iżycki, D.; et al. Human Umbilical Vein Endothelial Cells (HUVECs) Co-Culture with Osteogenic Cells: From Molecular Communication to Engineering Prevascularised Bone Grafts. J. Clin. Med. 2019, 8, 1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, Y.; Liang, Y.; Du, Q.; Fan, P.; Xu, H.; Xu, Y.; dan Shi, N. Apoptosis Induced by Trimethyltin Chloride in Human Neuroblastoma Cells SY5Y is Regulated by a Balance and Cross-talk between NF-kB and MAPKs Signaling Pathways. Arch. Toxicol. 2013, 87, 1273–1285. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Zhang, J.; Wang, C.; Yu, A.; Chen, Y.; dan Zuo, Z. Acute Trimethyltin Exposure Induces Oxidative Stress Response and Neuronal Apoptosis in Sebastiscus marmoratus. Aquat. Toxicol. 2008, 90, 58–64. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Metabolite |
|---|---|
| 1 | Isoleucine |
| 2 | Leucine |
| 3 | Valine |
| 4 | Ethanol |
| 5 | Threonine |
| 6 | Lactate |
| 7 | Alanine |
| 8 | Lysine |
| 9 | Arginine |
| 10 | Acetate |
| 11 | Glutamate |
| 12 | Glutamine |
| 13 | Methionine |
| 14 | Hydroxyproline |
| 15 | Hydroxyurasile |
| 16 | Choline |
| 17 | Pruvate |
| 18 | a-Glucose |
| 19 | b-Glucose |
| 20 | Tyrosine |
| 21 | Phenylalanine |
| 22 | Histidine |
| 23 | Tryptophan |
| 24 | Palmitic acid |
| 25 | Ethyl palmitoleate |
| 26 | Stearic acid |
| 27 | Cholecalciferol |
| 28 | Oleamide |
| 29 | Ethyl myristate |
| 30 | Nicotinamide |
| 31 | Format |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larasati, V.A.; Lembang, G.V.; Tjahjono, Y.; Winarsih, S.; Ana, I.D.; Wihadmadyatami, H.; Kusindarta, D.L. In Vitro Neuroprotective Effect of the Bovine Umbilical Vein Endothelial Cell Conditioned Medium Mediated by Downregulation of IL-1β, Caspase-3, and Caspase-9 Expression. Vet. Sci. 2022, 9, 48. https://doi.org/10.3390/vetsci9020048
Larasati VA, Lembang GV, Tjahjono Y, Winarsih S, Ana ID, Wihadmadyatami H, Kusindarta DL. In Vitro Neuroprotective Effect of the Bovine Umbilical Vein Endothelial Cell Conditioned Medium Mediated by Downregulation of IL-1β, Caspase-3, and Caspase-9 Expression. Veterinary Sciences. 2022; 9(2):48. https://doi.org/10.3390/vetsci9020048
Chicago/Turabian StyleLarasati, Vinny A., Gregorius V. Lembang, Yudy Tjahjono, Sugi Winarsih, Ika Dewi Ana, Hevi Wihadmadyatami, and Dwi L. Kusindarta. 2022. "In Vitro Neuroprotective Effect of the Bovine Umbilical Vein Endothelial Cell Conditioned Medium Mediated by Downregulation of IL-1β, Caspase-3, and Caspase-9 Expression" Veterinary Sciences 9, no. 2: 48. https://doi.org/10.3390/vetsci9020048
APA StyleLarasati, V. A., Lembang, G. V., Tjahjono, Y., Winarsih, S., Ana, I. D., Wihadmadyatami, H., & Kusindarta, D. L. (2022). In Vitro Neuroprotective Effect of the Bovine Umbilical Vein Endothelial Cell Conditioned Medium Mediated by Downregulation of IL-1β, Caspase-3, and Caspase-9 Expression. Veterinary Sciences, 9(2), 48. https://doi.org/10.3390/vetsci9020048